水生動物抗菌肽的研究進展
2019-12-03 11:37:02胡豐曉吳苗苗張偉妮
水產科學 2019年6期
胡豐曉,吳苗苗,張偉妮
( 福建農林大學 海洋研究院,福建 福州 350002 )
受到外界病原入侵時,大多數生物的防御系統合成并分泌抵抗病原的重要功能物質——抗菌肽。抗菌肽是一類小分子多肽類物質,對病毒、細菌和寄生蟲等均有明顯的抵御作用。它通常由20~60個帶正電或負電的氨基酸按不同比例組成,分子量為2~10 ku。當前抗菌肽數據庫(http:∥aps.unmc.edu/AP/main.php)收錄的已知的編碼抗菌肽或抗菌肽的預測基因序列已達3000余種。截至目前,已從水生動物中分離出多種抗菌肽,如Tachyplesin I、Misgurin、Pleurocidin、Hepcidin和Paradaxins等。水生動物抗菌肽具有很高的抗毒素活性、較低的溶血活性和毒性小等優點,有著重要的應用價值[1]。
近年來,我國水產養殖業飛速發展,隨著高密度集約化養殖的普及,水環境污染嚴重、水產動物疾病層出不窮成為制約養殖業發展的主要瓶頸[2]。目前行業內主要采用抗生素和化學藥物防治水產養殖動物的疾病。抗生素和藥物的大量使用造成了養殖對象藥物殘留、病原菌耐藥性增加及環境污染等嚴重問題,而抗菌肽不易產生耐藥性,可替代抗生素的抑菌殺菌作用[3],還可改善動物腸道環境、提高生產性能,已成為國內外學者研究的熱點。筆者總結了抗菌肽的分類、作用機制,綜述了近年來水產動物養殖中抗菌肽的應用,以期為抗菌肽在水產養殖中的深入研究和產業化應用提供理論基礎。
1 抗菌肽的分類
抗菌肽廣泛存在于細菌、植物、昆蟲、水生動物、哺乳動物中,動物性抗菌肽的種類最多,也可人工合成。目前已知的抗菌肽中,根據氨基酸結構和功能將其分為殺菌肽、防御素、富含甘氨酸的抗菌肽、富含脯氨酸的抗菌肽等。
1.1 殺菌肽
殺菌肽最早見于天蠶中,又被稱為天蠶素[4]。目前研究證實,殺菌肽僅存在于節肢動物門的雙翅目和鱗翅目[5-6]。殺菌肽是一類含約35~39個氨基酸的短肽類物質,有較少的半胱氨酸,不能形成分子內二硫鍵,有強堿性的N端和強疏水性的末端酰胺化的C端。圓二色譜和二維核磁共振發現,其二級結構含有2個α-螺旋,兩螺旋間由丙氨酸—甘氨酸—脯氨酸組成“L”型的鉸鏈區連接,這種螺旋—卷曲—螺旋的結構特點對保持高抗菌活性具有特殊的重要性[7]。天蠶素在水產飼料中應用廣泛,發展前景廣闊[8-9]。
1.2 防御素
1966年,美國科學家Spitznagel等在哺乳動物的嗜中性粒細胞內發現一種強堿性的短肽類物質,隨后,由美國Lehere實驗室在兔巨噬細胞中分離純化得到,命名為防御素。防御素有3個或4個分子內二硫鍵,根據二硫鍵的位置及蛋白結構不同,分為α-防御素、β-防御素和θ-防御素,其中β-防御素的種類最多。防御素廣泛分布于哺乳動物和海洋生物中,其中魚類的β-防御素主要分布于皮膚黏膜和免疫器官中[10]。
1.3 富含甘氨酸的抗菌肽
富含甘氨酸的抗菌肽又稱蜂蜜素,其分子量約為8~30 ku,肽內含有約14%~22%的甘氨酸,具有較強的溶血性能。海洋生物含有大量富含甘氨酸的抗菌肽,其中來源于克氏原螯蝦(Procambarusclarkii)名為Procambarin的抗菌肽氨基酸數目高達155個,推測Procambarin可能參與克氏原螯蝦的宿主抗病毒防御系統,但其調節機制有待探索[11-12]。
1.4 富含脯氨酸的抗菌肽
富含脯氨酸的抗菌肽又稱蛙皮素,是一類由動物免疫系統分泌的線性多肽,最初由Zasloff等[13]在非洲爪蟾(Xenopuslaevis)中分離獲得。肽內脯氨酸含量最多,精氨酸含量次之[14-15]。
2 抗菌肽的作用機理
抗菌肽的作用機理與其分子結構特點密切相關,但是目前尚未研究透徹。綜合目前研究,被公認的抗菌肽抑殺菌機理主要有兩種:膜裂解機制和非膜裂解機制。
2.1 膜裂解機制
抗菌肽能夠辨別真核生物和原核生物細胞膜組成的差異,與細菌細胞膜結合并產生不可逆孔隙或使膜不穩定的肽[16-17]。抗菌肽與細胞膜的作用模式有地毯模型、桶板模型、環孔模型和聚集體模型(圖1)[18]。
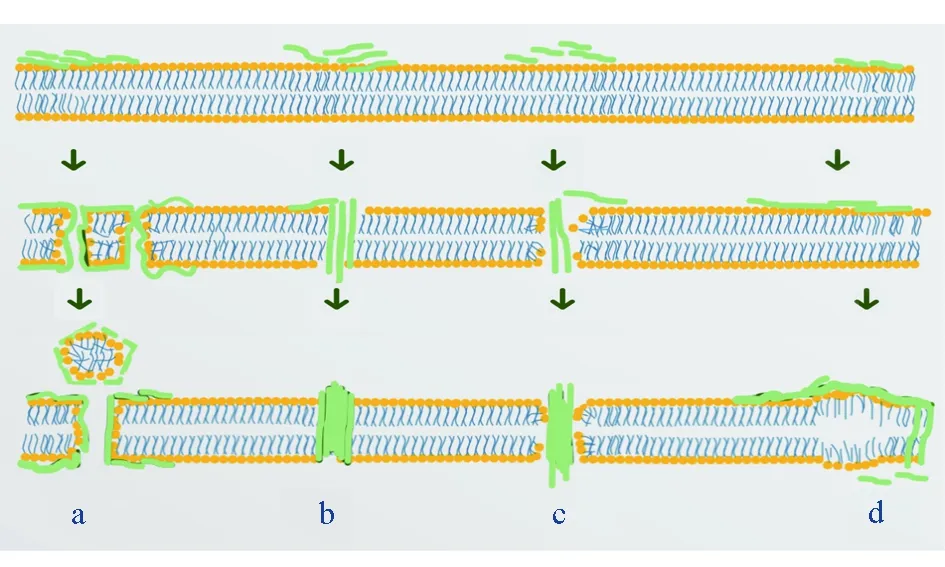
圖1 抗菌肽與細胞膜作用模型a.地毯模型; b.桶板模型; c.環孔模型; d.聚集體模型.
抗菌肽的正電區與細菌細胞膜的磷脂雙分子層通過靜電作用,由原來無規則的線狀結構變為有規則的二級結構——α-螺旋,增大與細胞膜接觸的表面積,改變了細胞表面的離子通透性,影響電勢、離子動態平衡和酸堿平衡及酶活性,抑制呼吸,并造成胞內物質流出,最終導致細菌細胞死亡[17,19]。
不同種類抗菌肽中殺菌肽、防御素和富含甘氨酸的抗菌肽的作用機制大致相同,機體在受到病原體侵害時,這些抗菌肽容易與革蘭氏陰性菌外膜的脂多糖或者革蘭氏陽性菌的肽聚糖相互作用,破壞其細胞膜,使細胞質外泄,從而殺死細菌[20-21]。
2.2 非膜裂解機制
部分抗菌肽不能通過破壞細胞膜結構起到殺死細胞的效果,而是與細胞膜上的特異性受體結合,進入到胞內干擾細胞的正常代謝[22]。這種非膜裂解機制主要有以下作用模式:(1)干擾與阻斷DNA和RNA的復制與合成;(2)抑制細胞蛋白質的合成[23];(3)抑制細胞分裂,促進細胞凋亡[24];(4)抑制細胞內酶的生物活性,起到抑菌和殺菌的功能[25-26]。
以這種作用機制類型為主導的是富含脯氨酸的抗菌肽,這類抗菌肽通過在細胞內結合Dank蛋白以抑制細菌的生物學功能,使細菌死亡[27]。
3 水生動物抗菌肽的結構與生物學活性
隨著水生動物抗菌肽研究的推進,來源于水生無脊椎動物(水母、貝、蝦、蟹等)和脊椎動物[大黃魚(Pseudosciaenacrocea)、石斑魚(Epinephelus)等]的抗菌肽新種類不斷被發現。基于不同的來源和結構特征,這些抗菌肽的活性特征也不盡相同,大多都具有不同程度的抗革蘭氏陽性菌和陰性菌活性,有的還具有抗真菌、抗寄生蟲和抗腫瘤活性等(表1)。
3.1 水生無脊椎動物抗菌肽
3.1.1 腔腸動物
腔腸動物最初被稱為刺細胞動物,種類可達1萬余種,分布于多種水域。有學者從腔腸動物體內提取出了多種抗菌肽,例如Aurelin,一種來自海月水母含40個氨基酸、分子質量約為為4.3 ku的免疫活性蛋白,具有適度的抗革蘭氏陽性和陰性細菌活性。
3.1.2 軟體動物
目前關于軟體動物抗菌肽的報道也較多,研究來源主要有牡蠣、扇貝等常見種類。這些軟體動物的免疫機制主要依賴非特異性免疫系統,血淋巴和組織中的各種抗菌肽是重要的組成部分。貝類中已發現了多種抗菌肽,例如牡蠣的β-thymosin和亞洲海蛤的Perinerin,它們是一類含有40~50個氨基酸的抗菌肽,具有抗革蘭氏陽性和革蘭氏陰性細菌活性。
3.1.3 甲殼動物
截至目前已發現多種來源于甲殼動物血細胞的抗菌肽物質,如Crustin、Sphistin、Arasin 1等。這類抗菌肽多數含約40個氨基酸,且肽中附帶二硫鍵,大部分對革蘭氏陽性和革蘭氏陰性細菌有抗性,少數也具有抗真菌活性。來源于擬穴青蟹的抗菌肽Sphistin,對革蘭氏陽性菌和陰性菌具有抗性,通過吸附發揮其抗微生物活性,最終破壞細菌細胞膜,而對哺乳動物或蟹正常細胞沒有表現出毒性[49]。
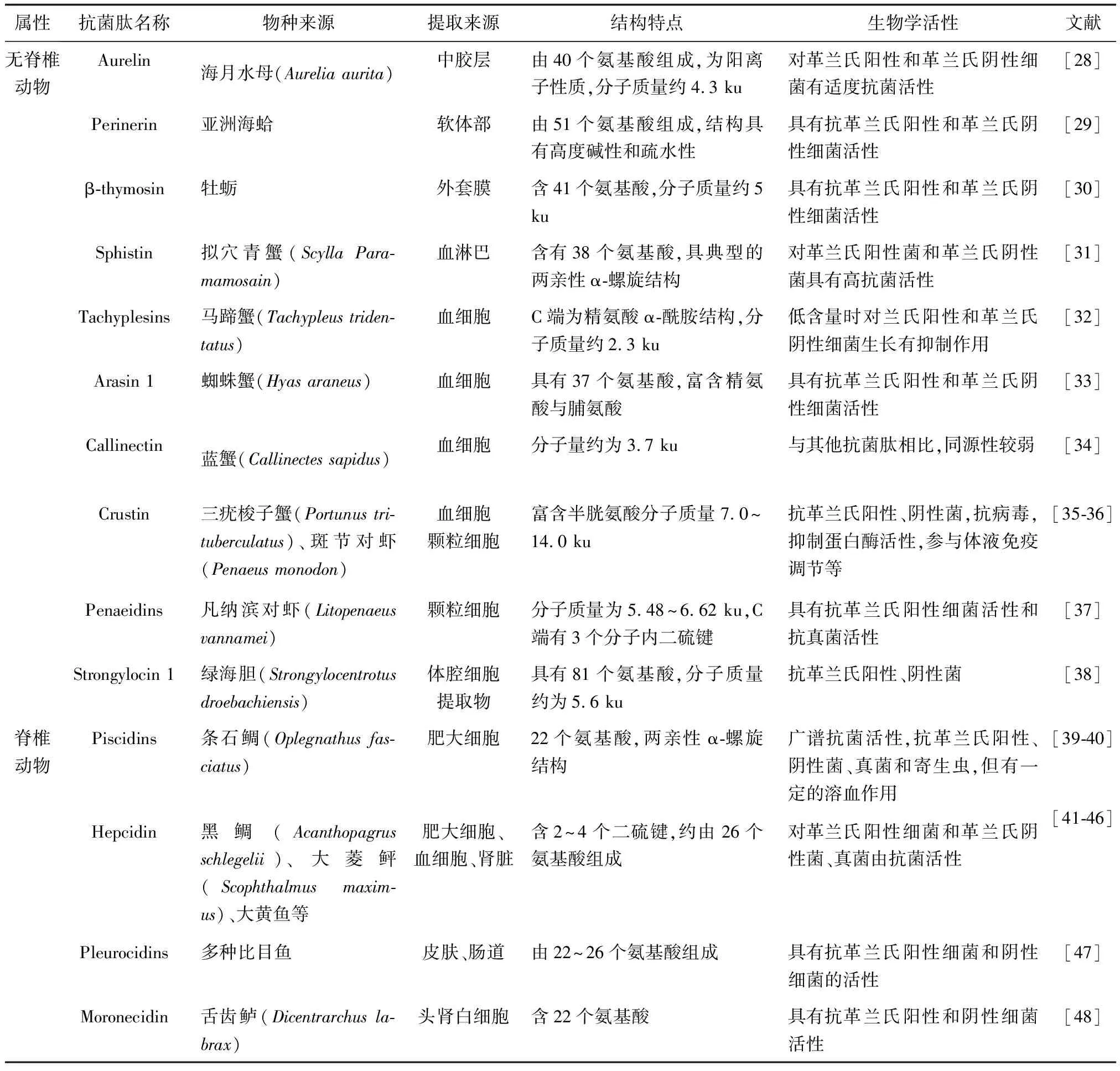
表1 水生動物常見抗菌肽的結構與活性特征
3.1.4 棘皮動物
對棘皮動物中抗菌肽的研究,關于海膽的較多。例如來自綠海膽的Strongylocin 1與Strongylocin 2是一類分別有89和90個氨基酸組成的抗菌肽,具有抗革蘭氏陽性、陰性菌活性。
3.2 水生脊椎動物抗菌肽
目前,水生脊椎動物抗菌肽方面的研究關于魚類的最為豐富。例如Piscidins、Hepcidin、Pleurocidins等,這些抗菌肽大多來源于魚類皮膚黏液、腸道、血細胞等,具有約24個氨基酸,部分含有α-螺旋結構或二硫鍵等結構,都具有抗革蘭氏陽性、陰性菌活性,少數還可抵抗真菌和寄生蟲等。
近期研究發現,某些魚類的機體含有多種類型功能相似或不同的抗菌肽。Peng等[50]報道了尼羅羅非魚(Oreochromisniloticus)5種Piscidin類抗菌肽(TP1~5),其中TP3和TP4具有強大的抗菌活性。溶血性試驗表明,TP4可誘導人紅細胞溶血,兩種抗菌肽都顯示出對革蘭氏陽性菌和陰性菌的抗菌活性,TP3對腫瘤細胞的選擇性高于正常細胞,TP3和TP4都具有抗革蘭氏陽性和陰性菌活性。Mu等[51]將大黃魚的lchampl、lchamp2-1/4、lchamp2-2等5種成熟肽純化合成,發現合成肽對革蘭氏陽性和革蘭氏陽性有抗菌活性。從不同魚類體內獲得的同種類型的抗菌肽也可能具有不同的活性特征。Shin等[52]通過比較發現,從南極魚(Nototheniacoriiceps)分離出的兩種抗菌肽moronecidin樣肽,顯示出比2002年首次從雜交條紋鱸(Moronechrysops×M.saxatilis)中分離出來的抗菌肽moronecidin具有更窄的活性光譜和更高的耐鹽性。
4 抗菌肽的應用
4.1 提高水產動物的生長性能
抗菌肽可以調理水產動物腸道性能,清除腸道內有害物質,保證胃腸黏膜結構和功能的完整性,提高動物對飼料的消化吸收而促生長[53]。陳奕彬等[54]提出,水產動物食用抗菌肽一段時間后,不僅能提高水產動物生長性能與水產品質量,還可解決抗生素在體內蓄積的問題。適宜蛋白水平下的抗菌肽濃度可降低赤點石斑魚(E.akaara)組織中部分氧化酶活力,增強腸道消化能力,促進石斑魚的生長[55]。鄭宗林等[56]發現,飼料中添加0.3~0.5 g/kg的抗菌肽顯著促進了紅姑魚(Sciaenopsocellatus)的生長。在飼料中添加適量Surfactin,可改善中華鱉(Pelodiscussinensis)稚鱉血液生化指標、提高生長性能[57];在飼料中添加50、100 mg/kg和150 mg/kg的抗菌肽后,促進羅非魚生長性能的效果與高劑量的黃霉素相似[58]。由此可見,抗菌肽在作為飼料添加劑方向有很大的發展潛力。綜上所述,抗菌肽可作為飼料添加劑促進水產動物生長,提高質量增加率和特定生長率,降低飼料系數。但是,水產動物種類多樣,抗菌肽對水產動物應用的最適劑量也不盡相同,抗菌肽對水產動物的促生長機制及合理使用亟待深入研究。
4.2 抑制水體病原菌活性
現階段水產行業迅速發展,水體中普遍存在的病原菌嚴重影響行業的生產效益。研究發現,水生生物分泌以及人工合成的抗菌肽可有效抑制病原菌的活性。學者從大口黑鱸(Micropterussalmonides)的皮膚、脾臟、肝臟、卵、黏液中提取抗菌肽,對這些抗菌肽進行抑菌性檢測,對比檢測結果發現,經肝臟與皮膚提取出的抗菌肽對大腸桿菌(Escherichiacoli)和嗜水氣單胞菌(Aeromonashydrophila)有抑菌活性,而其他3種提取物則無抑菌效果[59-60]。劉權偉等[61]發現,人工合成鲇魚(Silurusasotus)黏液的抗菌肽可嚴重破壞腐敗希瓦氏菌(Shewanellaputrefacines)和大腸桿菌的細胞結構,對這兩種菌體活性產生有效的抑制。郭子好等[62]將來源鋸緣青蟹(S.serrata)的抗菌肽Scygonadin成功轉化到畢赤酵母(Pichia)中,并進行抑菌試驗,結果發現,Scygonadin的表達產物在長達62 h內可有效抑制溶壁微球菌(Micrococcuslysodeikticus)的生長,相比較而言,此產物對嗜水氣單胞菌的抑菌能力較低。將石斑魚β-防御素的酵母表達載體轉化到畢赤酵母中,結果顯示,重組抗菌肽可有效抑制大腸桿菌和嗜水氣單胞菌的繁殖[63]。人工合成擬穴青蟹抗菌肽Sphistin對嗜水氣單胞菌、熒光假單胞菌(Pseudomonasfluorescens)和施氏假單胞菌(P.stutzeri)有極高的抑菌活性,并可破壞大腸桿菌和金黃色葡萄球菌(Staphylococcusaureus)的細胞膜[31]。以上研究表明,抗菌肽可有效抑制水域中多種常見病原菌的活性,如大腸桿菌、嗜水氣單胞菌和金黃色葡萄球菌等,從而有效提高養殖動物的成活率。
4.3 增強水產動物的免疫功能
面對著養殖水體中種類繁多的病原微生物,如細菌、病毒和寄生蟲等,水產動物的非特異性免疫機制非常重要,是機體基礎的防御機制。諸多研究表明,在飼料中添加抗菌肽能夠有效提高水生動物的免疫功能。例如在飼料中添加適量抗菌肽S200養殖一段時間后,對凡納濱對蝦進行攻毒試驗結果表明,抗菌肽可提高凡納濱對蝦的成活率、溶菌酶活性以及超氧化物歧化酶活性,以此提高機體免疫功能[64]。據彭開松等[65]報道,將雞的抗菌肽雞β-防御素13按40 mg/kg的劑量添加到尼羅羅非魚飼料中,能提高尼羅羅非魚抗維氏氣單胞菌(A.veronii)BJCP-9感染的能力。用抗菌肽Chelonianin處理后,羅非魚的死亡率大大降低,抗病力顯著增強[66]。斜帶石斑魚(E.coioides)經100 mg/kg的Surfactin處理后,肝臟抗氧化能力有所提高[67]。花鰻鱺(Anguillamarmorata)僵苗在飼養過程中因生理代謝功能低下產生機體抗氧化能力降低,繼而引起飼養成本增高,對僵苗施以適量的Surfactin后,魚體腸道的抗氧化能力顯著提高[68]。在中華絨螯蟹(Eriocheirsinensis)的飼料中添加0.4%的抗菌肽,飼喂兩個月后,中華絨螯蟹的血清溶菌酶活性和一氧化氮含量與對照組相比有增加的趨勢(P>0.05),顯著提高肝臟、肌肉組織超氧化物歧化酶和總抗氧化水平(P<0.05)[69]。綜上所述,添加適量的抗菌肽可有效地提高水產動物免疫相關酶活性和抗氧化能力,進而增強水產動物抗病力。
5 展 望
抗菌肽為一種內源性防御成分,具有廣譜性、無殘留、不損害正常細胞的優點,在水產養殖業和生物健康免疫領域可替代抗生素,顯著提高水產動物的生長性能和免疫功能。但是抗菌肽在成為未來的“綠色藥物”道路上仍面臨諸多問題,生物體內的抗菌肽含量極微,制成藥物時,程序復雜、成本較高,難以大規模生產。在今后的研究中要探明以下幾點:(1)抗菌肽種類繁多,結構多樣,具體的抗菌抑菌機制還未完全清楚,結構與功能的相關性還需進一步闡明;(2)目前抗菌肽利用的主要方式還是作為飼料添加劑,但是抗菌肽易被蛋白酶水解,如何高效利用抗菌肽仍需進一步探討;(3)目前抗菌肽主要以發酵生產為主,成本高昂,研究抗菌肽的高效提取及人工合成方法非常必要。未來,隨著技術手段的不斷創新,新發現的天然抗菌肽及人工合成的抗菌肽種類會越來越多,經濟、高效、廣譜的抗菌肽無疑具有巨大的應用潛力和廣闊的發展前景。
