樂山市金口河區板廠坪蜜環菌的分離及系統發育分析
2019-12-05 02:45:02陳克保林宜錦歐夢瑩
現代農業科技 2019年20期
陳克保 林宜錦 歐夢瑩

摘要 ? ?蜜環菌作為天麻重要共生菌,能為天麻的生長提供營養。了解區域性蜜環菌的多樣性有助于選育優良的土著蜜環菌,進而提高區域性天麻的產量和質量。本文應用純培養技術和基于ITS基因序列的系統發育分析,對金口河區板廠坪天麻種植區分離的21份蜜環菌進行了多樣性研究。結果表明,21株蜜環菌劃分為6個形態類群;系統發育分析顯示分離菌株與Armillaria同源性最近,相似性都在99% 以上,主要分布在Armillaria gallica、Armillaria sinapina和Armillaria mellea 3個種內。
關鍵詞 ? ?蜜環菌;分離;系統發育;四川樂山;金口河區
中圖分類號 ? ?Q949.32 ? ? ? ?文獻標識碼 ? ?A
文章編號 ? 1007-5739(2019)20-0049-03 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 開放科學(資源服務)標識碼(OSID)
Abstract ? ?As an important symbiotic bacterium of Gastrodia elata,Armillaria can provide nutrition for the growth of Gastrodia elata. Understanding the diversity of regional honey fungus will help to select excellent Armillaria,and thus increase the yield and quality of Gastrodia elata. In this paper,the pure culture technique and phylogenetic analysis based on ITS gene sequence were used to study the diversity of 21 Armillaria isolated from the Gastrodia elata planting area of Jinkouhe district Banchangping.The results showed that 21 strains of Armillaria were divided into 6 morphological groups. The phylogenetic analysis showed that the isolates had the closest homology with Armillaria,and the similarity was over 99%,mainly distributed in Armillaria gallica,Armillaria sinapina and Armillaria mellea.
Key words ? Armillaria;isolation;phylogeny;Leshan Sichuan;Jinkouhe District
蜜環菌(Armillaria)隸屬于真菌門擔子菌亞門層菌綱傘菌目口蘑科蜜環菌屬的一種藥食兩用真菌,在世界各地均有分布[1]。目前全世界已經鑒定的蜜環菌生物種有40多種,其中歐洲7個、北美洲10個、非洲5個、澳洲5個,亞洲至少19個,我國分布有15種[2]。該屬真菌有些種可與天麻和豬苓共生為天麻和豬苓的生長提供營養,天麻共生蜜環菌生物種有Armillaria gallica、Lineage 1、CBS O、CBS L、Armillaria nabsnona、Nag. E 和Armillaria cepistipes,而豬苓共生蜜環菌的種類尚未明確[3]。蜜環菌屬有些種還是嚴重的森林病原菌[4],能侵染針葉樹、闊葉樹等600多種樹木引起根腐病,造成經濟損失[5]。此外,有些種則具有重要的食藥用價值。東北名菜“小雞燉蘑菇”中的蘑菇別稱榛蘑就是蜜環菌中的一種[6]。蜜環菌還具有催眠、鎮靜、改善心腦、血液循環、降血糖、抗氧化、調節免疫和抑制腫瘤等藥用價值[7]。
在過去的幾十年里,基于形態特征(子實體大小、形狀、孢子顏色等)和交配試驗對蜜環菌物種的鑒定取得了很大的進展。然而,由于蜜環菌生物種類眾多且形態特征相似,這些方法都非常有限[8]。近年來,分子生物學技術在蜜環菌生物種的鑒定方面得到了很好的應用[9]。在分子生物學技術中,采用單個的ITS、IGS、EF-1α序列鑒定方法,不能準確鑒定到每一個蜜環菌種[6],而多序列結合RFLP 等技術的應用,使難以區分的近緣種或未知種的準確鑒定成為可能[1]。
金口河板廠坪天麻種植區地處四川盆地西南邊緣,該區分布有大量的杉木、柳杉、核桃等喬木樹種以及珙桐、水杉、銀杏等珍貴樹種。同時板廠坪天麻種植區海拔1 900 m,年均氣溫17 ℃,年日照時數1 130 h,年均降水量1 100 mm,平均濕度在80%左右,非常適合蜜環菌的生長[10],而此地區蜜環菌的研究還未見相關報道。本研究嘗試從金口河板廠坪天麻種植區采集樣本,分離蜜環菌,應用可培養技術和基于ITS基因序列的系統發育分析該地區蜜環菌的多樣性,以期為開發金口河蜜環菌資源及探索適合天麻生長的本地蜜環菌選育提供參考。
1 ? ?材料與方法
1.1 ? ?試驗材料
1.1.1 ? ?菌株。蜜環菌菌材采自樂山市金口河區板廠坪天麻種植區,將采回后的菌材培養在外尺寸530 mm×370 mm×270 mm、內尺寸485 mm×330 mm×220 mm、裝有2/3土壤、濕度70%~80%的周轉箱中,同時立刻進行分離蜜環菌。
1.1.2 ? ?分離純化培養基。①分離培養基:玉米麥麩培養基(玉米粉100 g,麥麩80 g,大豆10 g,白糖18 g,水1 000 mL);純化培養基:馬鈴薯葡萄糖瓊脂培養基(新鮮土豆200 g,切成小塊,加入蒸餾水1 000 mL,水煮沸時開始計時,30 min后,停止加熱,用紗布過濾,補充濾液至1 000 mL,濾液中加入2%葡萄糖,濾液加熱后,加入1.5~2.0%瓊脂,培養基制作完成,并常規倒平板或斜面,待用)。
1.1.3 ? ?主要試劑和儀器。玉米粉、麥麩、大豆粉、白糖和新鮮土豆,購于超市;Biospin真菌基因組DNA提取試劑盒,購于杭州博日科技有限公司;PCR試劑,購于生工生物工程(上海)股份有限公司;瓊脂糖為BIOWEST AGAROSE;PCR擴增儀為EasyCycler;電泳儀為DYY-8C型,北京六一儀器廠;凝膠成像儀為BioDocAnalyze;本研究使用的真菌通用引物ITS4和ITS5由生工生物工程(上海)股份有限公司合成。
1.2 ? ?試驗方法
1.2.1 ? ?菌索分離、純化。試驗采用黃萬兵等[11]和張玉方[12]相結合的分離方法對蜜環菌進行分離,即在超凈工作臺中用無菌水將蜜環菌菌索洗凈,用75%酒精浸泡30 s,用無菌濾紙擦干菌索,用左右手的拇指和食指分別捏住菌索相距1 cm處,在酒精燈火焰旁緩慢用力向兩邊拉至斷裂,斷裂處會拉出長短不一的白色菌絲,用無菌剪刀剪取3~5 mm白色菌絲,接種于玉米麥麩培養基,置于26 ℃恒溫培養箱中避光培養。培養6 d后將分離培養基中的菌索按以上操作接種于PDA培養基內,并在相同條件下培養,觀察記錄菌索形態特征。
1.2.2 ? ?DNA提取。蜜環菌菌索用無菌水沖洗,用無菌濾紙吸干菌索表面水分,-80 ℃冷凍備用;DNA提取采用Biospin真菌基因組DNA提取試劑盒,提取步驟按說明書嚴格操作。
2 ? ?結果與分析
2.1 ? ?分離菌株的形態特征
從板廠坪天麻種植基地分離得到21株蜜環菌,采用黃萬兵[15]對蜜環菌形態類群劃分方法,將分離的21株蜜環菌劃分為6個形態類群。G1類群菌株菌索開始生長時間相對較早,菌索直徑較細,分支不明顯,生長不彎曲;G4類群菌株菌索開始生長相對較晚,菌索直徑較粗,分支明顯,生長卷曲(表1)。
2.2 ? ?菌株DNA提取及系統發育分析
各類群菌株在培養6 d后菌索達到DNA提取用量,試驗選取6個代表菌株進行DNA提取,真菌通用引物ITS4和ITS5擴增的DNA分子大小在850 bp左右,擴增結果和凝膠電泳檢測結果一致(圖1),PCR擴增得到了較高質量的總DNA,擴增片段經1%瓊脂糖凝膠電泳可以檢測到清晰明亮的PCR擴增條帶。
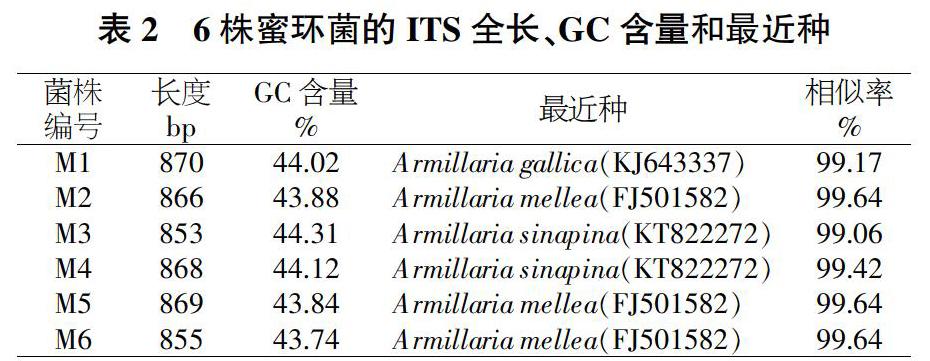
根據測序結果,經SeqMan軟件拼接序列后,6個菌株的ITS序列長度范圍在853~870 bp之間,GC含量在43.84%~44.31%之間。將6個菌株的ITS序列在GenBank公共數據庫中用Blast程序進行序列比對,分離菌株與Armillaria同源性最近,相似性比對都在99%以上。M1和Armillaria gallica的相似性為99.17%,M2、M5和Armillaria mellea的相似性均為99.64%,M3、M4和Armillaria sinapina的相似性為99.06%,M6和Armillaria mellea的相似性為99.30%(表2)。
本文對蜜環菌的ITS序列進行測定和系統發育分析,并構建了系統進化樹(圖2)。系統發育分析和序列比對結果一致,M1菌株和Armillaria gallica聚在一支,支持率為100%;M2、M5、M6親緣關系較近,以87%的支持率占據一支,同時和Armillaria mellea以73%的支持率聚在一大支。
3 ? ?結論與討論
本文基于ITS序列對金口河區板廠坪天麻種植區蜜環菌進行了分子鑒定,共鑒定出3種蜜環菌物種:Armillaria gallica、Armillaria sinapina和Armillaria mellea。與貴州天麻主產區和鄂西地區的蜜環菌生物種相比較,金口河區板廠坪天麻種植區蜜環菌物種多樣性相對較低[6,11],造成這種現象的原因可能是單個的ITS、IGS、EF-1α序列的鑒定方法不能準確對此地區部分蜜環菌進行鑒定[6]。門金鑫等[1]研究表明,用于蜜環菌生物種的鑒定還有限制性片段長度多態性技術(RFLP,restriction fragment length polymorphism)和隨機擴增多態性 DNA 技術(RAPD,random amplified polym-orphic DNA)等,但這些技術也只能準確鑒定部分蜜環菌生物種。多序列結合RFLP 等技術更能較好地對金口河區板廠坪天麻種植區蜜環菌資源進行鑒定。
在本文鑒定出的蜜環菌生物種中,3種蜜環菌在歐洲、北美洲以及亞洲均有分布,卻在澳洲和非洲未見分布[16-17]。通過比較發現造成這種差異性的原因可能是蜜環菌對不斷變化的地理環境的適應,從而進化出不同的物種。此外,根據Coetzee MPA等[18]對中國蜜環菌的研究,這3種蜜環菌在中國均有分布,然而在不同省份[6,11]蜜環菌的分布情況也不同,說明不同蜜環菌對環境因子的適應性存在著差異。另一方面,對于氣候和環境變化如何影響蜜環菌以及寄主和它們的共存卻知之甚少,過去幾年對病原體方面的研究并沒有帶來許多新見解。例如,預測氣候變化對蜜環菌根腐病發病率的影響是具有挑戰性的,因為生命周期的不同階段,包括傳播、感染和繁殖,可能會直接或間接的受到影響,并且影響可能完全不同。缺乏與蜜環菌相關樹木死亡率的長期研究及其與人類活動引起的環境變化的多重復雜相互作用,限制了對不斷變化的環境中蜜環菌的進化理解。
