黃顙魚和雜交黃顙魚“黃優1號”形態及性腺發育的比較
2019-11-26 02:17:14胡偉華郭穩杰樊啟學
水生生物學報 2019年6期
關鍵詞:生長
胡偉華 丹 成 郭穩杰 樊啟學 梅 潔
(華中農業大學水產學院, 武漢 430070)
黃顙魚(Pelteobagrus fulvidracoRichardson)是我國一種重要的小型底棲經濟魚類, 廣泛分布于全國各流域水體中, 因其肉質細嫩、肉味鮮美、無肌間細刺, 受到廣大消費者的青睞。由于黃顙魚雄性比雌性生長快, 我國學者開拓出了一條X和Y染色體連鎖標記輔助的全雄魚培育技術路線, 并培育出新品種全雄黃顙魚“全雄1號”[1—6]。全雄黃顙魚顯著提高了養殖經濟效益, 其大范圍推廣極大推動了黃顙魚產業的發展。近年來, 親本超雄魚繁育系經過多代自交之后發生了退化, 自交系在生長和抗病等性能方面呈現減弱的趨勢, 此時需要對全雄黃顙魚進行品種改良或開發出其他黃顙魚新品種。根據養殖情況調查和研究發現, 雜交黃顙魚(黃顙魚P.fulvidraco♀×瓦氏黃顙魚P. vachelli♂)形態特征與黃顙魚接近, 生長速度優于黃顙魚[7—9]。
雜交育種是遺傳育種中最經典的方法之一, 不同種間生物的雜交, 有可能迅速和顯著地提高雜種后代的存活率、生長速度等經濟性狀, 繼而可以經培育而成新品種[7]。迄今為止, 已通過不同種類雜交培育出了一些在生長性能上具有顯著優勢的雜交魚類品種, 例如雜色鮑“東優1號”(臺灣雜色鮑Haliotis diversicolor♀和日本雜色鮑Haliotis diversicolor♂)、湘云鯽(白鯽Carassius auratus cuvieri♀和異源四倍體鯉鯽Allotetraploid hybrids♂)以及雜交鱧“杭鱧1號”(珠江水系斑鱧Channa maculata♀和錢塘江烏鱧Channa argus♂)等[10]。華中農業大學黃顙魚種質資源與品種改良團隊采用雜交育種的方法, 選擇梁子湖水域采捕并經連續3代選育的黃顙魚為母本、長江河段中采捕并經連續2代選育的瓦氏黃顙魚為父本, 再經人工雜交獲得F1代即為雜交黃顙魚“黃優1號”。與黃顙魚相比,一齡“黃優1號”生長速度和成活率均提高30%以上。據市場調查反應, “黃優1號”的雌雄生長速度和性腺發育情況較黃顙魚也具有差異, 對“黃優1號”的基礎研究有待進一步挖掘。
由于黃顙魚和雜交黃顙魚的形態特征非常接近, 養殖過程中會存在混雜在一起的現象, 引起從事水產一線的工作者們對于雜交黃顙魚(黃顙魚♀×瓦氏黃顙魚♂)的生長性能、外觀形態等的爭議。最近, 我們開發了一對DNA分子標記能穩定、高效識別黃顙魚、瓦氏黃顙魚及其雜交種,為研究它們之間的差異奠定了基礎[11]。雜交后代不育現象在動植物中普遍存在, 雜交黃顙魚后代的遺傳能力以及生長優勢是評價其能否成為養殖新品種的重要指標之一。本研究以養殖時間10—34月齡黃顙魚和雜交黃顙魚“黃優1號”為研究對象, 比較分析黃顙魚和“黃優1號”的形體指標和性腺發育情況。
1 材料與方法
1.1 材料
2014年7月、2015年7月于湖北省武漢市江夏區魯湖養殖基地進行黃顙魚和雜交黃顙魚“黃優1號”的繁育, 待完成飼料訓食后, 選取6個相同養殖條件、面積為3333.33 km2(5畝)的池塘進行養殖試驗, 投放規格為0.26—0.35 g/尾, 養殖密度均為12000 尾/畝, 黃顙魚和“黃優1號”投喂量均相同。于2016年6月、2017年6月在池塘中隨機捕撈黃顙魚和“黃優1號”各60尾, 進行采樣、檢測和數據處理。
1.2 形體指標測量
將實驗魚置于200 mg/L MS-222(間氨基苯甲酸乙酯甲磺酸鹽)的環境中麻醉, 于解剖盤上測量體長(BL)、體重(BW)、性腺重(GW)和性腺成熟系數(GSI, 性腺體重比)。
1.3 魚體解剖及形態學觀察
將麻醉后的實驗魚置于解剖盤上, 剪去腹部肌肉, 去除內臟團, 用濾紙吸取流出的血液, 觀察實驗魚性腺發育情況并于解剖盤上拍照記錄。然后, 取出完整的性腺置于4%多聚甲醛中固定。經梯度酒精脫水, 二甲苯透明, 石蠟包埋, 連續切片厚度為 8 μm。將石蠟切片用常規蘇木精-伊紅染色(HE)染色, 用Axio Imager A2顯微拍照觀察。
1.4 計算機輔助精子分析系統(CASA)
實驗魚麻醉后剪開腹部肌肉, 取下完整的性腺并置于潔凈的定性濾紙上, 去除附著在性腺上的組織液及其他物質, 以免激活精子。將黃顙魚和“黃優1號”精巢按照質量體積比=1:2添加精子保存液(精子保存液配方: 63 mmol/L NaCl、19 mmol/L KCl、1.3 mmol/L CaCl2、4.7 mmol/L MgSO4·7H2O、2.5 mmol/L NaHCO3, pH 7.4)[12], 在潔凈的勻漿器中均質, 備用。
精子活力和運動參數利用計算機輔助精子分析系統(CASA)檢測分析, 設定60幀/s, 選擇10×相位差進行分析每個樣品中的精子移動[13—15]。用0.45%的NaCl溶液激活測定, 每個樣品至少隨機檢測3次。精子活力的指標可以通過多個參數來顯示,如軌跡速度(VCL), 也稱曲線速度, 即精子頭部沿其實際行走曲線的運動速度; 平均路徑速度(VAP), 精子頭沿其空間平均軌跡的運動速度, 這種平均軌跡是計算機將精子運動的實際軌跡平均后計算出來的; 直線運動速度(VSL): 也稱前向運動速度, 即精子頭部直線移動距離的速度; 直線性(LIN): 也稱線性度, 為精子運動曲線的直線分離度, 即VSL/VCL;鞭打頻率(BCF): 也稱擺動頻率, 即精子頭部跨越其平均路徑的頻率。
1.5 “黃優1號”繁殖能力測試
選取同一養殖基地池塘中養殖22月齡的“黃優1號”與黃顙魚作為親魚開展人工催產試驗。挑選形體、體色正常, 體質健壯, 無疾病、無傷病和畸變的“黃優1號”和黃顙魚雄魚各10尾, 黃顙魚雌魚20尾。在水溫達到23—26℃時, 進行“黃優1號”雄魚和黃顙魚雌魚人工催產試驗測試“黃優1號”繁殖能力, 并以黃顙魚作為對照。
催產方式: “黃優1號”和黃顙魚雄魚不注射催產藥物; 雌性黃顙魚采用兩次注射(胸鰭基部注射),催產藥物為地歐酮(DOM)、魚用絨毛膜促性腺激素(HCG)和促黃體激素釋放激素(LRH-A2)3種藥物配伍; 注射計量為第一針: LRH-A2 10—16 μg +HCG 100—200 IU/kg; 第二針: LRH-A2 16—20 μg +HCG 1200—2000 IU/kg + DOM 5—8 mg/kg, 兩針間距時間為8—12h。
采用“半干法”進行黃顙魚人工授精, 達到效應時間前2—3h檢查親魚動態, 待60%—70%的雌性黃顙魚個體都能達到順利完成人工擠卵時, 開始人工收卵。將收集到的卵混勻, 按照重量分置6個玻璃培養皿 (HY-1、HY-2、HY-3;HF-1、HF-2、HF-3)中。HY-1、HY-2、HY-3 用“黃優1號”精液進行人工授精, HF-1、HF-2、HF-3用黃顙魚精液進行人工授精。完成人工授精的卵置于玻璃平皿中充氣孵化。受精卵發育至原腸胚期后計算卵的總數和受精卵數量, 然后繼續充氣孵化, 魚苗出膜后計算魚苗出膜數量[16]。
1.6 數據處理
采用SPSS18.0統計軟件進行實驗數據的統計學分析, 采用單因素方差分析(One-way ANOVA)進行各組數據的組間差異分析,P<0.05表示差異具有統計學意義。
2 結果
2.1 黃顙魚和“黃優1號”的形體指標測量
在針對生產實踐中, 雜交黃顙魚與黃顙魚從外觀形態不易區分的問題, 本實驗在運用DNA分子標記識別黃顙魚、瓦氏黃顙魚及其雜交種的基礎上,通過對比形體指標及鰭式差異, 發現黃顙魚胸鰭前緣和后緣均有鋸齒狀凸起, 而“黃優1號”胸鰭前緣光滑, 僅在后緣存在鋸齒狀凸起(圖 1)。這一性狀提供了簡便快速區分雜交黃顙魚“黃優1號”和黃顙魚的方法。
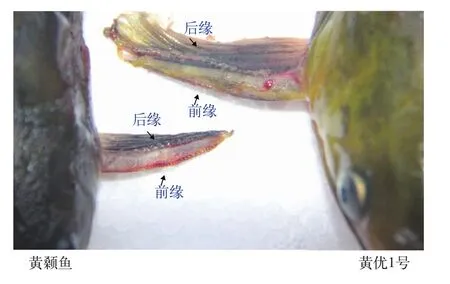
圖1 黃顙魚與“黃優1號”胸鰭前后緣的差異Fig. 1 Phenotypic differences of anterior and posterior margin of pectoral fin between yellow catfish and hybrid yellow catfish“Huangyou No. 1”
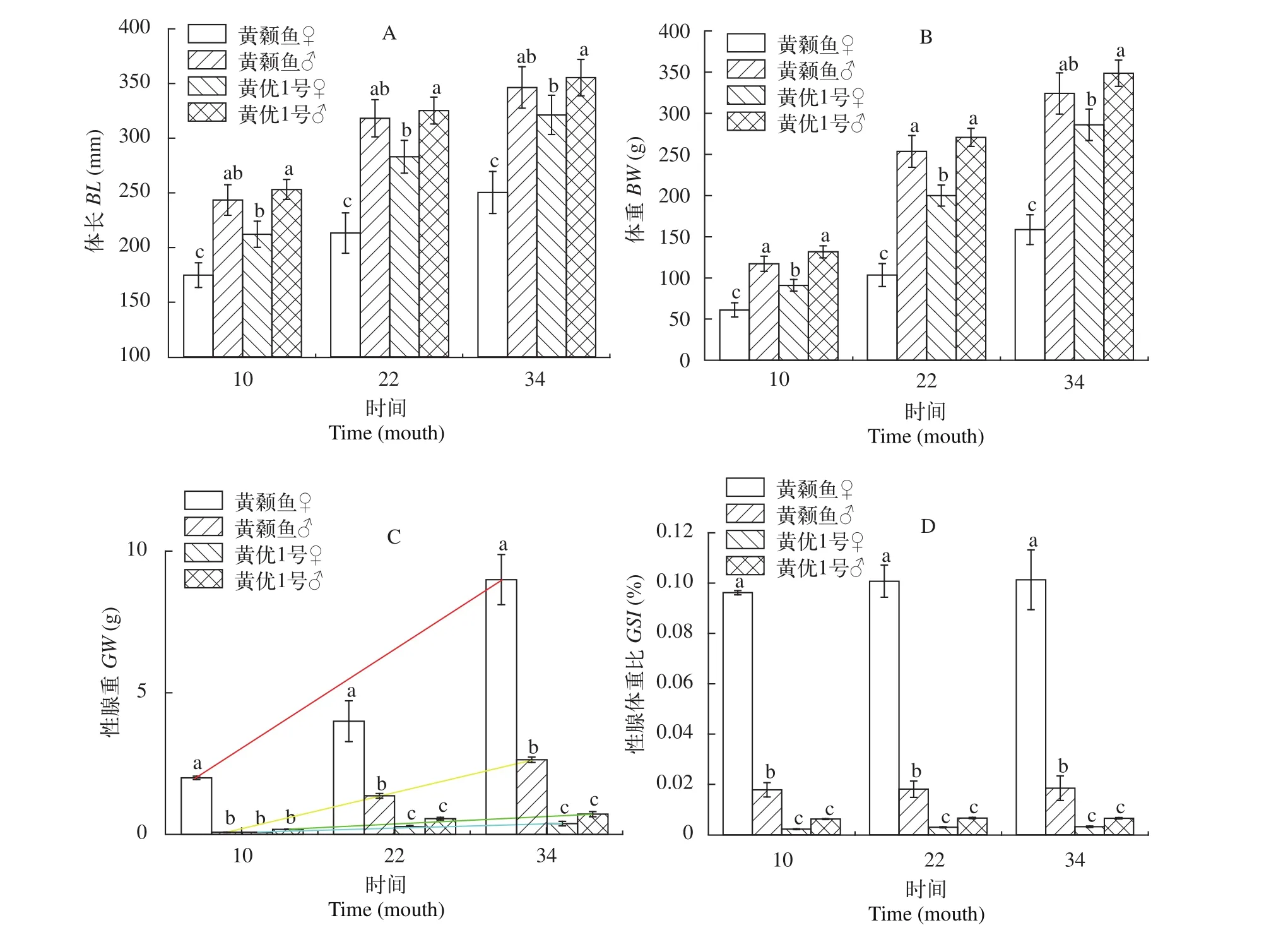
圖2 黃顙魚和“黃優1號”形體指標Fig. 2 The physical indicators of yellow catfish and hybrid yellow catfish “Huangyou No. 1”
養殖1年齡為養殖黃顙魚可上市的初步規格,2年齡為大規格黃顙魚, 3年齡可更好地觀察比較黃顙魚性腺發育情況, 故本實驗選取10月、22月和34月齡黃顙魚和“黃優1號”進行了形體指標檢測(圖 2)。在10—34月齡的對比試驗中, 選擇相同養殖月份的形體數據做統計分析, 發現雄性黃顙魚與“黃優1號”在體長和體重上差異不顯著(P>0.05), 雌性黃顙魚與“黃優1號”則具有顯著差異(P<0.05)(圖 2A), 說明“黃優1號”相較于黃顙魚雌雄生長差異縮小。另外, 10—34月齡“黃優1號”的平均體重比黃顙魚高28.33%—46.48%, 其中, 雄性黃顙魚10月齡、22月齡和34月齡平均體重比雌性分別高65.16%、145.51%和119.87%, 而雄性“黃優1號”平均體重比雌性分別高46.33%、49.73%和31.90%(圖 2B), 雌雄生長差異縮小。雄性“黃優1號”生長速度與雄性黃顙魚相近, 雌性“黃優1號”生長速度比黃顙魚提升明顯, 故“黃優1號”的整體生長速度是明顯提升的。同時, 雌雄生長差異縮小, 達到上市規格的比例較黃顙魚有所提高。本實驗的黃顙魚和“黃優1號”養殖條件和投喂量一致, 不同的養殖條件實驗結果也會略有不同, “黃優1號”的搶食和攝食能力均強于黃顙魚。
黃顙魚雌性的性腺重(GW)顯著高于雄性(P<0.05), 而“黃優1號”雌雄性腺重沒有顯著差異(P>0.05)。進一步比較兩者的性腺重量(GW)的增長率發現“黃優1號”性腺重量(GW)增長率顯著低于黃顙魚(圖 2C;P<0.05); 以性腺體重比(GSI)換算而來的成熟系數, 結果為黃顙魚成熟系數顯著高于“黃優1號”(圖 2D;P<0.05)。
2.2 黃顙魚和“黃優1號”的性腺形態學和組織學觀察
如圖 3中的黃顙魚性腺解剖圖所示, 10月、22月和34月齡黃顙魚卵巢正常發育, 有大量的卵子發生(圖3A—3C), 而“黃優1號”的卵巢為白色線狀組織呈現退化狀態(圖 3D—3F), 故“黃優1號”沒有卵子生成表現為明顯的雜交不育。10月、22月和34月齡黃顙魚精巢正常發育, 為乳白色的精小葉結構(圖 3G—3I), 而“黃優1號”的精巢為透明的精小葉結構和精巢退化狀態(圖 3J—3L), “黃優1號”的精巢結構需要通過組織學進一步分析。
對黃顙魚和“黃優1號”精巢組織切片進行HE染色分析發現, 黃顙魚在10月齡時精巢發育完全并且精小囊內存在大量精子(圖 4A), 而10月齡“黃優1號”精巢中精小囊僅存在空殼, 幾乎沒有精子存在(圖 4B)。22月齡“黃優1號”精巢中精小囊內發現少量精子存在(圖 4C), 34月齡“黃優1號”精巢中出現一些類似于精原細胞和精母細胞的細胞(圖 4D)。
2.3 計算機輔助精子分析系統(CASA)比較分析黃顙魚和“黃優1號”的精子質量
如圖 5所示, 黃顙魚的單位體積精子數量隨著年齡的增長呈現遞增趨勢, 大約為1×107—1×108個精子/mL, 大部分為運動的精子; 而“黃優1號”的精子數量顯著減少(P<0.05), 僅為1—5×103個精子/mL。同時, 相較于黃顙魚的運動狀態下精子數量占精子總數的83.00%—89.00%, “黃優1號”的活力精子數量占精子總數僅為17.61%—24.91%, 且靜止狀態下精子數量多于運動狀態下精子數量。
在醫學和動物學研究中, 計算機輔助精子分析系統(CASA)被廣泛用于評價精子的質量, VAP、VSL和VCL等參數與不育男性生育能力具有顯著的相關性, 是評判人類有無生育能力的重要參數[17,18]。“黃優1號”精子的VAP、VSL和VCL等參數均顯著低于黃顙魚(P<0.05), 但LIN(VSL/VCL)和BCF參數沒有顯著變化(P>0.05; 表 1)。這些參數表明“黃優1號”的運動狀態下精子的活力極其低下。
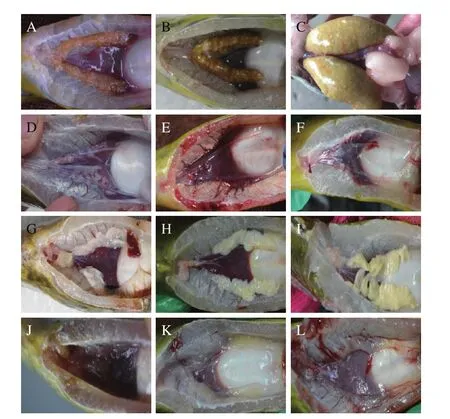
圖3 黃顙魚和“黃優1號”性腺解剖圖Fig. 3 Anatomy of the gonad of yellow catfish and hybrid yellow catfish “Huangyou No. 1”
2.4 “黃優1號”雄魚繁殖能力測試
如圖 6和表 2所示, “黃優1號”♂與黃顙魚♀(HY組)受精率和孵化率均為0; 而黃顙魚♂與黃顙魚♀(HF組)受精率最高為83.15%, 最低為71.74%,平均值為76.71%。通過胚胎發育觀察, HF組胚胎發育至2胞期時, 而HY組胚胎不能發育到2胞期。當HF組胚胎發育至原腸胚時期時, HY組胚胎都已經變白, 死亡。
經對比發現, 22月齡“黃優1號”雄魚不具備繁殖能力, 同時“黃優1號”雌魚未能形成囊狀卵巢, 不能進行人工催產, 故判斷“黃優1號”不具備繁殖能力。
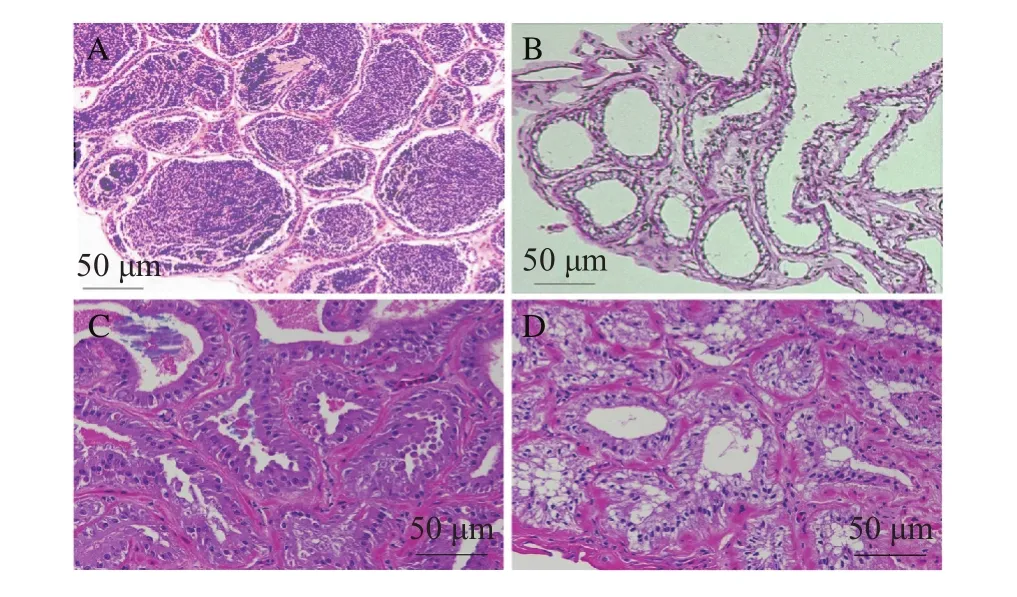
圖4 黃顙魚和“黃優1號”精巢組織切片HE染色圖Fig. 4 The HE staining results of testis sections of yellow catfish and hybrid yellow catfish “Huangyou No. 1”
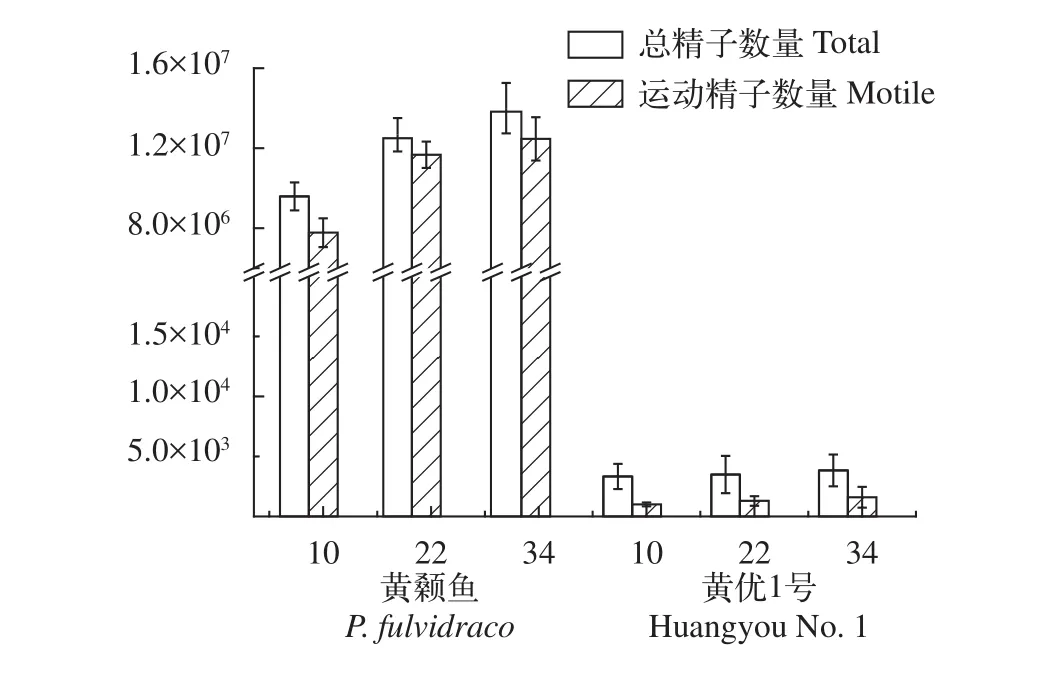
圖5 黃顙魚和“黃優1號”1 mL體積精液的精子數量(Total)和運動精子數量(Motile)Fig. 5 The total and motile sperm per mL in yellow catfish and hybrid yellow catfish
3 討論
據中國漁業統計年鑒統計結果顯示, 2017年黃顙魚養殖產量為4.8×108kg, 而瓦氏黃顙魚的需求量非常小而被忽略沒有統計。主要是由于消費者比較喜歡黃顙魚的體型和大小, 而瓦氏黃顙魚由于其體型、大小和體色等特征消費者的需求不足。華中農業大學黃顙魚種質資源與品種改良團隊采用雜交育種的方法, 選擇梁子湖水域采捕并經連續3代選育的黃顙魚為母本、長江河段中采捕并經連續2代選育的瓦氏黃顙魚為父本, 再經人工雜交獲得F1代即為雜交黃顙魚“黃優1號”。“黃優1號”從形態特征上比較接近黃顙魚, 在生長速度和上市規格整齊度等方面明顯優于黃顙魚, 具有推動黃顙魚產業的潛力。經過外觀形態比較, 我們發現通過胸鰭前緣和后緣的鋸齒狀凸起這一性狀可以簡便快速區分雜交黃顙魚和黃顙魚(圖 1)。
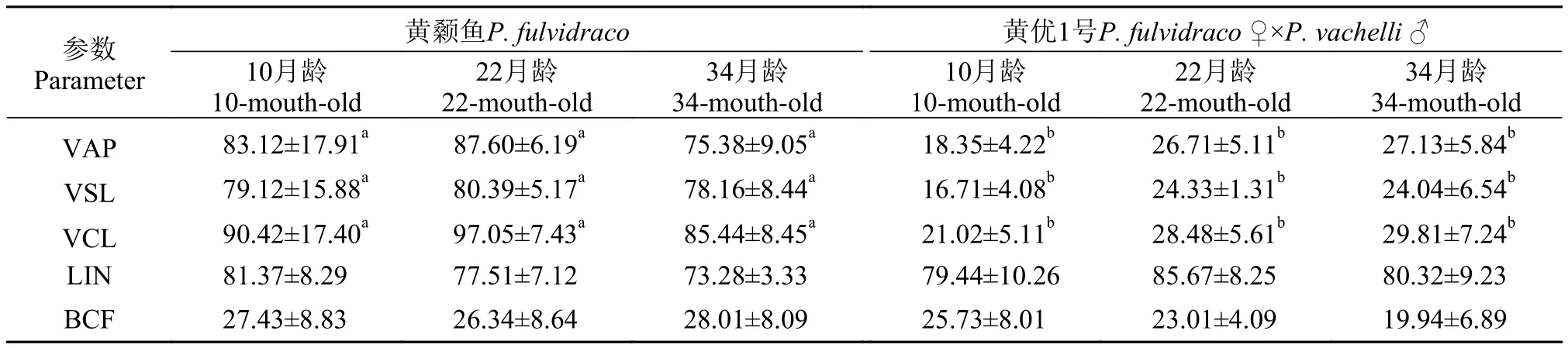
表1 黃顙魚與“黃優1號”精子運動參數的檢測Tab. 1 The parameters of sperm motility in yellow catfish and hybrid yellow catfish (μm/s)
黃顙魚和瓦氏黃顙魚同是黃顙魚屬但為不同種[19], 該2個物種之間的雜交為遠緣雜交。遠緣雜交能整合雙親的優點, 使后代在外形、生長速度、存活率以及抗病能力等方面表現出雜種優勢[19,20]。關于遠緣雜交后代的生殖能力已有一些研究, 發現一部分遠緣雜交后代具有生殖能力, 如紅鯽(♀)×湘江野鯉(♂)、錦鯉(♀)×紅鯽(♂)、團頭魴(♀)×翹嘴紅鲌(♂)[21]的正反交的遠緣雜交中能獲得可育的后代; 一部分遠緣雜交后代不具備生殖能力, 如blue drum (Nibea mitsukurii)和white croaker (Pennahia argentata)[22]、三角魴(♂)×草魚(♀)、團頭魴(♂)×草魚(♀)雜種均雜交是不育的[23]。關于雜交不育的生物學機制, 主要包括以下2種情況, 一種是雜交不孕(雜交不親和)現象, 即親本雜交不能產生后代; 另一種是雜種不育的現象, 即親本雜交后產生的子代不能生殖[20—25]。其中雜種不育性不僅僅與染色體的數目和形態及其基因有關, 還與異常的生理環境和生殖器官結構等有關。通常認為雜種不育性是雜種生殖細胞減數分裂過程中出現的異常同源染色體配對造成的。如牦牛與普通牛在部分常染色體和性染色體上的差異使雄性犏牛的同源染色體不能很好地配對形成精母細胞聯會復合體 (Synaptonemal complex, SC), 可能是導致不育的主要原因之一[25]。Blue drum (Nibea mitsukurii)和white croaker(Pennahia argentata)的雜交不育, 是由于雜種的原始生殖細胞發育因為有絲分裂缺陷發生了停滯而造成的[22]。
在魚類遠緣雜交研究中, 發現當母本染色體數大于或等于父本染色體數時, 可突破雜交F1生殖難關; 當母本染色體數目小于父本染色體數目時, 難以形成存活的后代[20—24]。當前, 魚類遠緣雜交的遺傳規律研究主要集中在染色體水平機制, 核-核、核-質之間的協調機制[20,21]以及RNA水平和蛋白水平的分子生物學方面的機制研究上[26,27]。雜交黃顙魚(黃顙魚♀×瓦氏黃顙魚♂)及其父母本的核型均為2n=52, 其雜交不育的相關機制有待進一步研究。

圖6 黃顙魚♀×黃優1號♂(HY)和黃顙魚♀×黃顙魚♂(HF)胚胎發育情況Fig. 6 The embryo development of yellow catfish♀דHuangyou No. 1”♂ (HY) and yellow catfish ♀×yellow catfish♂ (HF)
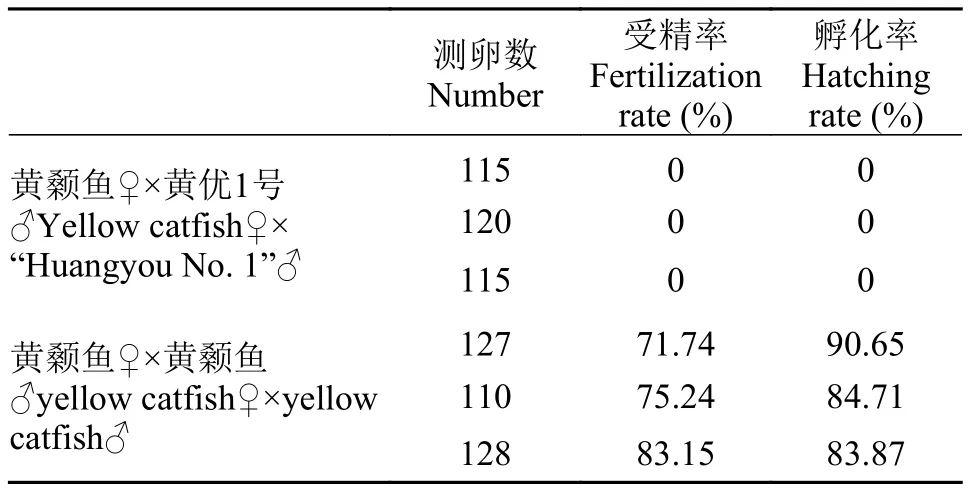
表2 黃顙魚♀ד黃優1號”♂和黃顙魚♀×黃顙魚♂的受精率和孵化率Tab. 2 Fertilization rate and hatching rate of yellow catfish♀דHuangyou No. 1”♂ and yellow catfish♀×yellow catfish♂
在動物世界中, 多種動物的雌雄個體的生長速度存在明顯的兩性生長差異, 也叫兩性生長異形(Sexual size dimorphism)[4], 人們發現許多魚類的雌雄個體的性成熟年齡和生長速度存在明顯差異。性成熟較早的個體, 由于在性成熟期間大部分營養物質轉移至性產物的生產, 身體的增長就減慢。例如半滑舌鰨(Cynoglossus semilaevis)和虹鱒(Oncorhynchus mykiss)等魚類, 它們的雌性個體性成熟時間要晚于雄性個體的性成熟時間, 從而致使雌性個體的生長速度顯著大于雄性個體[28]; 相反, 如果雄性個體性成熟晚, 其生長速度就會超過雌性個體,例如黃顙魚(Pelteobagrus fulvidraco)和羅非魚(Oreochromis niloticus)等[3,29,30]。本研究中, 雜交黃顙魚(黃顙魚♀×瓦氏黃顙魚♂)雌雄性腺發育均存在缺陷, 性成熟期間大部分營養物質轉移至身體生長,故雜交黃顙魚較黃顙魚有明顯的生長優勢。同時,雌雄性腺發育缺陷也導致了黃顙魚兩性生長異形的現象在雜交黃顙魚中被顯著減弱, 提高了商品魚的上市規格整齊度。雜交黃顙魚(黃顙魚♀×瓦氏黃顙魚♂)的性腺發育存在明顯缺陷, 卵巢為白色線狀組織呈現退化狀態且沒有卵子生成; 精小囊呈空腔狀結果, 精子總數及精子活力遠低于黃顙魚, 低于受精精液濃度的最低閾值[12]。根據從事水產一線工作者的經驗, 養殖4—5年齡的雜交黃顙魚雄性能產生可育的精子, 有待于進一步研究和證實。“黃優1號”在生長和生殖發育上的一些特征表明其可作為一個優良的黃顙魚養殖品種, 大幅度提高漁民經濟效益。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
