黑龍江鯉肌肉發育中miRNA對間隙連接蛋白基因(Cx43)的調控
2019-12-03 11:40:32艾顯輝趙金艷閆學春田麗紅
水產科學 2019年6期
艾顯輝,趙金艷,于 釗,閆學春,田麗紅,梁 洋
( 1.東北林業大學,黑龍江 哈爾濱 150040; 2.哈爾濱工業大學,黑龍江 哈爾濱 150001; 3.中國水產科學研究院 黑龍江水產研究所,黑龍江 哈爾濱 150040 )
在脊椎動物中,肌肉的生長除了受生肌決定因子家族成員(包括MyoD、Myogenin、Myogenin M 和MRF4等)以及一些負調控因子如肌肉生長抑制素(MSTN)的調控外[1],一些微小RNA(MicroRNA或miRNA)也通過對各自靶基因的作用協同參與肌肉的發生和分化過程[2-3]。富含在心肌和骨骼肌組織中的RNA被稱為肌相關微小RNAs(myomiRs),在龐大的myomiRs家族中,微小RNA-1(miR-1)和微小RNA-206(miR-206)具有共同的種子序列,屬于同一個基因家族,在骨骼肌中特異表達。研究顯示,在小鼠成肌細胞(C2C12)中過表達miR-1和miR-206,有明顯促分化作用;在肌肉疾病和功能紊亂模型試驗中,miR-1和miR-206也有明顯改變,說明二者與肌肉生長發育及肌肉相關疾病關系密切[4]。
miR-1和miR-206靶基因定位及其功能在小鼠等模式生物中研究較為深入而透徹,c-Met、HDAC4、pax7、pax3等均已確定是兩者共同的靶基因[5-6],另有研究顯示,在小鼠中間隙連接蛋白基因Cx43也是miR-1和miR-206的靶基因[7]。目前在小鼠基因組中發現20種間隙連接蛋白(Cx)基因,在人類基因組中存在21種間隙連接蛋白[8-9],其中Cx43基因是最重要的間隙連接蛋白基因之一。胚胎發育早期,Cx43基因在肌纖維中有廣泛表達,顯示能夠促進成肌細胞的分化、融合。體內試驗中,當肌肉受損后,肌衛星細胞通過誘導增殖分化來應對肌肉的損傷,此時在增殖的肌衛星細胞中Cx43基因的表達會迅速上調,并一直持續到肌細胞發生融合[10-11]。體外細胞試驗中也呈現這一趨勢,加入庚醇、辛醇等Cx43基因的抑制劑后,成肌細胞不能再正常分化,即使再加入生肌調節因子MRF4等也無法使肌細胞恢復分化[12],說明細胞間隙連接通訊對于體外肌細胞的分化融合是必不可少的。
魚類是重要的脊椎動物,地球上存在的魚類超過30 000種,約占脊椎動物總物種的50%,但魚類miRNA的研究相對滯后[13]。目前尚未見關于黑龍江鯉(Cyprinuscarpio)肌肉中miR-1和miR-206對Cx43基因調控的報道。因此,筆者以黑龍江鯉為試驗材料,自已經測序完成的鯉魚功能基因組數據庫中調取肌肉生長相關基因Cx43,通過熒光素酶報告系統,在細胞水平分析miR-1和miR-206對靶基因的調控,通過整體原位雜交等方法探明胚胎早期Cx43基因的表達情況,為探明鯉魚肌肉發育中miRNA的調控機理,優化黑龍江鯉品質,提高其生產性能奠定基礎。
1 材料與方法
1.1 試驗材料
試驗所用鯉魚精子、卵子及鯉魚成體均由中國水產科學院黑龍江水產研究所提供。NIH-3T3細胞實驗室自存。
組織獲取:新鮮黑龍江鯉處死,首先取出腦部組織,然后腹部解剖,取心、肝胰臟、腎、脾、鰓、腸、皮、紅肌和白肌等組織,迅速將取出的組織放入預凍的凍存管中(凍存管事先做好標記),凍存管放入液氮中冷凍,所有組織取出后置于-80 ℃冰箱保存備用。
1.2 試驗方法
1.2.1 RNA提取
分別取適量鯉魚胚胎,利用液氮充分研磨,加入Trizol使其充分裂解,10 000 r/min離心5 min,取上清液,分別以氯仿和異丙醇抽提、沉淀RNA,后以75%乙醇充分洗滌沉淀。用適量無RNA酶的去離子水溶解沉淀,變性凝膠電泳檢測RNA含量,測量總RNA量和純度,再將總RNA利用PolyA聚合酶加尾,利用接頭引物(Ambion)反轉錄成cDNA。
1.2.2 基因克隆及探針的制備
參照斑馬魚(Daniorerio)MyoD和Cx43基因的cDNA序列,設計簡并引物(表1),同時設計Cx43基因片段的突變引物,以鯉魚肌肉cDNA為模版,進行PCR擴增。將克隆結果送測序公司進行測序并按照說明書進行標記探針的制備。
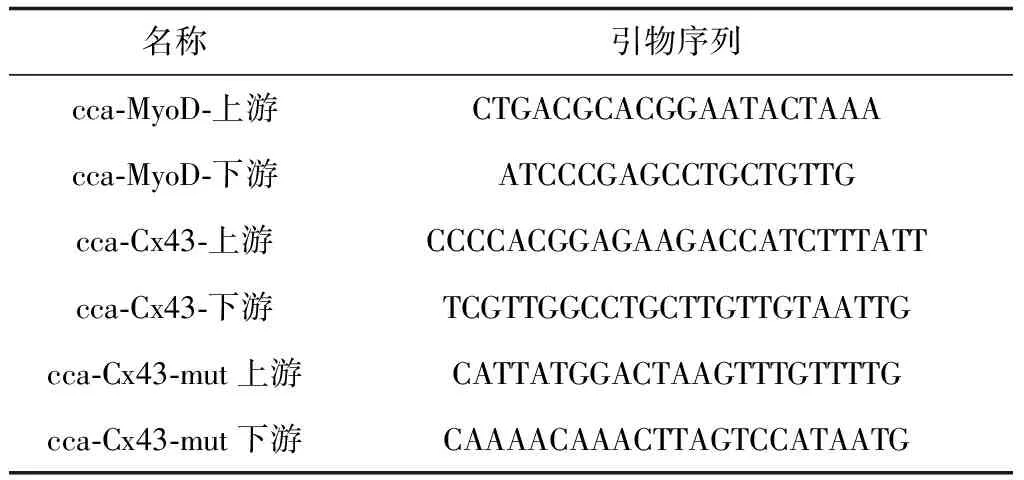
表1 引物序列
1.2.3 雙螢光素酶載體的構建與鑒定
根據表1所設計引物,PCR擴增得Cx43基因片段和Cx43-mut基因片段,膠回收后,利用hindⅢ和mluⅠ(紐英倫生物技術北京有限公司)進行雙酶切,同時雙酶切熒光素酶載體PMIR,將酶切后的目的片段進行檢測,正確后用T4 DNA連接酶,16 ℃過夜連接。連接檢測正確后進行轉化,并提取質粒進行雙酶切和測序鑒定。
1.2.4 細胞培養與轉染
1.2.4.1 細胞的復蘇與傳代培養
NIH-3T3細胞實驗室自存。自液氮中取出后置于37 ℃水浴中快速融化,轉移細胞懸液至15 mL離心管,加入5倍體積預熱到37 ℃的合成培養基,1000 r/min離心5 min,棄上清液,加入3 mL合成培養基,溫和吹打使其混勻,轉移到60 mm培養皿中,置于CO2培養箱中進行培養,培養條件:37 ℃,5%CO2、95%空氣。每日觀察細胞狀態。培養基配方:DMEM高糖培養基,添加10%胎牛血清(ES-FBS,BI公司,貨號04-002-1A),添加1%雙抗和1% GlutaMAX(100×)。
當復蘇培養細胞匯合度達到約80%時,棄去舊培養基,緩慢加入2 mL PBS,對細胞進行2遍清洗;棄去PBS,加入0.25%胰蛋白酶+0.04%EDTA的消化液500 μL,1~2 min后待大部分細胞變圓自底部脫落,再加入1.5 mL合成培養基終止消化;水平離心機離心(1000 r/min,5 min),棄上清液,按1∶3比例傳代培養。
1.2.4.2 細胞轉染
轉染前1 d將細胞接種于不含抗生素的培養基中,細胞轉染密度60%~70%匯合。按照Lipofectamine 2000說明書程序轉染細胞:
溶液A:每孔240 μL減血清培養基(Opti-MEM)+10 μL lipofectamine 2000,靜置5 min。
溶液B:每孔240 μL Opti-MEM培養基+2.5 μg質粒(總體積250 μL,與A液相同)。
溶液C:將A、B兩管緩慢混合,室溫下靜置20 min。
將溶液C逐滴均勻加入孔中,輕輕晃動培養板混勻,置37 ℃細胞培養箱中保溫4~6 h。之后更換為含有雙抗的培養基,37 ℃,5%的CO2中培養,48 h后檢測轉染水平。
1.2.4.3 雙熒光素酶檢測
細胞轉染48 h后棄去舊培養基,用預冷的1×PBS輕輕洗滌細胞2~3次;24孔板中加入1×ULB(通用裂解液)200 μL,水平搖床室溫搖15 min使細胞充分裂解;離心(10 000 r/min)3 min,取10 μL細胞裂解液于1.5 mL EP管,加50 μL螢火蟲熒光素酶底物,熒光光度儀測定螢火蟲熒光素酶的活性值F,再加50 μL海腎酶底物,檢測海腎熒光素酶發光值R,記錄F值、R值和比值。
1.2.5 RT-PCR
依照Takara PrimeScript產品的使用說明,用1 μg總RNA作為模板反轉錄,反轉錄引物為oligo-dT。PCR使用的基因特異性引物與Cx43探針合成引物相同。
1.2.6 鯉魚整體原位雜交
雜交第1 d:將受精后6、12、24 h和48 h的鯉魚胚胎分別依次放入裝有75%、50%和25%的1.5 mL無核糖核酸酶的EP管中復水,H2O2對鯉魚胚胎進行漂洗,蛋白酶K消化,4% PFA固定胚胎,加入含有探針的雜交液,70 ℃雜交箱中雜交過夜。
雜交第2 d:將雜交過夜的胚胎經溶液Ⅰ(50%甲酰胺,5×SSC pH=4.5,1% SDS)洗滌3次,溶液Ⅱ(0.5 mol NaCl,10 mmol Tris-Cl pH=7.5,1% Tween20)洗滌3次,溶液Ⅲ(50%甲酰胺pH=4.5,2×SSC)洗滌3次,每次洗滌均為30 min。洗滌后的胚胎利用馬血清封閉90 min,之后置于裝有地高辛抗體的TBST溶液中(1∶2000),放4 ℃冰箱中搖床洗滌16 h以上。
武戲自然就熱鬧些,武將們負旌旗持鋒刀,八字步一步高過一步,鏗鏘鏗鏘鏗鏘鏘。伴樂的那幾個老頭禿頭皺臉的,手底下忙著拉胡打鼓,頭也跟著不停地晃,恣意得很!
雜交第3 d:用TBST溶液(8 mg/mL NaCl,0.2 mg/mL KCl,25 mmol/L Tris-HCl pH=7.5,1% Tween20)對胚胎進行徹底洗滌,放于4 ℃冰箱中搖床洗滌過夜。
雜交第4 d:室溫下用NTMT(100 mmol NaCl,100 mmol Tris-HCl(pH 9.5),50 mmol MgCl2, 1% Tween20和2 mmol左旋咪唑)對胚胎進行搖床洗滌,3次/5 min。將胚胎移至裝有顯色液的培養皿中避光顯色(-4 ℃冰箱),顯色完成后用PBS溶液對胚胎進行洗滌以終止顯色,50%甘油透化30 min,觀察拍照。
2 結 果
2.1 鯉魚肌肉中miR-1和miR-206靶基因預測及分析
通過Targetscan軟件進行預測,結合前期閱讀資料,在斑馬魚中尋找到Cx43基因是miR-1和miR-206潛在的靶基因(圖1)。將鯉魚Cx43基因與斑馬魚序列進行比對,結果顯示,鯉魚Cx43基因的3′UTR區能夠和斑馬魚Cx43基因絕大部分匹配,且有6位堿基能與miR-1和miR-206種子序列完全匹配(圖2)。
2.2 PMIR-Cx43熒光素酶報告載體的構建
根據以上分析結果,在鯉魚Cx43基因3′-UTR區設計引物(表1),以提取的鯉魚肌肉cDNA為模板,進行PCR擴增。結果顯示,擴增條帶大約為500 bp,與理論值445 bp相符(圖3);將目的片段與克隆載體連接送去測序,測序結果與Cx43基因理論值匹配(通過DNAMAN比對)(圖4)。
2.3 PMIR-Cx43熒光素酶報告載體的鑒定
將上述含有Cx43基因3′-UTR區的克隆載體和熒光素酶載體PMIR通過酶切、連接、轉化獲取重組質粒DNA,再用hindⅢ和mluⅠ雙酶切鑒定。結果表明,酶切后得到的約500 bp的Cx43基因3′-UTR片段與預期大小相符(圖5)。

圖1 斑馬魚Cx43基因miRNA結合位點預測結果

圖2 斑馬魚和鯉魚Cx43基因序列比對結果
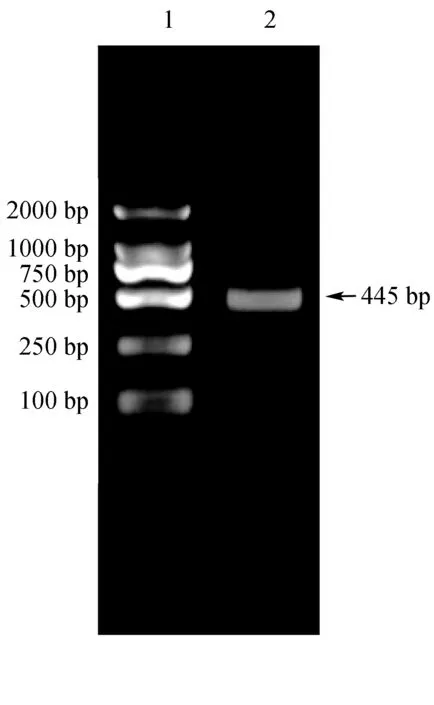
圖3 PCR結果1:DL2000 DNA Marker; 2:PCR產物.

圖4 Cx43基因3′-UTR測序結果
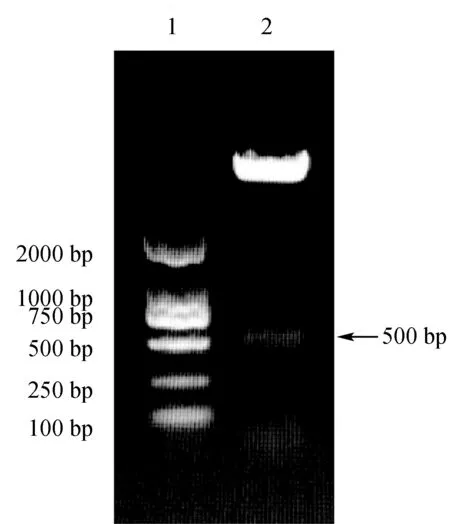
圖5 重組質粒 PCR檢測結果1:DL2000 DNA Marker; 2:PCR產物.
2.4 PMIR-Cx43-mut熒光素酶報告載體的構建
另外設計Cx43基因的突變引物(表1),擴增成功后連接克隆載體測序檢測,結果顯示突變體構建成功(圖6)。隨后構建熒光素酶報告載體PMI-Cx43-mut,分別用PCR和酶切檢測片段大小,與理論值約447 bp相符(圖7)。
2.5 miR-1和miR-206對靶基因Cx43調控作用分析
在3T3細胞中,通過檢測熒光素表達變化驗證miRNAs對靶基因表達的調控作用。利用脂質體將PMIR-Cx43載體、突變載體與合成的miRNAs,陰性對照及海腎熒光素酶載體共轉染3T3細胞,海腎熒光素酶的活性為內參,轉染后48 h檢測熒光素酶的活性。由圖8可知,與對照組相比,miR-1和miR-206均能下調pMIR-Cx43-3′UTR的熒光素酶活性,其中miR-206下調明顯。突變組miR-1熒光水平與對照組基本持平,miR-206組略低于對照組。
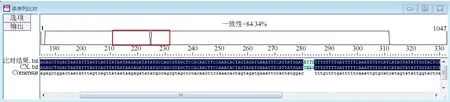
圖6 Cx43基因突變測序結果
圖7 PMI-Cx43-mut載體鑒定結果a.1,DL2000 DNA Marker; 2~7,PCR產物;8,陰性對照. b.1,DL2000 DNA Marker; 2,酶切產物.
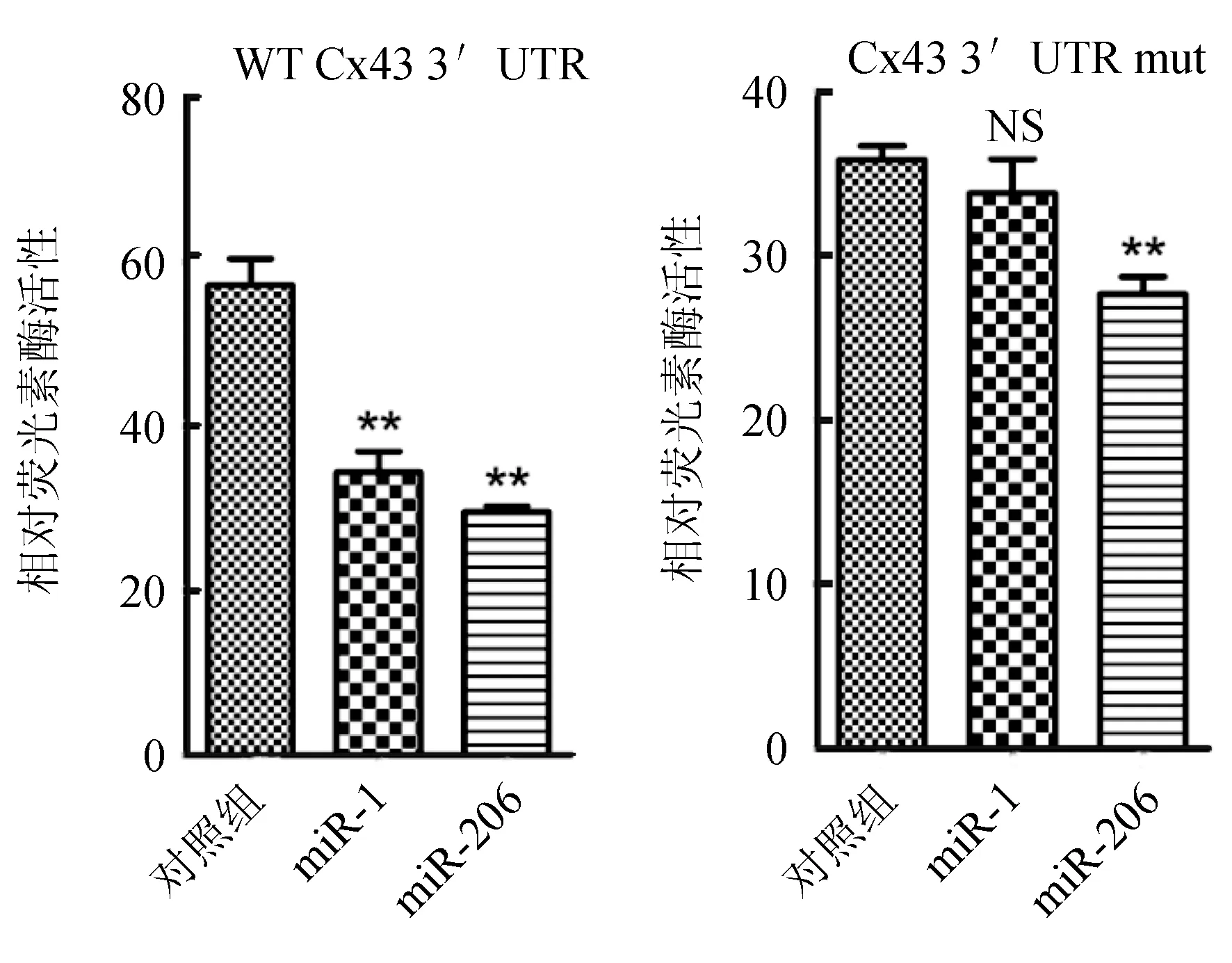
圖8 miR-1和miR-206對Cx43基因3′UTR的調控作用NS代表差異不顯著; **代表差異顯著(P<0.01).
2.6 Cx43基因在鯉魚胚胎不同時期的表達
對鯉魚胚胎發育6、12、24 h和48 h時期的Cx43表達情況進行分析,RT-PCR結果顯示,不同時期的鯉魚胚胎中Cx43的表達有明顯差異。6 h胚胎中已經開始表達但表達量較低,隨后表達量開始上調。當胚胎發育到24 h時,達到最高峰,此后表達呈現下降趨勢。MyoD基因作為陽性對照,Cx43基因表達趨勢與其有相似性(圖9)。
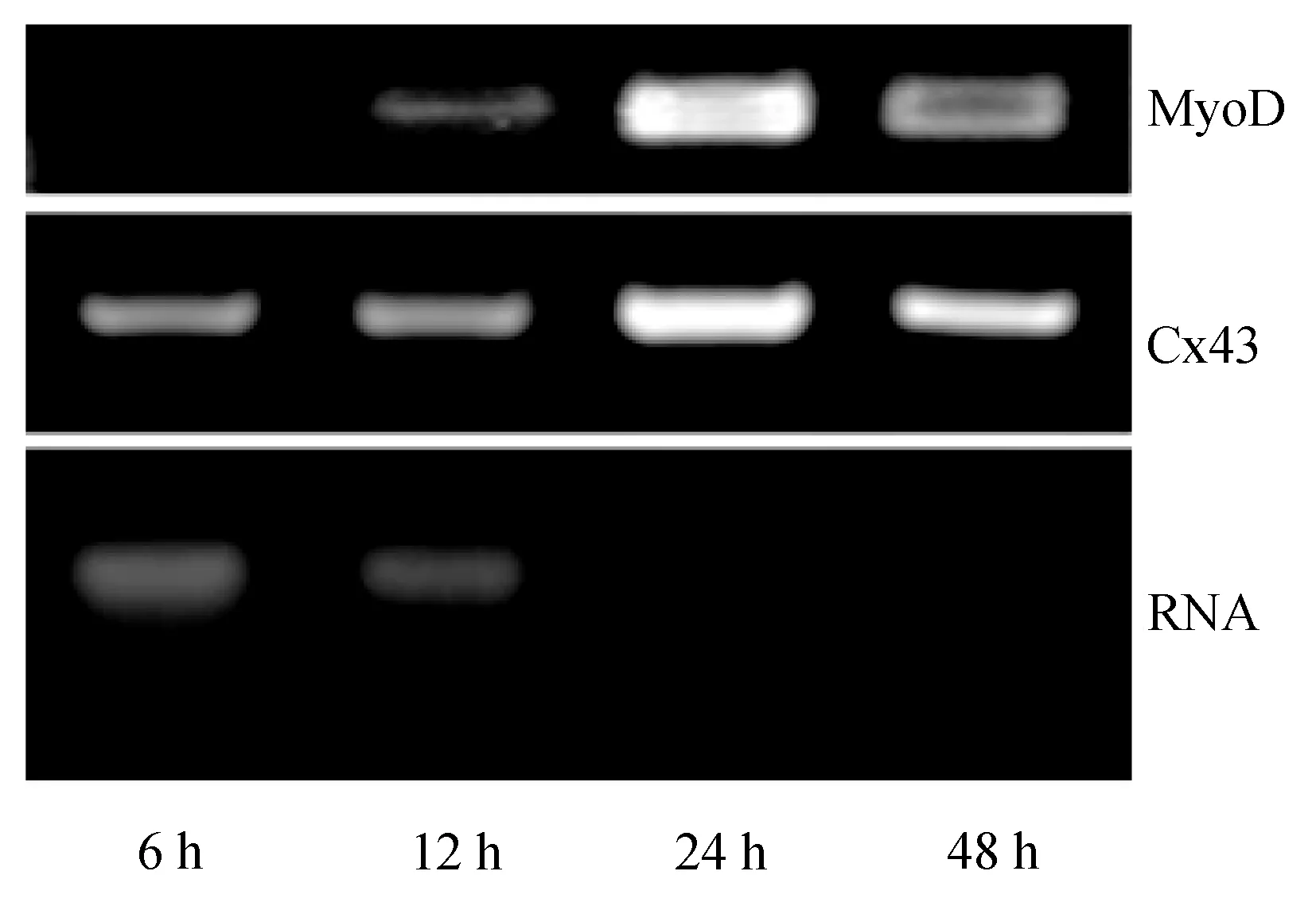
圖9 鯉魚胚胎發育不同時期RT-PCR結果
2.7 原位雜交分析Cx43基因在鯉魚胚胎中表達情況
3 討 論
在模式動物小鼠和斑馬魚中,發現多種與肌肉發育相關的miRNA,這些富含在心肌和骨骼肌組織中的miRNAs被稱為myomiRs,這些myomiRs通過對靶基因的調控而促進肌肉的發生、肌細胞的分化以及肌肉再生等過程[2-3]。其中miR-206和miR-1在骨骼肌中特異表達,且具有共同的種子序列,Cx43、c-Met、HDAC4、pax7、pax3等均是兩者共同的靶基因,但目前在黑龍江鯉肌肉中miR-206和miR-1對Cx43基因的調控尚未見報道。
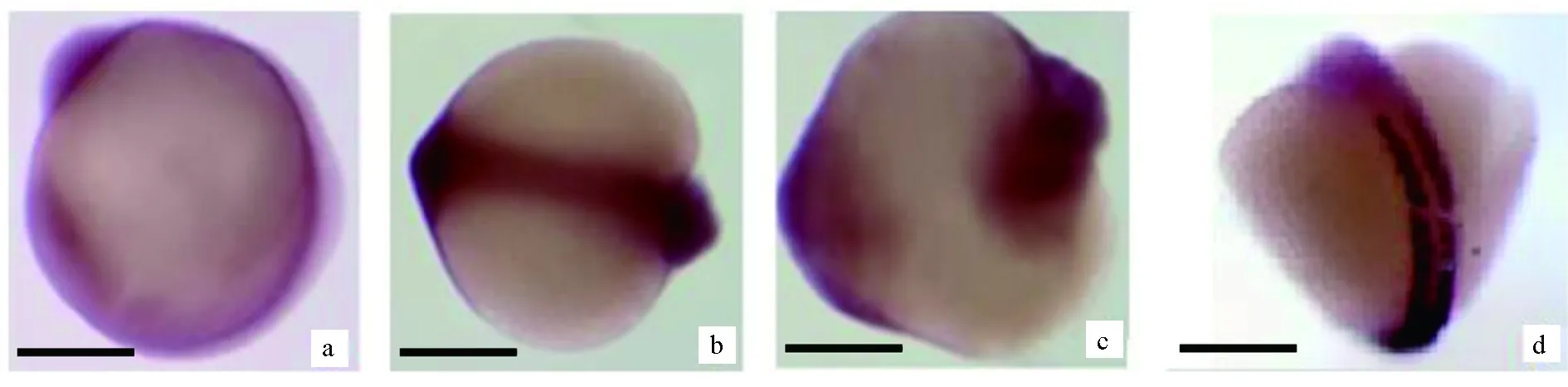
圖10 Cx43基因在12 h鯉魚胚胎中的表達情況a.側面觀察; b.背面觀察; c.腹側觀察; d.MyoD基因. 標尺為100 μm.
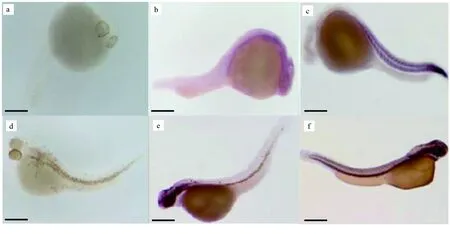
圖11 Cx43基因在24、48 h鯉魚胚胎中表達情況a.24 h陰性胚胎; b.24 h Cx43基因表達情況; c.24 h MyoD基因表達情況; d.48 h陰性胚胎; e.48 h Cx43基因表達情況; f.48 h MyoD基因表達情況. 標尺為100 μm.
3.1 miR-1和miR-206對Cx43基因的調控
研究顯示,Cx43作為一種重要的間隙連接蛋白,在小鼠等模式動物中受到miR-1和miR-206的調控,在胚胎發育早期的肌纖維中有廣泛表達,能夠促進成肌細胞的分化、融合[14]。Anderson等[15]利用小鼠C2C12細胞研究miR-206和miR-1對Cx43基因的調控作用時發現,在細胞融合過程中Cx43基因高表達,但分化后表達明顯下調,說明了myomiRs通過調控細胞間信息交流通道的關鍵蛋白Cx43蛋白的表達來促進細胞融合。李積永[16]通過研究大鼠骨骼肌運動性微損傷時發現,miR-206能通過調節Cx43基因的表達而促進大鼠腓腸肌的運動性損傷修復。另有研究顯示,在人血管平滑肌細胞中,miR-206與Cx43基因形成調控網絡來影響其增殖和發育[17]。
目前已經發現,在小鼠、大鼠、人、狗、牛和雞中Cx43的種子序列完全保守[18-19],本試驗前期利用生物信息學分析在斑馬魚中也發現了miR-206和miR-1結合位點,進一步分析鯉魚Cx43基因,發現鯉魚3′UTR區域與斑馬魚Cx43基因絕大部分匹配,且有6位堿基與miR-1和miR-206種子序列完全匹配,因此預測,在鯉魚中Cx43基因也能被二者調控。為進一步檢測該結果,克隆了鯉魚Cx43基因3′UTR區并構建熒光素酶報告載體與突變載體,利用脂質體轉染小鼠成纖維3T3細胞,結果表明,miR-1和miR-206均能下調含有Cx43 3′UTR序列載體的熒光素酶活性,其中miR-206作用明顯,miR-1對其抑制作用較弱。該結果與Anderson等[15]在小鼠中發現的作用機制相似,暗示在鯉魚中Cx43基因也是miR-206和miR-1的靶基因。
3.2 黑龍江鯉胚胎期Cx43基因表達模式
胚胎原位雜交方法可以分析mRNA在不同發育時期組織中的表達情況,是研究胚胎基因表達的常用技術,而地高辛所標記的探針具有較高靈敏度,有助于探討目的基因的調控機制[20-21]。本試驗利用胚胎整體原位雜交方法,MyoD基因作為陽性對照,研究鯉魚不同發育時期胚胎Cx43基因表達時發現,受精12 h后,Cx43基因在胚胎的眼周、頭部以及心臟部位表達,同時在近軸細胞和神經管一直延伸至胚胎的尾牙部分表達顯著;當胚胎發育至24 h時,Cx43基因主要集中在神經管、背部近軸細胞以及胚胎的體節中,且表達量最高,隨后呈現下降趨勢;至胚胎發育48 h在體軸和近軸細胞以及體節等處幾乎觀察不到基因表達。該結果與前期在鯉魚胚胎早期發育過程中MyoD基因的表達模式[22]類似,也有研究顯示,Cx43基因主要在小鼠早期胚胎中高表達,因此推測,在鯉魚中Cx43基因對成肌分化早期有重要調控作用。
另外,原位雜交結果顯示,Cx43基因始終在胚胎頭部、眼部高表達,由于該基因在腦、心臟等部位分布廣泛,尤其在神經元中,是神經膠質細胞等完成功能主要依賴的蛋白之一,因此幾乎在所有胚胎時期的腦中Cx43基因始終表達明顯[23]。
綜上,該研究在黑龍江鯉肌肉中探索了miRNA-206和miRNA-1和對Cx43的調控作用,并利用其胚胎進行Cx43基因表達差異性研究,為進一步探討鯉魚肌肉發育的調控機制,以及改良鯉魚肌肉品質提供了理論基礎。
