鯉sPLA2-III基因結(jié)構(gòu)、系統(tǒng)發(fā)育和表達(dá)特征
2019-11-26 02:17:08許迪輝李紅霞李建林唐永凱磊沈澤恩俞菊華
水生生物學(xué)報 2019年6期
關(guān)鍵詞:一致性
許迪輝 李紅霞, 李建林, 唐永凱, 王 欽 張 磊沈澤恩 俞菊華,
(1. 南京農(nóng)業(yè)大學(xué)無錫漁業(yè)學(xué)院, 無錫 214128; 2. 中國水產(chǎn)科學(xué)研究院淡水漁業(yè)研究中心農(nóng)業(yè)部淡水魚類遺傳育種和養(yǎng)殖生物學(xué)重點開放實驗室, 無錫 214128)
磷脂酶A2(Phospholipase A2, PLA2)是一種水解磷脂二位酰基(sn-2)生成非酯化脂肪酸和溶血磷脂的酶[1], 廣泛存在于動植物體內(nèi)。PLA2在生物體內(nèi)發(fā)揮多種生理功能, 如膜磷脂的降解和代謝、信號傳導(dǎo)、宿主防御等[2]。PLA2可分為分泌型PLA2(sPLA2)、胞漿型PLA2(cPLA2)、非鈣依賴型PLA2(iPLA2)、血小板活化因子乙酰水解酶(PAFAH)和溶酶體型PLA2(LPLA2)等[3]。其中, 分泌型PLA2(sPLA2)是PLA2中最大的亞家族, 是導(dǎo)致炎性脂質(zhì)介質(zhì)產(chǎn)生的關(guān)鍵分子[4]。sPLA2在中心結(jié)構(gòu)域上均含有偶數(shù)個半胱氨酸, 可形成二硫鍵, 對穩(wěn)定和保持蛋白空間三維結(jié)構(gòu)起著重要作用[5]。
在sPLA2家族中, sPLA2-III(sPLA2group III)蛋白分子量為55 kD, 遠(yuǎn)大于其他亞型, 由10個半胱氨酸構(gòu)成5個二硫鍵穩(wěn)定結(jié)構(gòu)[1,6]。蜂毒sPLA2-III只有中心結(jié)構(gòu)域(Centre sPLA2domain): 3個α螺旋、2個β折疊、1個Ca2+結(jié)合環(huán)。人sPLA2-III則除了中心結(jié)構(gòu)域外還有獨特的N端(N-terminus)和C端(C-terminus), 與其他哺乳動物相比, 人sPLA2-III的中心結(jié)構(gòu)域與蜂毒的同源性更高[6]。
哺乳動物sPLA2-III由多種細(xì)胞分泌, 尤其是附睪上皮細(xì)胞[7]、神經(jīng)元[8]、微血管的上皮組織細(xì)胞以及腫瘤細(xì)胞[9]有高表達(dá)。該基因具有多種生理功能。Sato等[7]發(fā)現(xiàn), sPLA2-III在小鼠睪丸組織中呈現(xiàn)高表達(dá), 該基因與小鼠精子獲能、精子運動能力、頂體反應(yīng)等有關(guān), 該基因缺失導(dǎo)致精子發(fā)育不良和弱精癥[10]。轉(zhuǎn)染sPLA2-III小鼠的低密度脂蛋白(Low-Density Lipoprotein, LDL)降低, 而小致密低密度脂蛋白(Small dense Low-Density Lipoprotein, sd LDL)上升, 后者能夠誘導(dǎo)巨噬細(xì)胞泡沫化,導(dǎo)致動脈粥樣硬化(Atherosis, AS), 在人類和apoE缺陷小鼠的動脈粥樣硬化病變組織中也檢測到了sPLA2-III, 顯示sPLA2-III有促AS作用[11,12]。sPLA2-III在腫瘤細(xì)胞中也有異常表達(dá), 轉(zhuǎn)染sPLA2-III的結(jié)腸癌裸鼠與正常結(jié)腸癌裸鼠對照發(fā)現(xiàn), 前者形成更大的實體瘤, 用小干擾RNA(Small interfering RNA)干擾sPLA2-III的表達(dá)后可降低腫瘤細(xì)胞增殖, 表明該基因在腫瘤病變、細(xì)胞增殖中發(fā)揮著重要作用[13]。
sPLA2-III最先在蜂毒和一些蜥蜴中發(fā)現(xiàn), 隨后,該基因在水母、海洋螺類以及蝎子中陸續(xù)發(fā)現(xiàn)[6],推測sPLA2-III是在脊椎動物和無脊椎動物分化之前就存在的原始基因[14]。連接脊椎動物和無脊椎動物的魚類, 在進化史中具有重要地位, 但至今對魚類sPLA2-III的研究報道不多, 僅在基因庫中能獲得一些主要研究魚類的sPLA2-III序列。鯉(Cyprinus carpioLinnaeus)是重要的經(jīng)濟淡水魚類之一,也是典型的異源四倍體, 是研究重復(fù)基因進化的好材料。本實驗挖掘了鯉sPLA2-III成員, 分析了序列的保守性、進行了2R (Two rounds of genome duplication)和3R (Fish-specific genome duplication or 3R)魚類[15]sPLA2-III基因成員的系統(tǒng)發(fā)育和線性分析, 揭示了它們的時空表達(dá)模式, 并為今后進一步揭示其生物學(xué)功能奠定基礎(chǔ)。
1 材料與方法
1.1 實驗魚
實驗鯉及鯉早期胚胎樣品均來自中國水產(chǎn)科學(xué)研究院淡水漁業(yè)研究中心宜興養(yǎng)殖基地, 實驗鯉為當(dāng)年人工繁殖的同池養(yǎng)殖魚種, 平均體重70 g, 體長10—15 cm。
1.2 實驗試劑與儀器
總RNA提取試劑盒購自O(shè)mega Bio-Tek公司。逆轉(zhuǎn)錄試劑盒[Prime ScriptTMRT Master Mix (Perfect Real Time)]、SYBR?Premix ExTaqIITM等購自TaKaRa生物(大連)有限公司。PCR儀、紫外分光光度計等儀器購自德國Eppendorf公司。熒光定量PCR儀為TaKaRa TP800型的實時熒光定量PCR儀。
1.3 實驗引物
通過Blast和線性分析在鯉全基因組中獲得五個sPLA2-III基因序列。P1-P2、P3-P4、P5-P6、P7-P8、P9-P10分別用于Ccpla2g3a1、Ccpla2g3a2、Ccpla2g3b、Ccpla2g3c1和Ccpla2g3c2的序列驗證,并上傳基因庫, 獲得序列號。根據(jù)獲得序列設(shè)計特異性引物P11-P12、P13-P14、P15-P16、P17-P18測定樣品Ccpla2g3a1/2、Ccpla2g3b、Ccpla2g3c1和Ccpla2g3c2的表達(dá), 由于Ccpla2g3a1和3a2 cDNA序列一致性較高, 這里共用一對引物。引物P19-P20為內(nèi)參β-actin的定量引物。引物由蘇州金唯智生物科技有限公司合成(表 1)。
1.4 鯉早期胚胎樣品
鯉受精卵在水溫22℃的孵化箱中孵育, 收集受精0.5h (胚盤出現(xiàn))、12h (原腸中期)、25h (肌節(jié)出現(xiàn))、35h (腦泡出現(xiàn))、60h (心臟、眼點出現(xiàn))、120h (即將破膜)的受精卵[16,17], 并保存于RNA sample protector中。
1.5 鯉組織樣品
鯉雌、雄各三條(平均體重70 g), 麻醉后, 采集心、肝、脾、體腎、頭腎、腸、腦、鰓和性腺9個組織, 于-80℃冰箱保存。
1.6 生物信息學(xué)分析
鯉sPLA2-III基因所在連鎖群信息來自NCBI,雀鱔(Lepisosteusoculatus)、斑馬魚(Danio rerio)、三刺魚(Gasterosteus aculeatus)等魚類的sPLA2-III線性分析使用Genomicus (v87.01)[18], 序列在NCBI (http://www.ncbi.nlm.nih.gov)獲得, 得到的cDNA序列采用ORF (Open reading frame)finder尋找正確的開放閱讀框(http://www.ncbi.nlm.nih.gov/orffinder/)。使用MatGAT軟件[19]分析鯉及其他魚類sPLA2-III氨基酸序列的相似性和一致性[20]。在MEGA 7.0軟件[21]中進行鯉及其他魚類sPLA2-III多序列比對, 比對結(jié)果用鄰接法, 以1000次自檢率構(gòu)建N-J系統(tǒng)進化樹。
使用FancyGene[22]繪制基因結(jié)構(gòu)示意圖。使用protparam在線工具對鯉sPLA2-III基因的相關(guān)理化性質(zhì)進行預(yù)測分析(http://web.expasy.org/cgi-bin/protparam/protparam), SignalP 4.1 (version 4.1)在線工具[23](http://www.cbs.dtu.dk/services/SignalP/)預(yù)測信號肽。

表1 序列驗證和qPCR所用引物Tab. 1 Primers used for sequences and qPCR
1.7 RNA提取、逆轉(zhuǎn)錄及實時熒光定量PCR
使用總RNA提取試劑盒提取RNA, 測定OD值,OD值在1.8—2.0, 并于1%瓊脂糖凝膠電泳, 檢測RNA完整性。
逆轉(zhuǎn)錄: 采用Prime ScriptTMRT Master Mix(Perfect Real Time)試劑盒說明書進行操作, 所得cDNA于-20℃保存。
獲得的cDNA用實時熒光定量PCR(qPCR)法分別檢測Ccpla2g3as、Ccpla2g3b、Ccpla2g3c1和Ccpla2g3c1在鯉受精0.5h、12h、25h、35h、60h、120h以及不同組織心、肝、脾、體腎、頭腎、腸、腦、鰓、性腺組織中的相對β-actin表達(dá)水平。過程參照TaKaRa公司生產(chǎn)的SYBR?Premix ExTaqIITM試劑盒說明書。
實驗結(jié)果采用2-ΔΔCt法[25]進行計算, 鯉早期胚胎不同發(fā)育階段和各組織的相對表達(dá)量分別使用SPSS 20.0軟件中的Turkey多重比較法和T檢驗進行差異顯著性分析,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結(jié)果
2.1 鯉sPLA2-III基因序列
鯉sPLA2-III基因序列特征通過同源搜索,在NCBI中挖掘到5個鯉sPLA2-III類似基因, 分別位于LHQP01025954、LHQP010113959、LHQP01 028027、LHQP01026208、LHQP01015502連鎖群,命名為Ccpla2g3a1、Ccpla2g3a2、Ccpla2g3b、Ccpla2g3c1和Ccpla2g3c2[25]。氨基酸命名同樣參照文獻(xiàn)[24]。除了LHQP01028027上的Ccpla2g3b外,其余4個PCR驗證結(jié)果除了個別堿基存在差異外,序列基本一致, LHQP01028027上的Ccpla2g3b由于堿基缺失, 導(dǎo)致提前終止, 閱讀框比克隆到的基因短很多。克隆的基因已上傳基因庫, 登錄號分別為: KF793833、KF793834、MH990666、MH99 0664和MH990665。
序列分析表明:Ccpla2g3a1和Ccpla2g3a2由7個外顯子和6個內(nèi)含子組成,Ccpla2g3a1外顯子1和外顯子4分別比Ccpla2g3a2多9和6 bp。開放閱讀框(ORF)分別為1593和1578 bp, 分別編碼530和525個氨基酸。均含有22個氨基酸組成的信號肽,成熟肽分子量分別為57.69和57.72 kD, 等電點(pI)分別為8.00和7.82。成熟肽由N端、sPLA2中心結(jié)構(gòu)域(Ccpla2g3a1為141個氨基酸,Ccpla2g3a2為140個氨基酸)和C端組成。
Ccpla2g3b由4個外顯子和3個內(nèi)含子組成, 外顯子長度分別為520、133、135和598 bp。開放閱讀框1386 bp, 編碼461個氨基酸。有27個氨基酸組成的信號肽, 成熟肽分子量為49.78 kD, 等電點為9.97。成熟肽由N端、141個氨基酸組成的中心結(jié)構(gòu)域和C端組成。
Ccpla2g3c1和Ccpla2g3c2均有4個外顯子和3個內(nèi)含子, 除外顯子4相差3 bp外, 其余3個外顯子長度均相同。開放閱讀框分別為2259和2262 bp, 分別編碼752和753個氨基酸。均含有19個氨基酸組成的信號肽, 成熟肽分子量分別為82.18和82.33 kD, 等電點分別為9.17和9.20。成熟肽由N端、141個氨基酸組成的中心結(jié)構(gòu)域和C端組成。圖 1為各基因的結(jié)構(gòu)簡圖, 表 2為蛋白結(jié)構(gòu)比較。
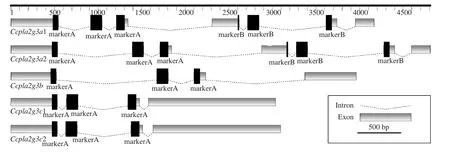
圖1 鯉sPLA2-III基因結(jié)構(gòu)示意圖Fig. 1 Schematic diagram of sPLA2 group III gene structure in common carp

表2 鯉sPLA2-III結(jié)構(gòu)域比較Tab. 2 sPLA2 group III domains in common carp
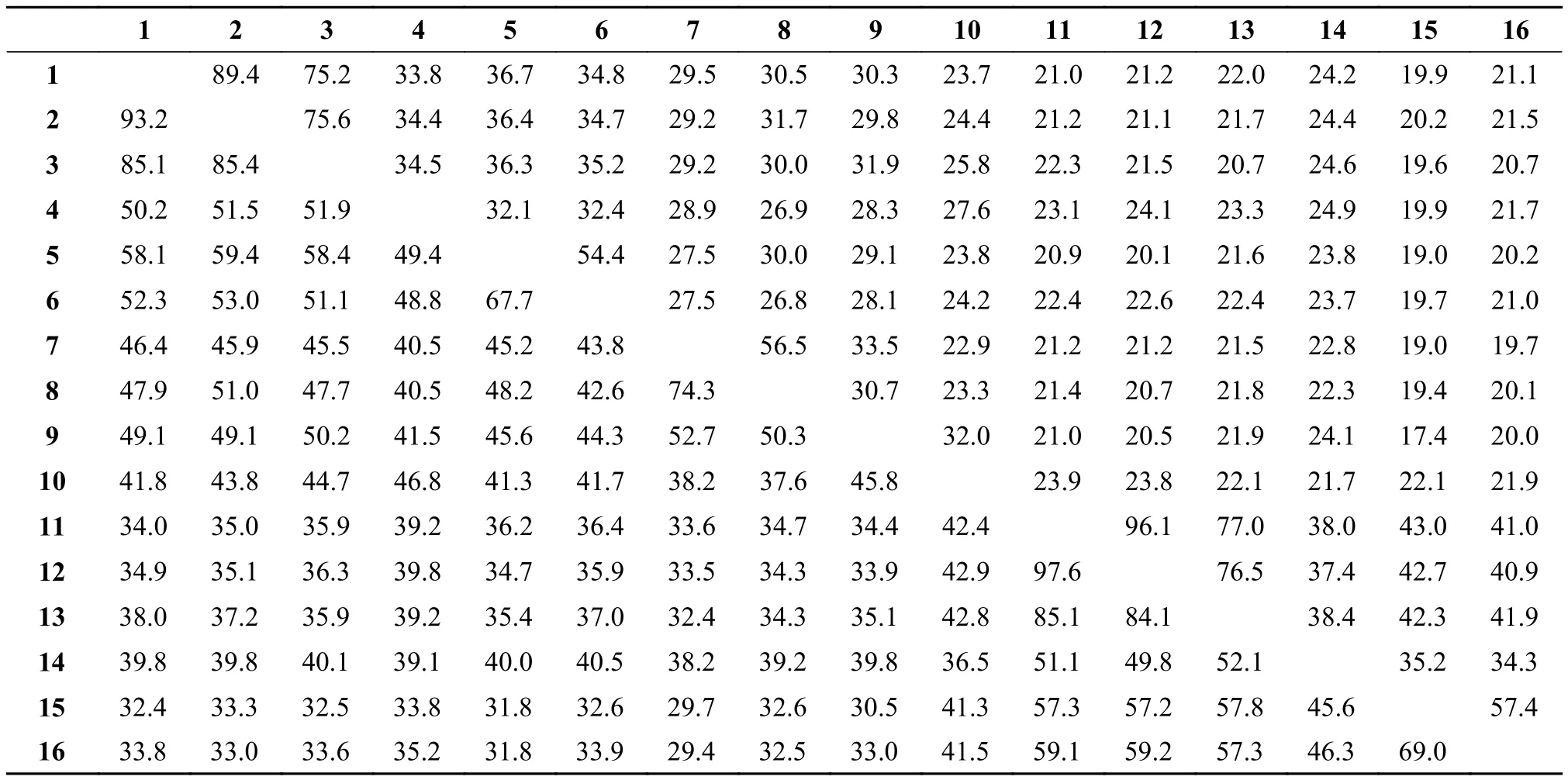
表3 鯉sPLA2-III與其他魚類sPLA2-III的氨基酸的一致性和相似性百分比(右上角, 一致性; 左下角, 相似性)Tab. 3 The similarity of amino acid identity of CcsPLA2-III and other fish sPLA2-III (upper triangle, identity; lower triangle, similarity)
sPLA2-III氨基酸序列比對通過Mat-GAT軟件對鯉與其他魚類sPLA2-III氨基酸序列進行相似性和一致性分析。結(jié)果顯示(表 3),CcPla 2g3cs之間的相似性(97.6%)和一致性(96.1%)最高,CcPla2g3as之間的相似性(93.2%)和一致性(89.4.%)次之, 但鯉sPLA2-III家族旁基因之間的相似性都較低(33.5%—46.4%)。與其他魚類相比, 同為鯉科魚的鯉和斑馬魚sPLA2-III的垂直同源基因之間相似性和一致性均最高, Pla2g3a之間相似性分別為85.1%和85.4%, 一致性分別為75.2%和75.6%;Pla2g3b之間相似性為74.3%, 一致性為56.5%;Pla2g3c之間相似性分別為85.1%和84.1%, 一致性分別為77.0%和76.5%。鯉與其他目的垂直同源基因之間的相似性和一致性普遍較低。
魚類sPLA2-III系統(tǒng)發(fā)育分析將鯉及其他魚類的同源序列采用Mega 7.0軟件構(gòu)建N-J系統(tǒng)進化樹(圖 2)。結(jié)果表明整個進化樹分成三個分支,所有魚類的Pla2g3as以95%置信度聚為一小支, 除鯉科魚之外的Pla2g3b以99%置信度聚為一小支, 這兩小支以96%置信度聚為完整的一支。CcPla2g3b與斑馬魚Pla2g3b單獨形成一大支。所有魚類的Pla2g3cs又以100%置信度聚為完整的一支。可以看出, Pla2g3as與Pla2g3bs先聚為一支, 最后與Pla2g3cs聚為一支, 共同形成魚類sPLA2-III的系統(tǒng)進化樹。
魚類sPLA2-III線性分析通過Genomicus(v87.01)進行雀鱔、斑馬魚、三刺魚、青鳉(Oryzias latipes)、羅非魚(Oreochromis niloticus)、亞馬遜帆魚(Poecilia formosa)sPLA2-III的線性分析。
分析發(fā)現(xiàn)(圖 3), 雀鱔(2R魚)中只有一個pla2g3a(b)基因, 其上游基因為ptpn11a(Protein tyrosine phosphatase, non-receptor type11, a)和RPH3A(Rabphilin 3A), 下游基因為tbx5a(T-box 5a)。雀鱔經(jīng)TGD (Teleost genome duplication, 又稱為3R)染色體加倍后, 硬骨魚類(3R魚)中出現(xiàn)了位于兩相似連鎖群上的pla2g3a與pla2g3b兩個不同的基因。ptpn11a和RPH3A都保守的存在于3R魚pla2g3a的上游, 而雀鱔下游基因tbx5a沒有發(fā)現(xiàn)。RPH3A的同源基因rap3ab[Rabphilin 3A homolog (mouse) b]仍保守地存在于3R魚pla2g3b的上游。pla2g3a在鯉中存在著Ccpla2g3a1與Ccpla2g3a2的兩個旁基因,RPH3A保守地存在于目的基因上游。Ccpla2g3b目前只在鯉的染色體LG1上發(fā)現(xiàn)了一個, 3R魚的HECTD4 (HECT domain E3 ubiquitin protein ligase 4)保守地存在于Ccpla2g3b下游。

圖2 魚類sPLA2-III成員系統(tǒng)進化樹Fig. 2 Phylogenetic tree of fish sPLA2 group III
雀鱔pla2g3c的上游基因為rab34a(RAB34,member RAS oncogene family b)和aptl(Adenine phosphoribosyltransferase-like), 下游基因為supt6h(SPT6 homolog, histone chaperone)和sdf2 (Stromal cell-derived factor2)。經(jīng)TGD后, 在3R魚中存在2個相似的連鎖群, 其中一個連鎖群上有pla2g3c, 除了三刺魚中aptl丟失,rab34a和2個基因仍保守的存在于3R魚pla2g3c的上游, 但雀鱔下游的supt6h和sdf2沒出現(xiàn)。在TGD過程中3R魚另一個pla2g3c基因已經(jīng)丟失, 但是雀鱔上游基因的同源基因rab34b以及下游基因supt6h和sdf2在3R魚另一連鎖群上中比較保守。鯉經(jīng)特異基因組復(fù)制,也發(fā)現(xiàn)了2個不同的Ccpla2g3cs基因, 其中,Ccpla2g3c1兩側(cè)基因與斑馬魚一致,Ccpla2g3c2的上游只發(fā)現(xiàn)了rab34a基因。
2.2 鯉sPLA2-III基因表達(dá)
鯉sPLA2-III基因在胚胎發(fā)育階段表達(dá)熒光定量PCR分別檢測了鯉sPLA2-III各基因分別在受精后0.5h、12h、25h、35h、60h、120h的表達(dá)變化。結(jié)果顯示(圖 4),Ccpla2g3as在鯉整個早期胚胎階段的表達(dá)量均較低。Ccpla2g3as在受精0.5h的表達(dá)量顯著高于25h和60h (P<0.05)。Ccpla2g3b在0.5h表達(dá)量最高, 顯著高于12h和120h (P<0.05), 極顯著高于25h、35h、60h (P<0.01), 于25h降到最低(P<0.01)。Ccpla2g3c1在0.5—60h期間表達(dá)量均較低, 120h表達(dá)量極顯著升高(P<0.01)。Ccpla2g3c2在0.5h至60h期間(除12h)表達(dá)量較低, 于120h表達(dá)量極顯著升高(P<0.01), 120h的表達(dá)量顯著高于12h(P<0.05)。
鯉sPLA2-III基因在不同組織表達(dá)熒光定量PCR結(jié)果顯示,Ccpla2g3as在肝臟中的表達(dá)量極顯著高于其他組織(P<0.01)。Ccpla2g3b在卵巢中的表達(dá)量極顯著高于其他組織(P<0.01); 精巢中的表達(dá)量顯著高于肝臟和肌肉(P<0.05), 極顯著高于其余組織(P<0.01); 卵巢中Ccpla2g3b極顯著高于精巢(P<0.01)。Ccpla2g3c1在腦中的表達(dá)量最高(P<0.01), 其他組織中表達(dá)量較低。Ccpla2g3c2在腦中表達(dá)量極顯著高于其他組織(P<0.01), 腸的表達(dá)量極顯著高于其余組織(P<0.01, 圖 5)。
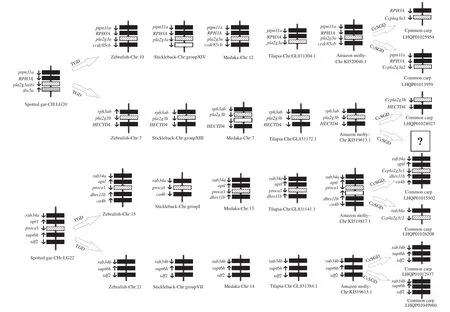
圖3 硬骨魚類及其祖先sPLA2-III共線性分析示意圖Fig. 3 Schematic diagram of synteny of sPLA2 group III in teleost’s and their ancestors
3 討論
3.1 鯉sPLA2-III基因
本文使用同源搜索在鯉基因組挖掘到了5個具有PLA2_bee_venom_like region的Ccpla2g3基因, 線性分析發(fā)現(xiàn)其中Ccpla2g3c是人PROCA1基因的同源基因, 追溯在人中定名為PROCA1的原因是其與cyclin A1能相互作用[26]。最初登錄為部分序列(登錄號: AAT09159), 但該序列在cDNA文庫中并沒有被找到, 目前該序列已被更正為Q8NCQ7。該序列也存在PLA2_bee_venom_like region結(jié)構(gòu)域, 但與人PLA2G3中的PLA2_bee_venom_like region差異較大, 與文章中的序列比較發(fā)現(xiàn)存在較大的差異, 沒有文獻(xiàn)[25]中確定的與cyclin A1作用的特征序列,人PROCA1基因在睪丸呈高表達(dá)。鑒于魚類PROCA1基因是否能與cyclin A1相互作用還沒有確認(rèn), 且其表達(dá)也與人的存在差異, 加之含有保守的PLA2_bee_venom_like region, 因此, 這里把該基因歸為sPLA2-III亞家族, 定名為Ccpla2g3c。5個Ccpla2g3s都由信號肽、中心結(jié)構(gòu)域、較長的C端和N端組成, 且中心結(jié)構(gòu)域中均存在10個半胱氨酸,可形成5對二硫鍵, 具有保守的sPLA2-III特征序列[1],表明本文挖掘到的是sPLA2-III亞家族成員。
5個Ccpla2g3s基因位于獨立的連鎖群上, 而二倍體硬骨魚類如斑馬魚、青鳉等在基因組中均存在3個pla2g3基因:pla2g3a、pla2g3b、pla2g3c基因, 表明鯉是四倍體魚[26]。魚類pla2g3s基因的同源性、系統(tǒng)發(fā)育和線性分析結(jié)果支持: 雀鱔(2R魚)中pla2g3a(b)經(jīng)過3R過程, 在3R魚中形成了pla2g3a和pla2g3b兩個不同的基因, 但鯉科魚類在進化過程中pla2g3b變異較大而單獨成了一支, 最明顯的變異是鯉和斑馬魚的pla2g3b只有4個外顯子, 而其他魚類則和pla2g3a類似, 有7-8個外顯子; 3R魚中只挖掘到1個pla2g3c基因, 是由于其中1個染色體上丟失了1個pla2g3c。以上說明染色體加倍過程中基因突變現(xiàn)象比較普遍, 但具體機理值得深入研究揭示。
硬骨魚類及其祖先雀鱔(2R魚)中均存在sPLA2-III, 表明sPLA2-III是一類古老的基因, 雀鱔與硬骨魚類分離時間發(fā)生在3.5億年前[15], 在進化過程中基因發(fā)生了較大的變異, 導(dǎo)致染色體上sPLA2-III的下游基因不同。相比之下, 鯉與斑馬魚分化發(fā)生在1.28億年前, 且鯉特異的基因組復(fù)制發(fā)生在820萬年前[27], 不論在染色體水平還是基因同源性, 一致性都比較高, 顯示染色體、基因的突變概率是很低的。
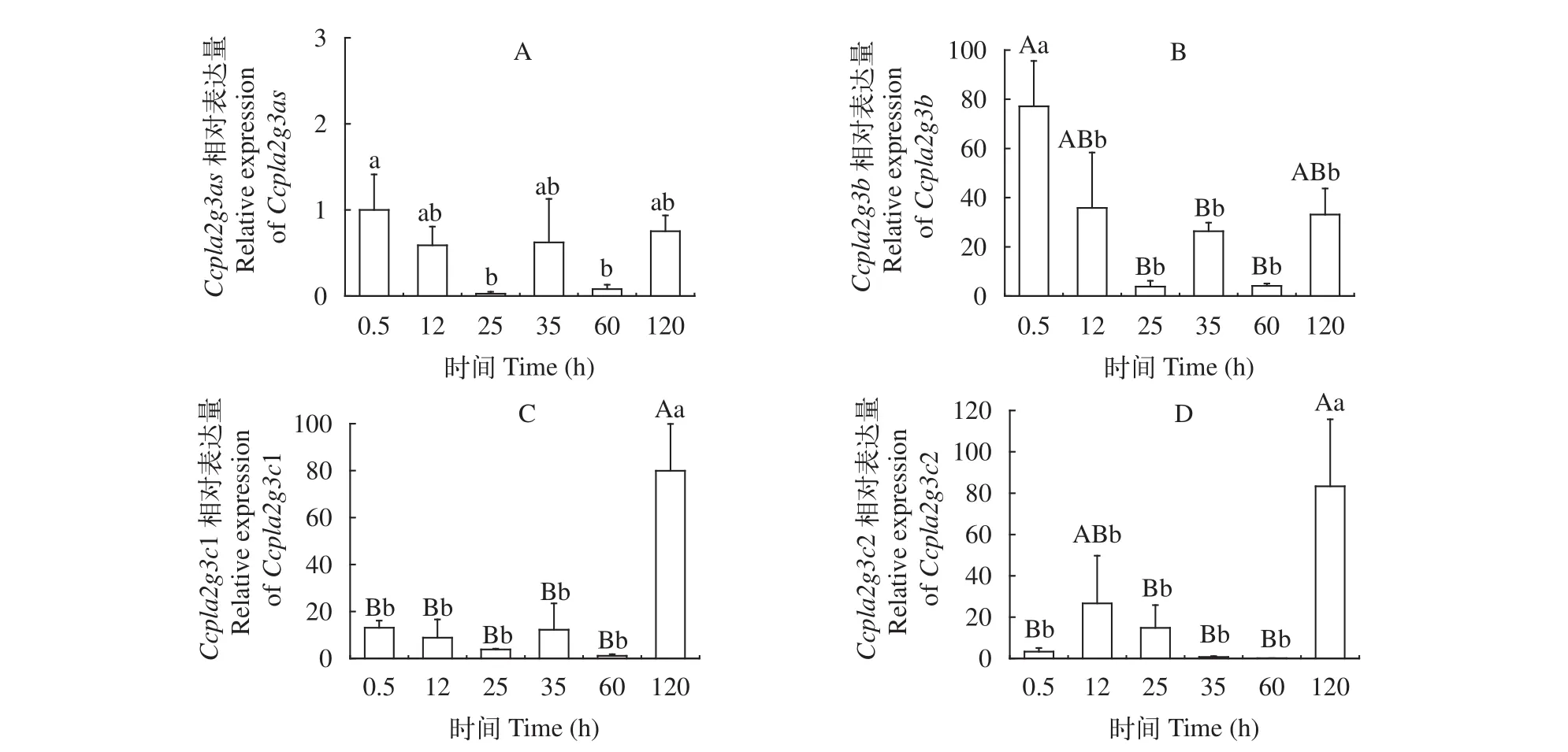
圖4 鯉sPLA2-III在胚胎發(fā)育階段的表達(dá)Fig. 4 The relative expression of sPLA2 group III genes during embryo development of common carp early
3.2 鯉sPLA2-III基因表達(dá)特征
sPLA2-III參與磷脂的降解和代謝[2], 存在于各組織器官中, 在本實驗中,Ccpla2g3as、Ccpla2g3b和Ccspla2g3cs呈現(xiàn)出了不同的表達(dá)模式,Ccpla2g3as在肝臟高表達(dá), 性腺中Ccpla2g3b表達(dá)量最豐富, 而Ccspla2g3cs則在腦中有明顯表達(dá), 很好地顯示了同源基因在進化過程中的表達(dá)分化; 進一步, 在鯉兩個旁基因中, 表達(dá)也存在著差異, 如Ccspla2g3c2在腸中的表達(dá)水平明顯高于Ccspla2g3c1 (P<0.05) (由于不在同一圖中差異標(biāo)識沒顯示)。推測同源基因功能的分化可能最早表現(xiàn)在組織表達(dá)差異。Ccpla2g3s在胚胎發(fā)育過程的表達(dá)特征, 和在成魚中的組織表達(dá)特征出現(xiàn)了很好的一致性,Ccpla2g3as在鯉整個受精卵階段表達(dá)量均較低, 這可能是因為肝臟組織在受精卵階段的發(fā)育分化不完善; 在鯉胚胎發(fā)育過程中,Ccpla2g3b在受精0.5h表達(dá)量最高, 隨后呈現(xiàn)下降的趨勢, 與該基因在性腺中的高表達(dá)量一致,表明可能與性腺脂肪代謝有密切關(guān)系, 另外推測受精卵中的高表達(dá)很可能是母源表達(dá), 并且和小鼠sPLA2-III在性腺組織中呈高表達(dá)[7]有一致性;Ccspla2g3cs在受精120h呈現(xiàn)高表達(dá), 可能與受精卵早期腦和腸道發(fā)育不完善有關(guān), 盡管該基因是哺乳動物PROCA1的同源基因, 但和PROCA1主要在人精巢中表達(dá)不一致[28], 推測該基因功能可能存在著一定的差異。研究表明磷脂酶A2在人的炎癥反應(yīng)中有著重要作用[29,30], 因此, 今后有必要進一步確定Ccpla2g3s在鯉炎癥中的生物學(xué)作用。
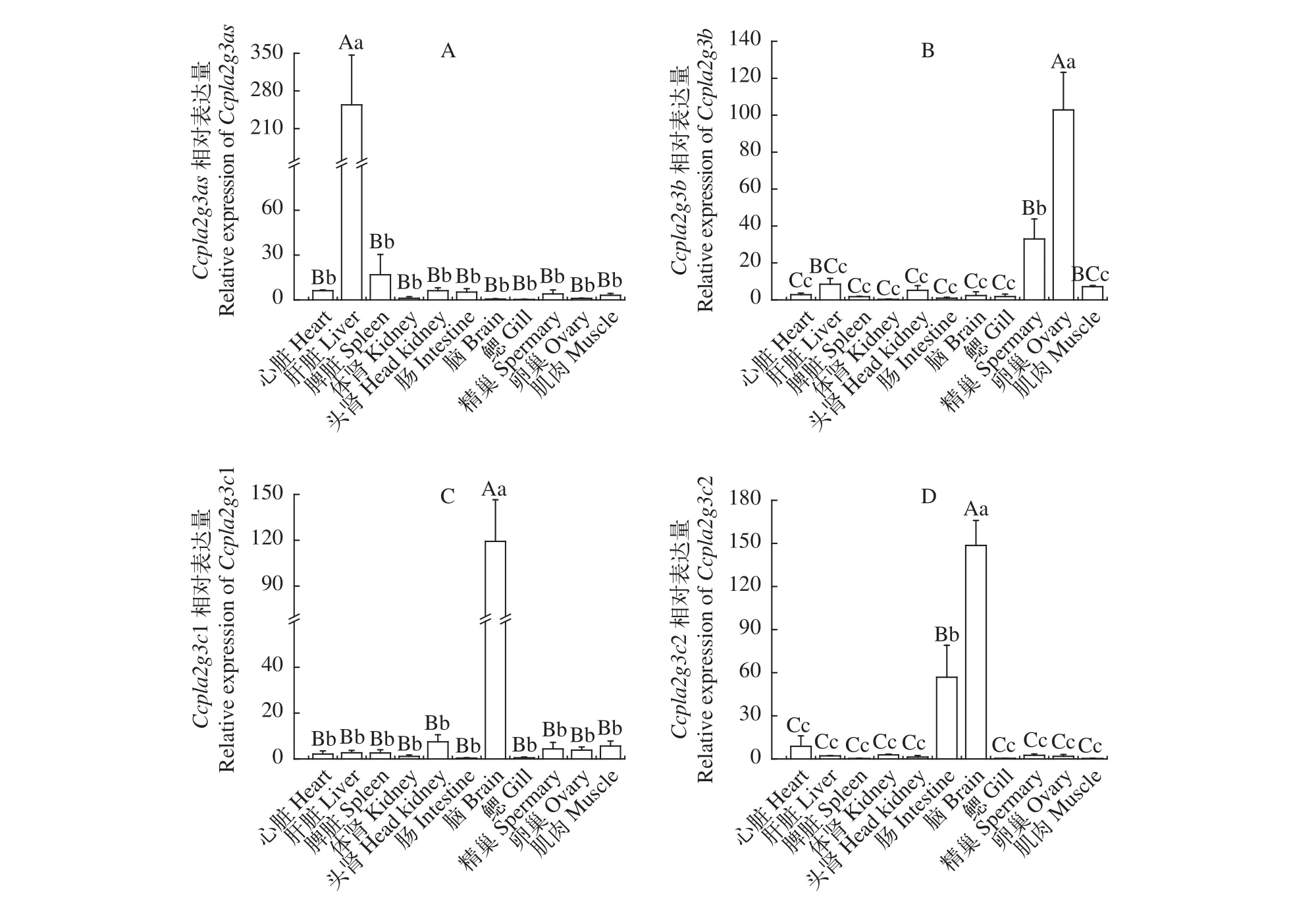
圖5 sPLA2-III在鯉成魚各組織的相對表達(dá)量Fig. 5 Relative expression of sPLA2-III in different tissues of common carp
猜你喜歡
遼寧教育(2022年19期)2022-11-18 07:20:42
公民與法治(2022年5期)2022-07-29 00:47:28
汽車實用技術(shù)(2022年9期)2022-05-20 05:51:26
教學(xué)考試(高考物理)(2021年5期)2021-11-08 10:31:22
歷史教學(xué)問題(2021年4期)2021-11-05 07:02:34
中醫(yī)眼耳鼻喉雜志(2021年1期)2021-07-22 07:38:14
裝備制造技術(shù)(2020年11期)2021-01-26 00:39:12
中國公共安全(2017年11期)2017-02-06 05:28:08
電測與儀表(2016年7期)2016-04-12 00:22:18
燕山大學(xué)學(xué)報(2015年4期)2015-12-25 02:19:49
