方格星蟲dmrt1基因的克隆和表達分析
2019-11-26 02:17:12李文華吳雅琴馬國興袁銘瑞許瑞安
水生生物學報 2019年6期
李文華 吳雅琴 馬國興 袁銘瑞 許瑞安
(華僑大學生物醫(yī)學學院, 分子藥物教育部工程研究中心, 福建省分子醫(yī)學重點實驗室, 廈門市海洋與基因工程藥物重點實驗室, 廈門 361021)
方格星蟲(Sipunculus nudus)又名裸體方格星蟲或光裸星蟲, 俗稱“沙蟲”, 隸屬于星蟲動物門, 方格星蟲綱, 方格星蟲科, 方格星蟲屬[1], 是一種名貴的海產珍品。目前, 關于方格星蟲繁殖生物學的研究甚少。方格星蟲為體外受精, 雌雄異體, 性成熟最短時間為11個月, 其生殖腺位于收吻肌基部, 腺體細小, 肉眼難以識別, 僅在排卵期明顯[2,3]。全年都可以在其體腔中觀察到生殖細胞, 生殖細胞的發(fā)育隨季節(jié)的變化而變化[4]。星蟲動物的配子發(fā)生過程比較特別, 原生殖細胞在發(fā)育早期便以單細胞或細胞團的形式從性腺脫落進入到體腔液中繼續(xù)發(fā)育至成熟, 發(fā)育成熟后進入腎管[3,4]。方格星蟲具有1對細長的呈圓筒狀的腎管結構, 腎管一般作為排泄器官, 負責排出體腔中的代謝廢物[5]。在繁殖季節(jié), 腎管兼作生殖道, 負責收集體腔中的精卵并使之獲能。產卵前, 體腔內成熟的生殖細胞通過內腎孔進入到腎管。產卵時, 成熟的生殖細胞通過外腎孔排出體外[6,7]。與大多數(shù)無脊椎動物的繁殖過程不同, 星蟲的繁殖生物學具有一定的特殊性, 其配子只有經(jīng)腎管收集并停留一段時間進行最后的“激活包裝”才能完全成熟, 才具有受精能力[4,6,8—11]。
DMRT (Double-sex and mab3-related transcription factor)基因家族, 在動物性別決定和分化過程中發(fā)揮著重要的調控作用[12]。該家族共同的特征是含有保守的鋅指DNA結合結構域(Doublesex and mab-3, DM domain), 其中, DMRT基因家族中的dmrt1在性別決定、分化和器官功能維持上發(fā)揮著重要的作用, 已在哺乳動物、禽類、兩棲爬行類及硬骨魚類等發(fā)現(xiàn)[12—16]。人類(Homo sapiens)dmrt1定位于9號染色體, 該基因的缺失會導致精巢發(fā)育障礙[17], 在XY雄性小鼠(Mus musculus)中的研究結果顯示, 敲除dmrt1基因后嚴重影響睪丸分化和精子形成[18]。鳥類dmrt1對于精巢的分化也起著至關重要的作用, 且高表達的dmrt1基因決定了鳥類的性別[19]。非洲爪蟾(Xenopus laevis)中的一個位于W染色體上的dmrt1的復制本,DM-W, 被證明為非洲爪蟾的性別決定基因,DM-W的表達能夠阻止精巢的分化, 從而導致卵巢的發(fā)生[20,21]。青鳉(Oryzias la-tipes)dmrt1基因在Y染色體上的一個復制本Dmy(又叫Dmrt1y)是一個雄性特異的性別決定基因, 能促使雄性發(fā)生也是促使雄性發(fā)生所必須的基因[22,23]。與脊椎動物相比, 無脊椎動物dmrt1基因的研究報道尚少, Jennifer等[24]在無脊椎動物東部大螯蝦(Sagmariasus verreauxi)中克隆到定位于Y染色體上的一個DMRT基因的復制本iDMY。dmrt1作為不同物種間保守的性別相關基因, 尚沒有研究報道其在方格星蟲中的研究情況。本研究旨在克隆得到在海洋無脊椎動物方格星蟲生殖發(fā)育中的重要功能基因dmrt1的全長cDNA序列, 分析它的序列特征、進化關系和表達特征。
1 材料與方法
1.1 實驗材料
1—1.5年齡方格星蟲(體長約15 cm)購買于廈門第八海鮮市場。抽取方格星蟲體腔液鑒定雌雄后, 取體腔液、腎管、神經(jīng)節(jié)、肌肉、翻吻肌和腸道組織用液氮速凍后, 置于-80℃保存, 用于成體組織實時熒光定量PCR。
1.2 方格星蟲SMART cDNA文庫構建
取方格星蟲雄性個體的體腔液, 使用TRIzol(TaKaRa)提取總RNA, 然后用SMART cDNA合成試劑盒(Clontech)構建SMART cDNA文庫。總RNA的提取和SMART cDNA的合成參照產品說明書。
1.3 Dmrt1基因的cDNA序列擴增
根據(jù)前期實驗獲得的部分dmrt1序列設計RACE擴增引物(表 1), 然后使用以下程序進行PCR擴增: 95℃變性15s, 60℃退火30s, 72℃延伸2min, 共36個循環(huán), 72℃加尾15min。將獲得的PCR產物上樣于1%的瓊脂糖凝膠中, 取適量目標條帶切膠純化后的產物與pMD18-T載體連接, 4℃過夜處理后轉入大腸桿菌感受態(tài)中, 篩選含有陽性插入片段的菌落進行測序驗證和分析。
1.4 Dmrt1基因的序列分析
在NCBI (http://www.ncbi.nlm.nih.gov/)數(shù)據(jù)庫中, 利用方格星蟲DM結構域檢索近緣物種DM結構域的氨基酸序列, 使用MUSCLE軟件對所有序列進行對位排列, 并用BioEdit軟件對排列不整齊的序列進行人工校正。選擇MEGA 7.0軟件, 基于Jones-Taylor-Thornton (JTT)模型, 將對位排列整齊的序列構建鄰接法(Neighbor-Joining, NJ)系統(tǒng)發(fā)育樹, 系統(tǒng)樹進行1000次自展重復, 其中大于50%的枝被認為是可信的。
1.5 實時熒光定量PCR
分別提取雌雄方格星蟲體腔液、神經(jīng)節(jié)、腸道、肌肉、腎管和翻吻肌的總RNA。用PrimeScript?RT reagent Kit With gDNA Eraser (TaKaRa)試劑盒合成單鏈cDNA, 具體操作步驟參見產品說明書。選擇方格星蟲β-actin作為內參基因, 使用ChamQ SYBR qPCR Master Mix (Vazyme)進行實時熒光定量PCR擴增, 具體操作程序和方法參見產品說明書,所用引物序列見表 1。

表1 引物序列Tab. 1 Primer sequences
1.6 原位雜交
選取方格星蟲dmrt1的部分cDNA序列設計原位雜交引物, 為了獲得含有T7啟動子的目標序列,我們分別在上下游引物的5′端插入T7啟動子序列TAATACGACTCACTATAGGGAGA, 以獲得方格星蟲dmrt1正反義探針, 具體參見表 1, 然后通過常規(guī)PCR擴增的方式獲得含有T7啟動子的方格星蟲dmrt1基因片段。接下來, 使用T7聚合酶(Roche)進行體外轉錄, 具體操作方法參見產品說明書。分別取雌雄方格星蟲體腔液涂布于載玻片上, 37℃烘干1h, 然后將切片進行蛋白酶消化、多聚甲醛固定、RNA探針雜交、洗脫、封閉、抗體孵浴、染色及顯色等步驟[25]。
1.7 統(tǒng)計分析
熒光定量的統(tǒng)計數(shù)據(jù)用平均值±標準差(Mean±SD)來表示。對各實驗組的熒光定量數(shù)據(jù)進行單因素方差分析(ANOVA), 取P<0.05為顯著性差異閾值, 差異分析使用SPSS 22.0軟件進行。樣品間基因表達量數(shù)據(jù)圖選用GraphPad Prism 5.0軟件進行繪制。
2 結果
2.1 Dmrt1基因的序列特征分析
方格星蟲dmrt1全長cDNA含有1509 bp (Gen-Bank: MK182259), 其中開放閱讀框的長度為615 bp,5′-UTR的長度為145 bp, 3′-UTR的長度為749 bp, 共編碼204個氨基酸。同時, 我們擴增出的方格星蟲dmrt1全長含有多腺苷酸化加尾信號AATAAA, 加尾信號位于poly (A)尾上游的18 bp處。
對方格星蟲dmrt1的氨基酸序列進行保守結構域分析, 顯示其與Dmrt家族成員具有極其相似且高度保守的DM結構域, 將方格星蟲DM結構域與已報道的其他物種DM結構域進行多重序列比對, 比對結果顯示方格星蟲DM結構域與海葵(Exaiptasia pallida)的DM結構域一致性最高, 高達81%。此外,方格星蟲與其他海洋軟體動物, 如牡蠣(Crassostrea gigas)、珠母貝(Pinctada margaritifera)的DM結構域的相似性也可達到72%, 且與脊椎動物人、小鼠、非洲爪蟾及斑馬魚(Danio rerio)的DM結構域的一致性分別為72%、72%、70%及70%。
為了確定方格星蟲dmrt1基因的進化關系, 我們構建了系統(tǒng)進化樹。進化樹的拓撲結構顯示方格星蟲dmrt1基因與數(shù)據(jù)庫中已知的脊椎動物或海洋軟體動物都沒有表現(xiàn)出明顯的聚集現(xiàn)象(圖 1),暗示方格星蟲dmrt1基因與我們找到的這些物種的親緣關系相對較遠。目前, 關于星蟲或其近緣物種的dmrt1基因的相關研究尚鮮有報道。
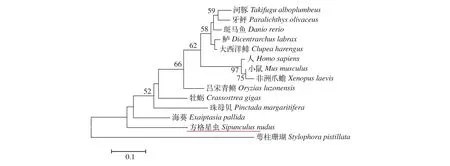
圖1 基于NJ法構建的不同物種DMRT1的系統(tǒng)發(fā)育樹Fig. 1 Phylogenetic tree of DMRT1 in species based on NJ method
2.2 方格星蟲dmrt1在雄性個體的體腔液中高表達
我們采用熒光定量PCR方法研究了dmrt1在不同組織(體腔液、神經(jīng)節(jié)、腸道、肌肉、腎管和翻吻肌)中的表達差異。方格星蟲dmrt1僅表達于雄蟲體腔液中, 在所檢測的其他組織包括神經(jīng)節(jié)、腸道、肌肉、腎管和翻吻肌中沒有表達(圖 2), 且在所檢測的雌性個體的各個組織(體腔液、神經(jīng)節(jié)、腸道、肌肉、腎管和翻吻肌)皆不表達。
為了進一步研究dmrt1在成體方格星蟲體腔液中的表達圖式, 我們抽取方格星蟲雌雄個體的體腔液, 采用方格星蟲dmrt1正、反義探針進行了體腔液涂片原位雜交實驗。結果顯示, 雜交信號主要出現(xiàn)在雄性個體體腔液, 且主要集中在精子團附近的滋養(yǎng)細胞中(圖 3A), 而在雌性個體體腔液中沒有檢測到雜交信號(圖 3B), 同時使用正義探針在雄性體腔液中也沒有檢測到雜交信號(圖 3C)。
3 討論
Dmrt1基因作為一個保守的性別相關基因, 廣泛參與哺乳類、爬行類、兩棲類和魚類的性別發(fā)育和分化過程, 在性別決定和生殖器官的功能維持等方面發(fā)揮著重要作用, 但該基因在不同進化地位和不同性別決定機制的物種中的表達特性及功能不盡相同[12—16]。本研究從方格星蟲體腔液中克隆得到dmrt1基因, 生物信息學分析顯示dmrt1基因具有保守的DM結構域, 同時通過熒光定量PCR, 我們報道了該基因主要表達于方格星蟲雄性個體的體腔液中。
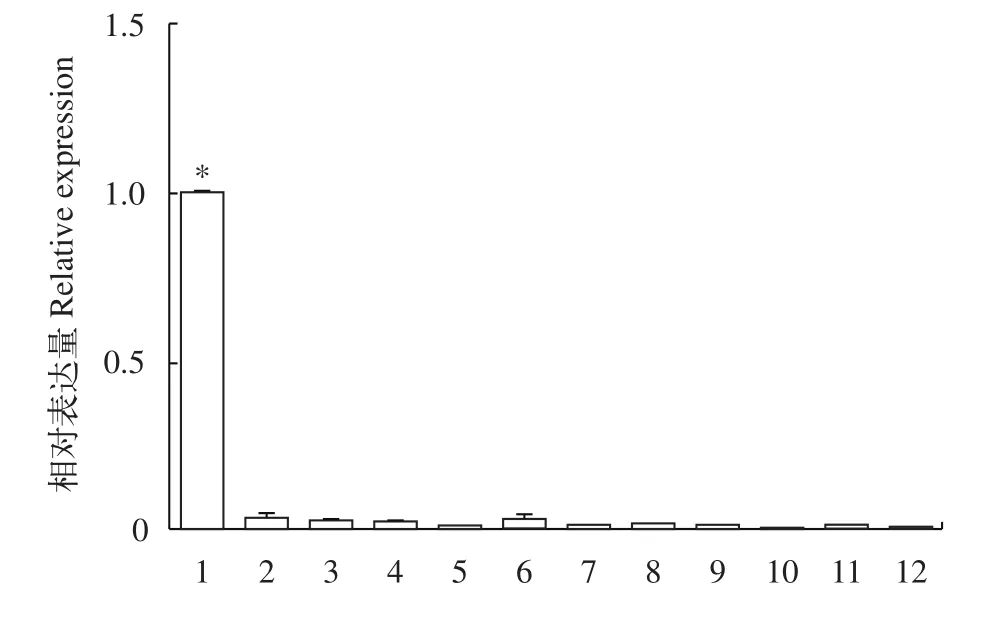
圖2 方格星蟲dmrt1在不同組織中的熒光定量結果Fig. 2 The relative expression of dmrt1 in adult tissues of Sipunculus nudus
DM結構域最早鑒定于果蠅中, 從無脊椎動物到脊椎動物DM結構域表現(xiàn)出高度的進化保守性[26]。方格星蟲DMRT1含有DM結構域, 表明我們克隆的dmrt1基因符合DMRT家族成員的共有特征。生物信息學分析結果顯示方格星蟲DM結構域與海洋軟體動物, 如海葵、牡蠣及珠母貝的一致性可達72%以上, 與脊椎動物人、小鼠和斑馬魚的DM結構域一致性也能達到70%以上, 說明方格星蟲DMRT1具有一定的保守性。
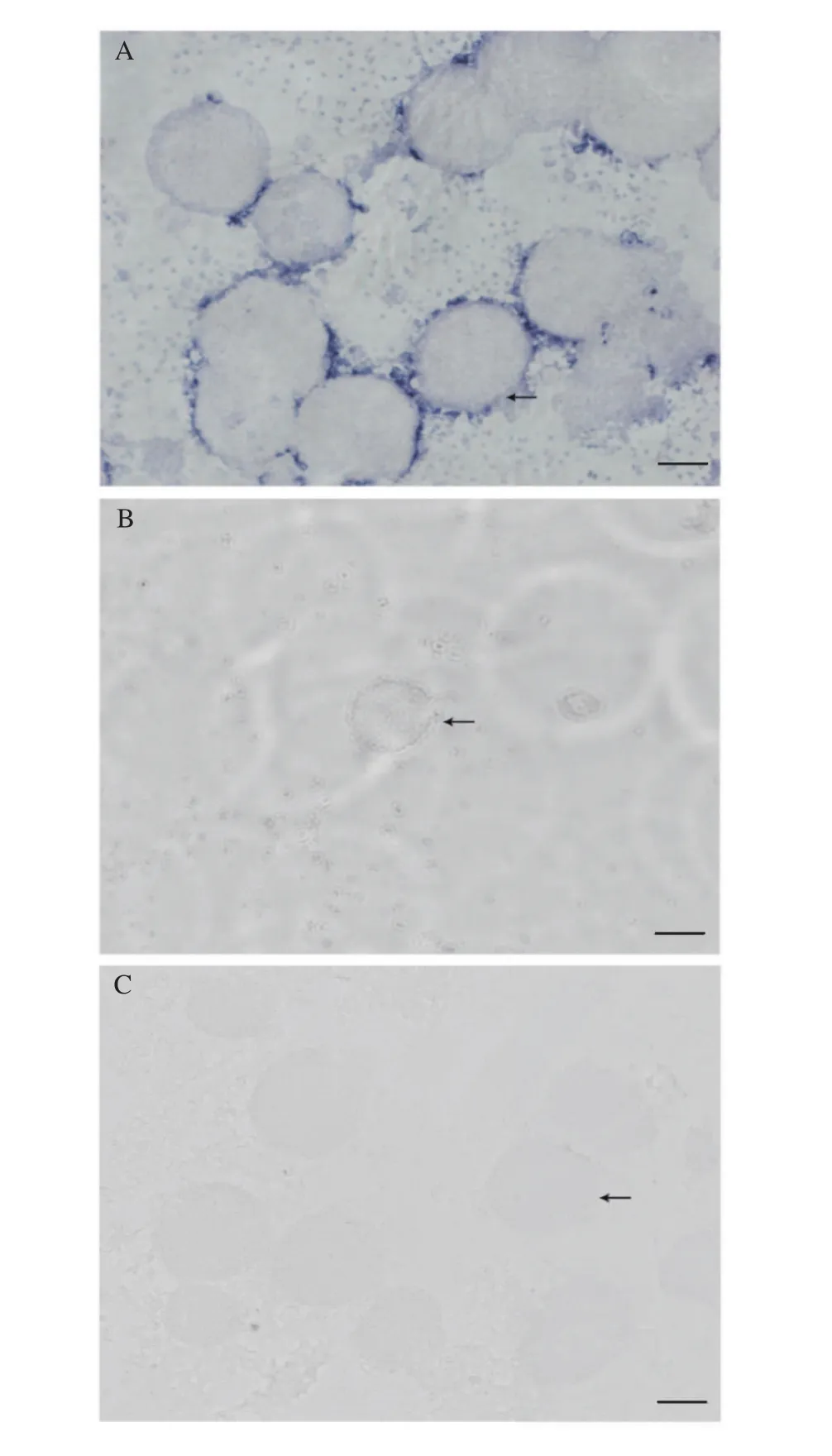
圖3 方格星蟲dmrt1 mRNA在雌雄體腔液中的細胞定位Fig. 3 Cellular distribution of dmrt1 mRNA in Sipunculus nudus coelomic fluid
早有報道稱星蟲是一類有體腔、兩側對稱、身體不分節(jié)及外型像蟲的古老的海洋無脊椎動物,化石證據(jù)表明星蟲至少出現(xiàn)于后寒武紀, 在5.2億年里, 星蟲的形態(tài)結構幾乎沒有發(fā)生變化[27]。系統(tǒng)進化樹的結果顯示方格星蟲dmrt1基因與現(xiàn)有數(shù)據(jù)庫中的部分脊椎動物和海洋軟體動物都沒有聚為一支(圖 1), 我們認為其主要原因可能是星蟲類動物作為一種古老的海洋軟體動物, 尚沒有與其遺傳距離較近的物種報道克隆出dmrt1基因, 且星蟲類物種本身的繁殖生物學研究甚少, 已有研究也僅集中在生殖細胞形態(tài)、分類及組織學研究等方面[28—30]。綜上所述, 本文開啟了方格星蟲類動物在性別決定及分化相關基因研究, 為進一步深入研究打下基礎。
在已經(jīng)檢測的多數(shù)物種中,dmrt1都在雄性性腺或者雌性向雄性轉變的性腺中高表達[13]。方格星蟲dmrt1在6個所檢測的成體(雌蟲和雄蟲)組織中, 主要表達于雄蟲體腔液中(圖 2)。星蟲動物的原生殖細胞在發(fā)育早期以單細胞或細胞團的形式從肉眼難以識別的性腺上脫落進入到體腔液中繼續(xù)發(fā)育, 形成的生殖細胞附著于具有運動能力的滋養(yǎng)細胞基部[3,31]。方格星蟲雄性生殖細胞一般以精細胞團形式存在, 卵細胞以單個游離形式存在[31]。原位雜交實驗結果顯示方格星蟲dmrt1的雜交信號主要出現(xiàn)在精子團附近的滋養(yǎng)細胞中(圖 3A), 蘭寶國等[4]認為滋養(yǎng)細胞帶著生殖細胞一起運動, 從而有利于生殖細胞從體腔液中攝取營養(yǎng)。哺乳動物精巢中有一類重要的細胞, 即支持細胞(Sertoli cells), 它為發(fā)育中的精子提供保護和營養(yǎng)[32], 我們認為方格星蟲雄性體腔液中的滋養(yǎng)細胞可能與哺乳動物精巢中的支持細胞具有類似的功能, 因此推測方格星蟲dmrt1可能與其雄性決定和精子發(fā)生相關聯(lián), 而具體生物學功能仍有待進一步的研究。
