溫度對海月水母橫裂生殖的影響研究
2019-12-03 11:36:58付志璐王一兵
水產科學 2019年6期
付志璐,王一兵
( 1.廣西民族大學,海洋生物資源保護與利用重點實驗室,廣西 南寧 530008; 2.廣西高校微生物與植物資源利用重點實驗室,廣西 南寧 530008 )
海月水母(Aureliaaurita)是一種廣泛分布于世界海域的常見災害水母,主要分布于N 40°~S 70°的沿岸水域[1-3]。近年來這種水母在東亞水域大規模暴發,對漁業和沿岸發電廠造成嚴重災害[4-5]。海月水母主要以浮游動物、魚卵和仔魚為食,是魚類的直接捕食者和食物競爭者,其暴發對漁業資源可捕撈量形成威脅[6-7]。此外,海月水母的大量出現會對沿岸工業設施造成不良影響,因為海月水母經常堵塞核電站、火力發電廠等以海水作為冷卻水的冷卻水系統管道。因此,闡明海月水母暴發機制,探尋預防及緩解災害的方法已迫在眉睫。
海月水母的生活史包括底棲無性繁殖的螅狀體和浮游有性繁殖的碟狀體兩個階段。海月水母浮游生活的第一個階段稱為“碟狀幼體”階段,碟狀幼體通過螅狀體“橫裂生殖”也就是變態發育而產生。在這個過程中,螅狀體莖部變長,體表收縮出現裂節,隨后裂節逐漸加深,每一個裂節都變成一個帶有缺刻的小圓盤,最后每個小圓盤都發育成一個新的碟狀幼體。這個過程有助于成體水母種群的擴增,因為一只海月水母的螅狀體釋放出的碟狀幼體可以多達20~30只[8]。有研究表明,海月水母螅狀體橫裂生殖的引發通常歸因于幾個環境因子的變化,這些環境因子包括溫度、光照、食物[9-20]以及水體的生物化學成分[21](特別是水體中含碘甲狀腺素的含量)。在這些因素中,溫度對海月水母橫裂生殖的發生有著決定性的影響,因為橫裂生殖的主要季節與年度最低溫度時期相吻合[22-24],但不同地區的海月水母螅狀體對溫度的響應因地理位置的不同而有差異。如溫帶地區海月水母的橫裂生殖主要發生在季節性降溫之后[22-24];然而在亞熱帶地區的臺灣,季節性溫暖水溫(19~32 ℃)反而會加速海月水母的橫裂生殖[16]。雖然國內外已有諸多對海月水母橫裂生殖的研究,但目前對瀨戶內海海月水母橫裂生殖的研究尚未見報道。筆者在前人研究基礎上,通過室內試驗詳細觀察了來自日本瀨戶內海海月水母橫裂生殖過程中的形態變化,橫裂生殖開始時間、持續時間、裂節的數量以及每只螅狀體橫裂生殖所產生的碟狀體數量,以便檢驗溫度對誘導瀨戶內海海月水母橫裂生殖的作用,并對照已有研究結果進行比較分析,另外檢驗瀨戶內海地區的海月水母對溫度的響應是否會因為生態習性的差異而與其他地區有所不同,以期闡明海月水母暴發機制,探尋預防及緩解海月水母災害的方法。
1 材料與方法
2010年夏季在瀨戶內海捕捉成熟海月水母,從其口腕上收集浮浪幼蟲,將浮浪幼蟲放入直徑15 cm、深6.5 cm的透明塑料容器中,容器中裝有1 L鹽度為32的過濾海水(濾孔直徑0.2 μm),容器放置在25 ℃黑暗條件下,以使浮浪幼蟲附著在容器的底面和側壁。當浮浪幼蟲附著并變態發育成螅狀體后,每周投喂足量的鹵蟲(Artemia)無節幼體1~2次,投喂1 h后更換新鮮的過濾海水。
試驗開始前,將100~200個體型規格相似(柄徑長約 1 mm)、發育完全的16觸手螅狀體用解剖針輕輕從培養容器的底面和側壁移除,然后將每個螅狀體單獨放置在1個一次性滅菌培養皿中(直徑3.5 cm、深1 cm)。將放有螅狀體的小培養皿放入大的透明培養容器中(直徑15 cm、深6.5 cm),容器中裝有1 L鹽度為32的過濾海水。一周后,大部分螅狀體均附著在一次性滅菌培養皿的底面。將100只成功附著的螅狀體自25 ℃直接轉移至9、12、14、16、18 ℃溫度下,每個溫度下20只螅狀體。將各溫度下的20只螅狀體進行編號(將編號寫在每個螅狀體所附著的培養皿底面)。每個溫度設3個平行,每個平行6~7只螅狀體。螅狀體培養在0.5 L透明容器中,容器內裝有0.5 L過濾海水。試驗進行90 d,試驗期間螅狀體保持饑餓狀態,以消除食物的影響,同時每20 d更換1次過濾海水。所有螅狀體每日在解剖鏡下觀察1次,并詳細記錄各溫度下1~20號螅狀體形態變化、第1個裂節出現的時間、出現的裂節數量、第1只碟狀幼體的釋放時間、釋放的碟狀幼體數量以及最后1只碟狀幼體的釋放時間。
采用SPSS 13.0軟件對各處理組的均值進行單因素方差分析,以P<0.05為差異顯著,P<0.01為差異極顯著。當方差分析的結果為差異顯著時,采用Tukey′s檢驗進行兩兩比較,以確定各處理之間的差異。
2 結 果
2.1 橫裂生殖期間的形態變化
橫裂生殖期間螅狀體形態變化的一般特征如下:螅狀體的莖上首先出現1個裂節,在隨后的幾日內,裂節的數量逐漸增多。隨后螅狀體的觸手開始收縮,同時身體漸漸變成紅色。當觸手完全收縮后,緣瓣上的肌肉開始形成。螅狀體莖部的裂節加深,形成多層小圓盤,小圓盤的邊緣逐漸形成8個緣葉。當每個小圓盤發育成接近碟狀幼體的形態后,就開始搏動。然后發育好的碟狀幼體從橫裂體的后端一個接一個分離出來。橫裂體的基部會再次形成1個小而發育完全的螅狀體。
2.2 橫裂生殖開始時間、持續時間以及橫裂率
試驗期間內無任何螅狀體死亡。橫裂生殖開始時間(或稱為預橫裂時間)定義為自螅狀體轉移到試驗溫度下至其莖部出現第1個裂節的時間。由圖1可知,第1只螅狀體出現橫裂的時間為轉移到14 ℃下的第17 d。到第40 d,發生橫裂的螅狀體增加到8只。直至試驗結束,沒有螅狀體再出現橫裂現象。12 ℃下,第1只螅狀體發生橫裂現象出現在第31 d,隨后發生橫裂的螅狀體數量快速并顯著增加。到第42 d為止,共有13只螅狀體發生橫裂。在9 ℃下,第1只螅狀體出現橫裂的時間為第41 d,到第70 d,發生橫裂的螅狀體數量增至13只。在16 ℃下,第1只螅狀體發生橫裂的時間為第39 d,共有5只螅狀體出現橫裂現象。在18 ℃下,僅有1只螅狀體在第83 d發生了橫裂。在90 d的試驗期間內,所有的螅狀體僅發生過1次橫裂生殖,無螅狀體出現2次橫裂生殖現象。
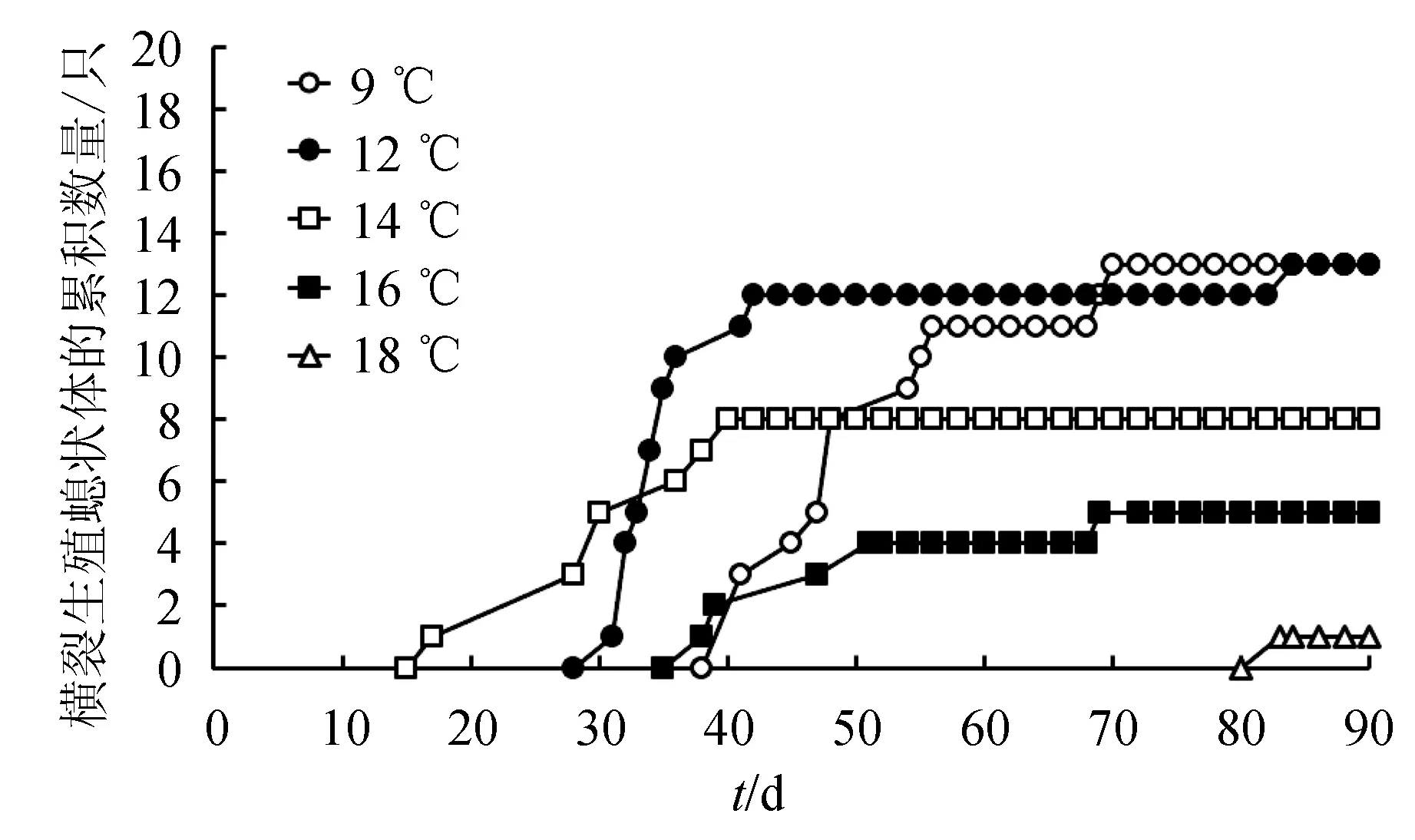
圖1 不同溫度下海月水母螅狀體橫裂生殖的累積數量
橫裂生殖開始所需要的平均時間在9、12、14、16、18 ℃下分別為(51.0±1.7)、(38.6±6.5)、(31.1±7.9)、(49.7±6.1)d和83 d(圖2)。由于16、18 ℃下的數據不足,只能對剩余溫度下的數據進行統計分析。統計結果顯示,不同溫度下海月水母螅狀體的橫裂生殖開始所需的平均時間差異顯著(單因素方差分析,P<0.05),14 ℃下橫裂生殖開始所需要的時間要明顯短于9 ℃下的時間(Tukey′s多重比較,P<0.05)。
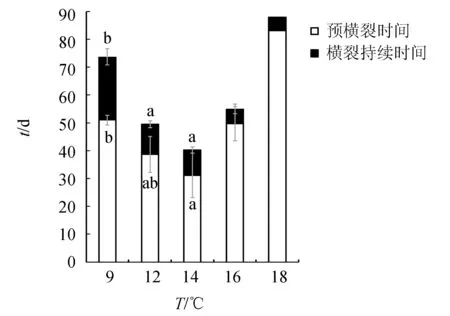
圖 2 不同溫度下海月水母的預橫裂時間和橫裂時間誤差線:標準差.圖中不同字母表示顯著差異(P<0.05).下同.
自螅狀體第1個裂節出現的時間到最后1個碟狀幼體釋放結束的時間被定義為橫裂生殖持續時間。平均橫裂持續時間在9、12、14、16、18 ℃下分別為(22.7±2.9)、(10.9±1.2)、(9.2±1.0)、(5.3±1.8)和5 d(圖2),各溫度間橫裂持續時間差異極顯著(單因素方差分析,P<0.01)。橫裂持續時間在12、14 ℃下顯著低于9 ℃(Tukey′s多重比較,P<0.05)。在9、12、14、16、18 ℃下分別有13、13、8、5、1只螅狀體出現了橫裂生殖現象,相當于各溫度下橫裂率分別為64.3%、65.1%、39.7%、26.2%和4.8%(圖3)。平均橫裂率在各溫度下差異顯著(P<0.05)。18 ℃下的橫裂率要顯著低于9 ℃和12 ℃(P<0.05)。
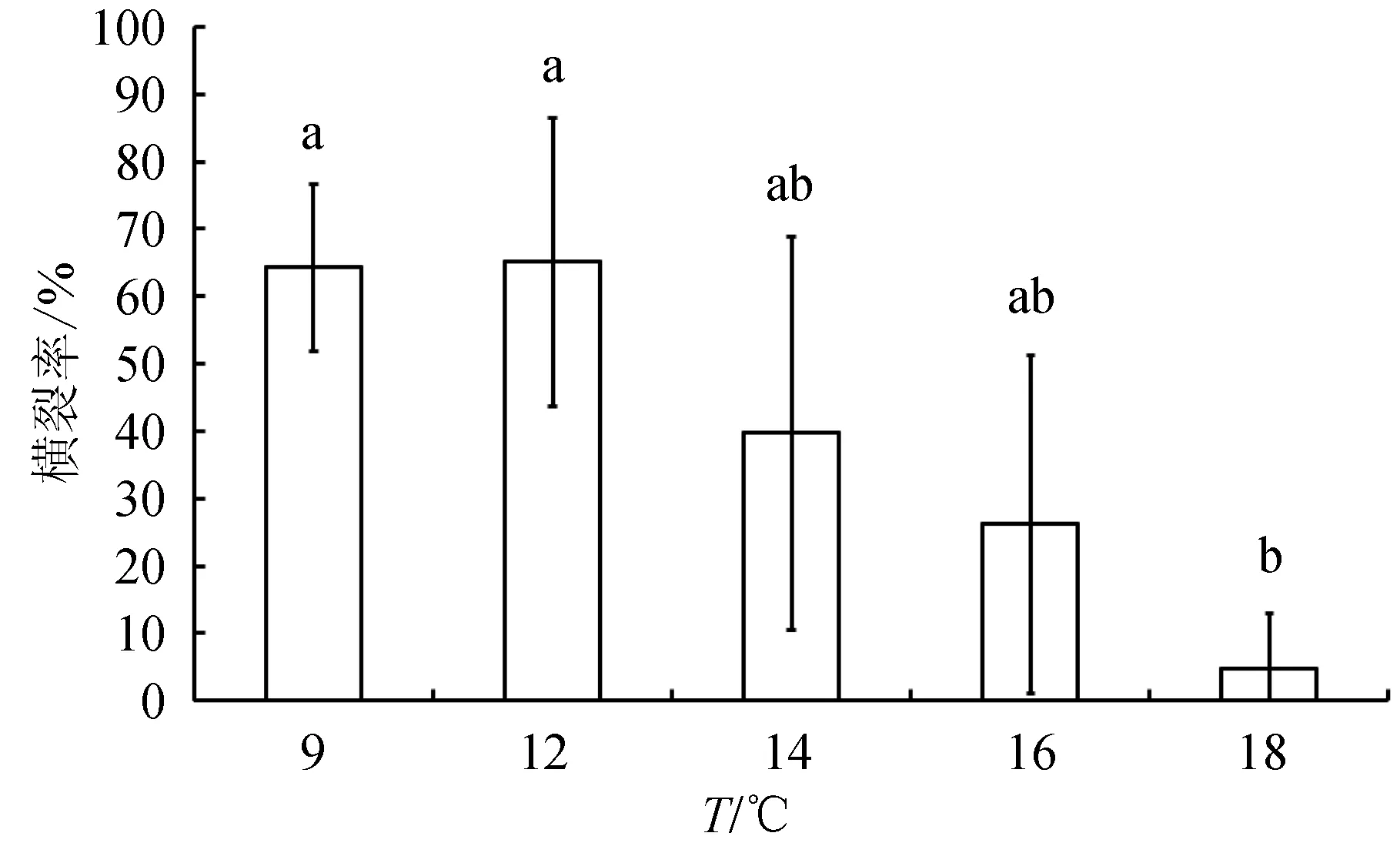
圖3 不同溫度下海月水母螅狀體的平均橫裂率
2.3 碟狀幼體的釋放
碟狀幼體釋放量隨時間的變化趨勢(圖4)與橫裂生殖發生模式(圖1)類似。9 ℃下,橫裂體所釋放的碟狀幼體數量最多,為36個。14 ℃下,橫裂體所釋放的碟狀幼體的數量與9 ℃相近,為34個。其余溫度下碟狀幼體的釋放數量依次為12 ℃>16 ℃>18 ℃(圖4)。9、12、14、16、18 ℃下,每只橫裂體釋放的碟狀幼體平均數量分別為(2.8±0.4)、(2.3±0.3)、(4.1±0.1)、(2.1±0.6)個和4個(圖5),水溫對釋放的碟狀幼體數量影響極顯著(P<0.01)。14 ℃下每只橫裂體所釋放的平均碟狀幼體數量極顯著高于其他溫度(P<0.01)。
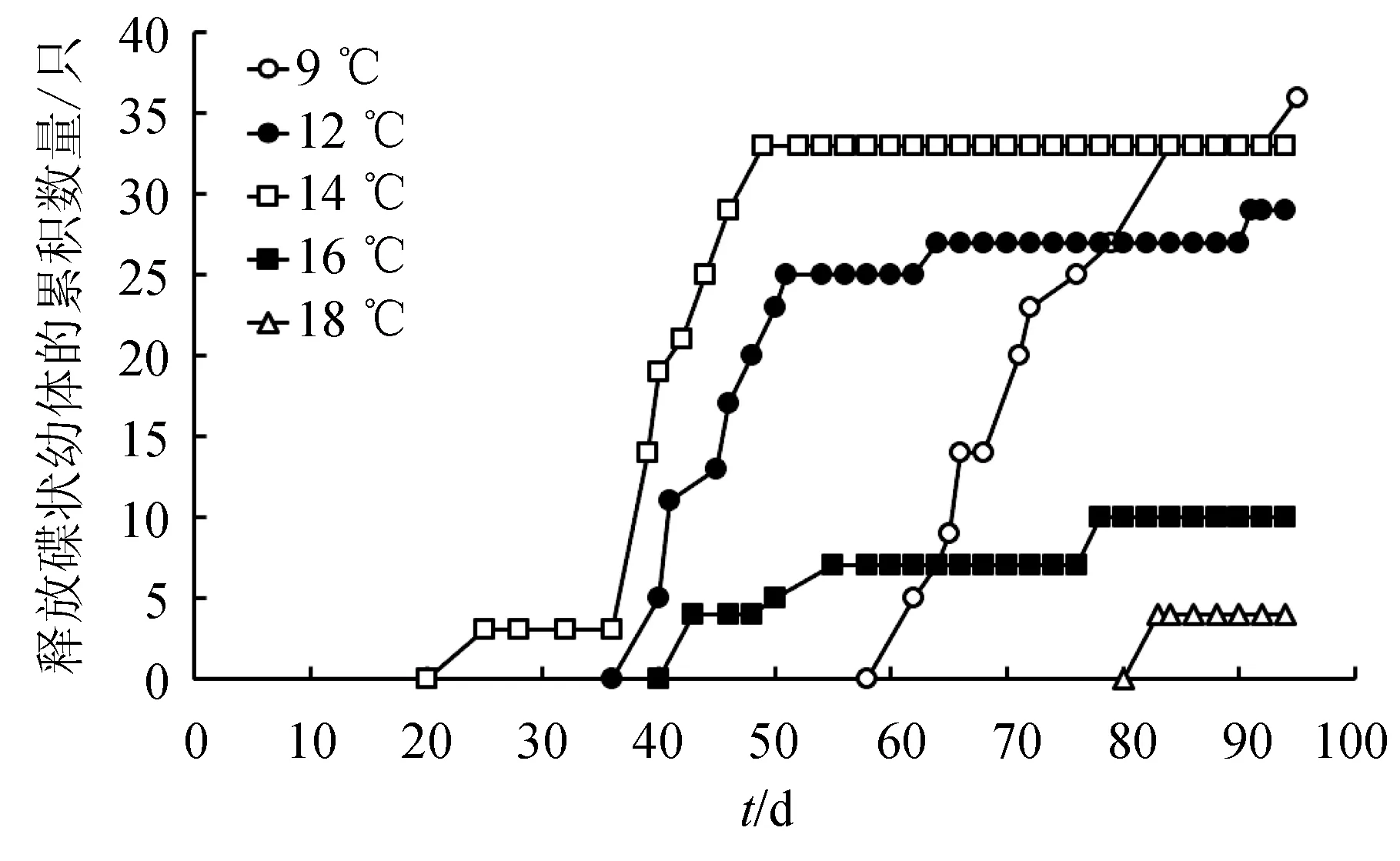
圖4 不同溫度下每月水母螅狀釋放的碟狀幼體的累積數量
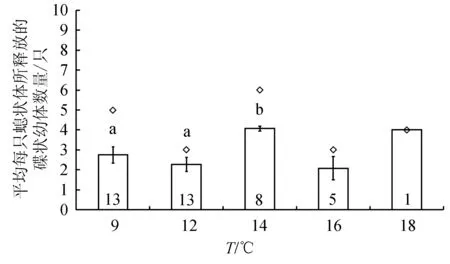
圖5 不同溫度下平均每只海月水母螅狀體所釋放的碟狀幼體數量柱內數字為用于計算的螅狀體數量,菱形為所釋放的碟狀幼體的最大數量.
3 討 論
與以往在整個群體水平上進行的大多數國內研究不同,本試驗對海月水母橫裂生殖的研究在個體水平上進行。每個水溫下的20只螅狀體均單獨附著在各自的培養皿中,且每個培養皿均有編號。這種方法優點是既可保證試驗數據的精確性,各螅狀體之間不會混淆,又可分清不同螅狀體所釋放的碟狀幼體。
3.1 水溫對海月水母橫裂生殖發育的影響
試驗證實,降溫可以誘導海月水母螅狀體發生橫裂生殖現象,這一結果與已有的研究結果基本一致。諸多研究表明,降溫能夠誘導海月水母發生橫裂生殖,Kroiher等[25]報道,降溫會導致法國北海地區的海月水母螅狀體發生橫裂生殖。Holst[26]表明,德國黑爾戈蘭島地區的海月水母螅狀體在10 ℃下的橫裂率要高于15 ℃下的螅狀體。Wang等[17]報道,中國膠州灣地區的海月水母在水溫降至17 ℃以下時會發生橫裂生殖。與其他溫帶地區相似,日本地區的海月水母螅狀體橫裂生殖的主要時間同樣出現在季節性降溫之后[22-24],但日本不同地區海月水母螅狀體發生橫裂生殖的水溫略有不同。據Kakinuma[27]報道,當溫度降至18 ℃以下時,日本北部奧陸灣的海月水母螅狀體會發生橫裂生殖。Han等[28]報道,日本西南部本莊地區的海月水母螅狀體在轉移至低于14 ℃的溫度后,停止攝食,隨之出現了橫裂生殖現象,但在18 ℃下螅狀體并未出現橫裂生殖。本研究所用海月水母螅狀體(來自日本瀨戶內海)在9 ℃和18 ℃之間出現了橫裂生殖現象。盡管與其他較低的溫度相比,18 ℃下發生橫裂生殖的速度相當緩慢,但在此溫度下螅狀體仍然出現了橫裂生殖現象,據此推斷,18 ℃可能是誘導瀨戶內海海月水母螅狀體發生橫裂生殖的溫度上限。
3.2 水溫對海月水母預橫裂時間的影響
研究證實,溫度會影響海月水母螅狀體橫裂生殖開始的時間。14 ℃下最先開始出現橫裂生殖現象,12 ℃或16 ℃下,橫裂生殖的開始時間延遲了1~3周。已有諸多研究報道過海月水母橫裂生殖在不同水溫下的開始時間,但不同螅狀體在相同或相似的水溫下橫裂生殖的反應卻并不相同。在臺灣南部,在20~30 ℃內,溫度越高海月水母螅狀體橫裂生殖現象出現越早[16]。中國膠州灣地區,在4~16 ℃內,海月水母螅狀體也呈現溫度越高橫裂生殖現象開始越早的趨勢[17-18]。海月水母的同屬種A.labiata也與此相似。太平洋東北部華盛頓地區的A.labiata在7~20 ℃內,同樣也是較高溫度時橫裂生殖現象出現更早[15]。與此相反,Kroiher等[23]報道,溫度降低幅度越大,橫裂生殖開始的時間越早。Han等[28]也報道,橫裂生殖在10 ℃下比14 ℃下出現的更早。本試驗結果顯示,中等寒冷的溫度下橫裂生殖開始得最早,這與一些研究所報道的在發生橫裂生殖的溫度范圍內,溫度越高橫裂生殖開始得越早,或降溫幅度越大橫裂生殖開始得越早不一致。此外,本試驗所使用的海月水母螅狀體與Han等[28]所用的螅狀體均來自日本,并且螅狀體來源地的地理距離并不遠,然而螅狀體對降溫的反應卻并不相同。本試驗結果與其他研究結果的不同,是由于螅狀體對各自棲息地環境條件的適應所造成的生理條件不同所致。
3.3 饑餓對海月水母釋放碟狀幼體數量的影響
為了消除食物引起的影響,螅狀體在轉移到試驗溫度后保持饑餓狀態,所以在90 d的試驗中,所有的螅狀體僅橫裂生殖1次,每只螅狀體所釋放的碟狀幼體的數量也較少,僅2~4個。有研究曾指出[17,29-30],在適宜的餌料豐度范圍內,餌料豐度越高,螅狀體所釋放的碟狀幼體數量就越多。此外,還有研究觀察到,海月水母螅狀體在攝食充分的食物后會發生多次橫裂生殖現象[12,31]。對A.labiata橫裂生殖的觀察也是同樣的發現[15],在每周充分投喂2次鹵蟲的條件下,多數A.labiata螅狀體在120 d的試驗中發生了2~3次橫裂生殖。因此本試驗中,如果螅狀體在橫裂生殖過程中攝食了充足的食物,它們可能會發生多次橫裂生殖以及釋放出更多的碟狀幼體。
3.4 暖冬對海月水母暴發的影響
盡管本試驗結果顯示9 ℃和12 ℃下海月水母的橫裂率幾乎相同,但是12 ℃下的橫裂時間僅為9 ℃的1/2。同時,12 ℃下橫裂生殖開始時間(預橫裂時間)比9 ℃下縮短了12.4 d。此外,已有的研究報道,海月水母螅狀體在較高的溫度下身體生長的速度更快[17,28-30]。因此,若食物充足,海月水母螅狀體在較高的水溫下會比在低溫條件下發生更多次橫裂生殖,從而釋放出更多的碟狀幼體。海月水母碟狀幼體在瀨戶內海的釋放時間通常為冬季及早春(水溫9~15 ℃),而此時水溫每升高1 ℃,海月水母橫裂生殖開始時間就會縮短4.1 d,同時橫裂持續時間會縮短3.9 d。因此,根據研究可以推斷,如果瀨戶內海冬季水溫偏高,則下一年度海月水母種群數量可能會大幅度增加。
對于海月水母螅狀體的所有試驗結果表明,12~14 ℃是最適合海月水母橫裂生殖的溫度,因為在此溫度范圍內橫裂生殖開始的最快,并且發生橫裂生殖的螅狀體比例更大。對其他溫帶地區海月水母進行的類似試驗結果與日本瀨戶內海海月水母有相同的趨勢:中等寒冷的溫度下有更高的橫裂率[25]。本研究的結果表明,暖冬可能會導致日本瀨戶內海海月水母在下一年度暴發。
4 結 論
試驗結果表明,溫度對誘導日本瀨戶內海海月水母的橫裂生殖有顯著影響。18 ℃是誘導螅狀體發生橫裂生殖的溫度上限。橫裂開始時間受溫度影響顯著,14 ℃時平均預橫裂時間最短,±2 ℃橫裂生殖開始時間會延遲1~3周。橫裂率同樣受溫度的影響,在9 ℃和12 ℃下最高(約65%)。橫裂持續時間隨溫度升高而下降。每個螅狀體釋放的碟狀幼體平均數量在各溫度下差異顯著,14 ℃時最高。綜上所述,12~14 ℃內橫裂生殖開始時間最短,橫裂生殖的螅狀體比例更大,是海月水母橫裂生殖的最佳溫度。由此可以推出,暖冬可能會導致日本瀨戶內海海月水母在下一年度暴發。
