微生物發酵產阿魏酸酯酶及釋放阿魏酸研究概述
2019-12-26 06:14:38孫曉明辛嘉英王艷吳永存王寧陳書明
食品研究與開發 2019年1期
孫曉明,辛嘉英,,*,王艷,吳永存,王寧,陳書明
(1.哈爾濱商業大學省高校食品科學與工程重點實驗室,黑龍江哈爾濱150076;2.山西農業大學動物科技學院,山西太谷030801)
阿魏酸(ferulic acid,FA)[1],其化學名稱為 4-羥基-3-甲氧基肉桂酸,是普遍存在于植物細胞壁中的一種酚酸,有兩種異構體:順式和反式,順式阿魏酸為黃色油狀物,一般指的反式阿魏酸為微黃色或白色斜方晶體,相對分子量194.29,熔點為174℃。反式阿魏酸水溶性差,穩定溶于乙醇,難溶于冷水、石油醚、苯,可溶于熱水、丙酮、甲醇。自然界中[2],阿魏酸以酯鍵和醚鍵分別在木質素與木質素之間、半纖維素與半纖維素之間、木質素與半纖維素之間相連接從而在植物細胞壁間形成致密的網狀骨架結構,從空間上阻礙了纖維素酶和半纖維素酶的降解作用,使得整個細胞壁變得堅硬。自然條件下的阿魏酸有水溶態、脂溶態和束縛態。水溶態阿魏酸存在于植物的細胞質中,該狀態下的阿魏酸與一些小分子(單糖、二糖、多胺等)結合呈易溶態;脂溶態是指阿魏酸與一些脂溶性物質(甾醇等)結合,主要存在與植物表面的蠟質層中;束縛態指阿魏酸以酯或醚的形式與植物細胞壁物質(多糖、蛋白質和木質素)結合[3]。
阿魏酸是國際公認的天然安全抗氧化劑,也是近年來認知度極高的防癌物質。具有降血脂、防治冠心病、抗血栓、抗氧化、抗菌消炎、抗突變和防癌等生理功能,被廣泛應用于保健品、食品添加劑、化妝品和醫藥等領域[4-5]。制備阿魏酸的主要方法有:從植物中直接提取法、化學合成法和生物合成法。化學合成法和生物合成法提取成本高,能耗大,不利于節約環保,所以采用酶法發酵從植物細胞壁中提取阿魏酸的綠色清潔生產方法由于其對環境友好、專一性強、反應條件溫和,已成為廣泛研究的熱點[6-8]。
真菌、細菌和酵母都能分泌的阿魏酸酯酶(ferulic acid esterase,FAE)[9]是指能將阿魏酸甲酯、低聚糖阿魏酸酯和多糖阿魏酸酯中阿魏酸游離出來的一種酶。屬于羥基酸酯酶的一個亞綱,能打斷阿魏酸在谷物細胞壁中以單體和多種二聚體的形式為主與阿拉伯木聚糖和阿拉伯糖殘基相連接的酯鍵[10-12],破壞細胞壁骨架結構,從而釋放阿魏酸。
本文主要對近年來國內外用不同產酶微生物來發酵產阿魏酸酯酶以及酶法發酵產阿魏酸的研究進展進行了綜述,為研發阿魏酸新工藝提供研究依據。
1 產阿魏酸酯酶研究進展
1.1 阿魏酸酯酶的培養條件及微生物來源
在植物細胞壁中,阿魏酸主要以酯鍵連接到存在于細胞壁的阿拉伯木聚糖的阿拉伯糖殘基上,還有一部分以阿魏酸二聚體的形式連接到阿拉伯木聚糖聚合物上,阿魏酸酯酶可以打斷阿魏酸與細胞壁多糖的連接,高效降解多糖并獲得低聚糖及阿魏酸[10-12]。自1987年MACKENZIE CR等[13]培養橄欖色鏈霉菌(Streptomyces olivochromogenes)時發現了阿魏酸酯酶,此后阿魏酸酯酶被認為是半纖維素水解酶。研究表明真菌、細菌和酵母都能分泌阿魏酸酯酶,目前從微生物中分離得到的FAE已達40多種。但絕大多數阿魏酸酯酶從真菌中分離得到,如黑曲霉(Aaspergillus niger)被普遍應用于科學研究,隨著研究深入和技術水平先進化,其它的產酶菌種逐漸被發現如泡盛曲霉(Aspergillus awamori)、鏈霉菌(如 Streptomyces avermitilis)、米曲霉(Aspergillus oryzae)、乳酸桿菌(Lactobacilli)等。目前的研究發現,不同微生物產阿魏酸酯酶的酶學特性有著較大的差異,來源于真菌阿魏酸酯酶酶活性較好。部分阿魏酸酯酶的微生物發酵來源以及培養條件詳見表1。
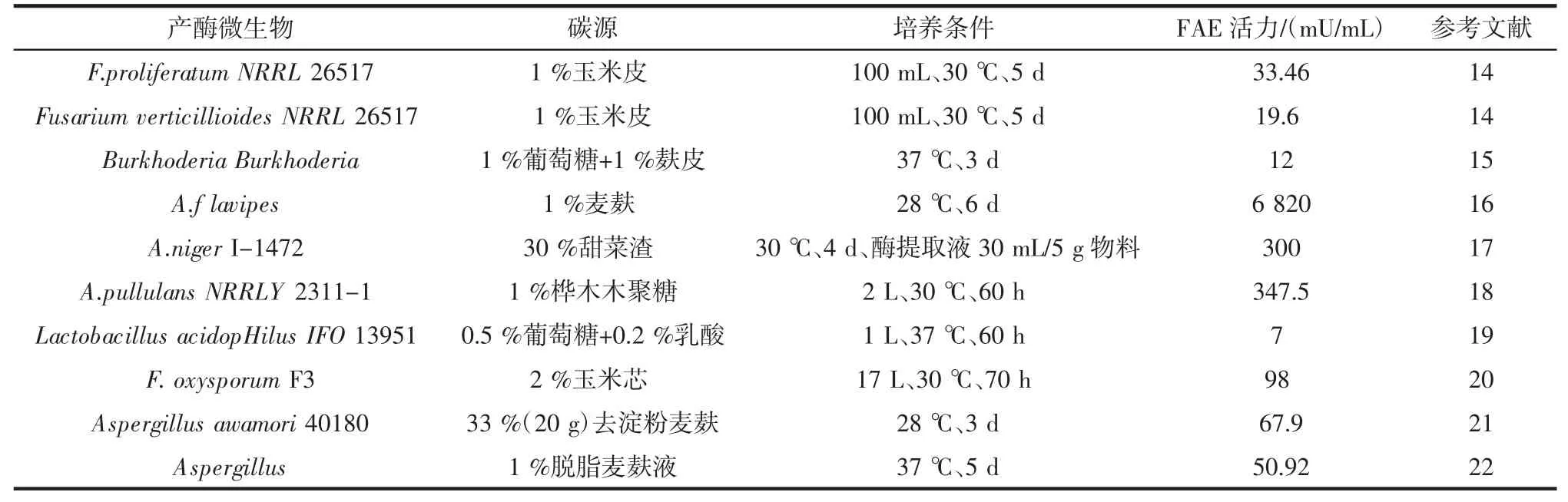
表1 自2000年以來部分產阿魏酸酯酶(FAE)的微生物來源及培養條件Table 1 Partial production of feruloyl esterase(FAE)from microbial sources and culture conditions since 2000
1.2 產阿魏酸酯酶的影響因素及理化性質
阿魏酸酯酶的真菌微生物固態發酵培養基最主要是含淀粉基和纖維素基,如甜菜渣、玉米麩、小麥麩、豆渣和醋糟。Shin等[14]以去淀粉麥麩和麥麩為發酵底物對比,發現麥麩的發酵產酶最高,接近去淀粉麥麩的2倍,表明去除麥麩中的淀粉后進行發酵產酶量會下降。并且以淀粉為唯一碳源發酵,其結果與以木糖為唯一碳源一樣,都不產阿魏酸酯酶,其原因可能與阿魏酸酯酶的誘導機制有關。麥麩中的淀粉可能僅僅有利于菌體發酵初期的生長,對阿魏酸酯酶的產生無誘導作用。阿魏酸酯鍵可能是誘導阿魏酸酯酶產生的主要因素,這一發現為進一步研究阿魏酸酯酶提供良好基礎。李干[15]經過對培養基的優化以及培養條件的測定,發現非誘導性碳源(葡萄糖、蔗糖、乳糖、甘油、阿魏酸)存在的培養基中基本不產生阿魏酸酯酶,當糖有消耗后,才開始有微量的酶活。這可能是由于糖分解代謝物的阻遏作用。目前纖維素也可用于產生阿魏酸酯酶[23]。由于不同微生物來源的FAE酶學特性有著較大的差異:大部分相對分子量在30 kDa~70 kDa之間有但是Neocallimastix MC-2所產阿魏酸酯酶相對分子質量為11 kDa;最適pH值為4~8之間,在pH 3~10范圍內穩定;最適溫度及熱穩定性為30℃~65℃;等電點在3~9.9之間不等。A.pullulans所產阿魏酸酯酶相對分子質量為210 kDa,等電點為6.5,最適pH值為6.7,最適溫度為60℃。鄧軼韜等[24]在對黑曲霉發酵去淀粉麥麩產的阿魏酸酯酶時,以葡萄糖為誘導碳源研究時發現麩皮的半纖維結構能有效的誘導產阿魏酸酯酶,并且碳氮源可以在發酵前期促進菌體快速大量生長。對于不同氮源,有機氮源如酵母提取物、蛋白胨為氮源時酶活大于硫酸銨、硝酸銨無機氮源,不同的菌株會喜好有利于自己生長和產生次級代謝產物的氮源。以1%接種量和近中性的pH值更有利于產酶。通過優化后菌株的酶活提高了37%。
1.3 產阿魏酸酯酶的分子克隆及專利
由于大多數微生物發酵產阿魏酸酯酶的量達不到工業化大規模生產的要求,在尋找新的FAE和應用來源方面,學術界和工業界都在積極研究。自2000年以來專利申請情況見表2[9]。
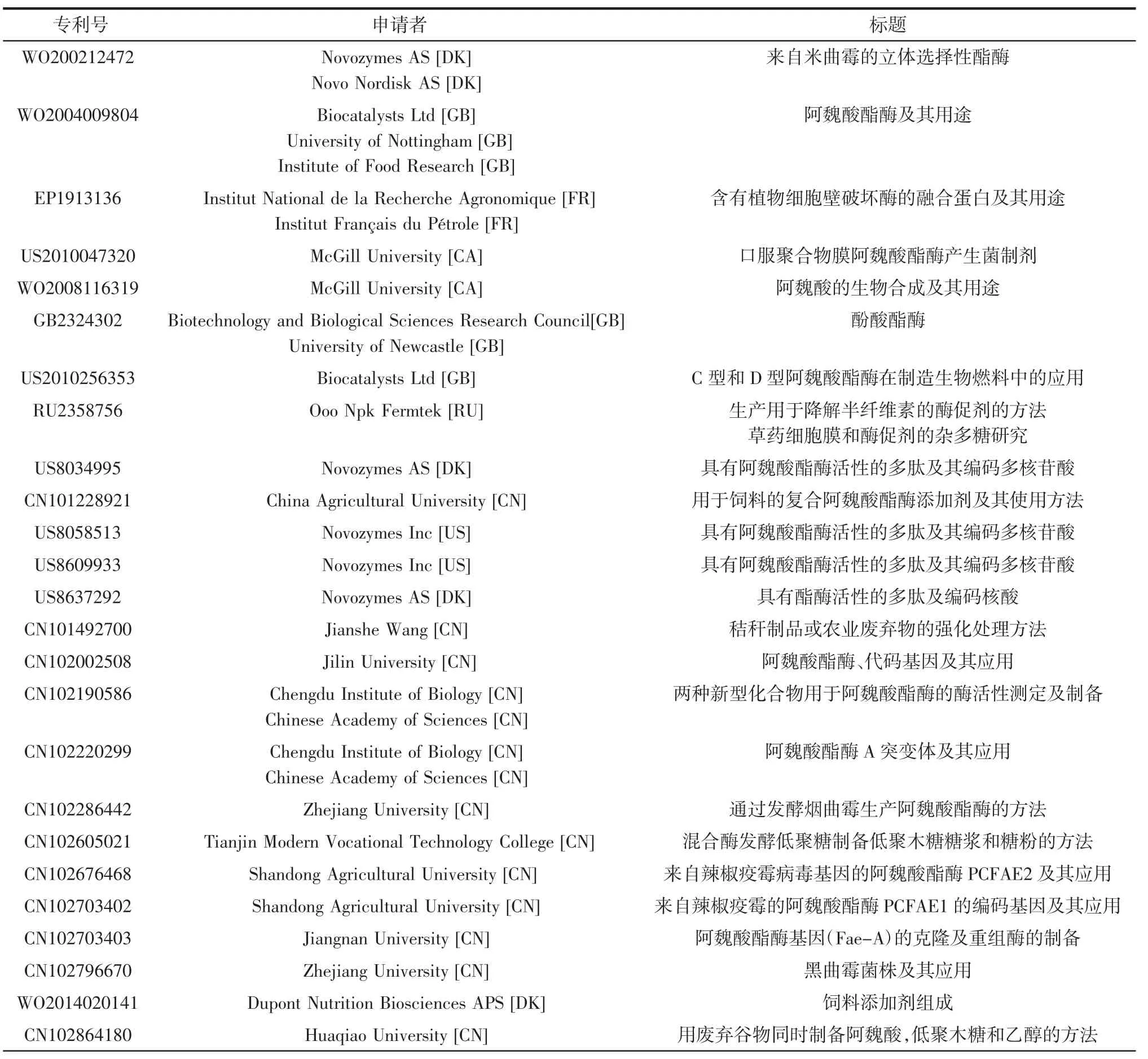
表2 自2000年以來阿魏酸酯酶申請和授予專利[9]Table 2 Ferulic acid esterase applications and patents granted since 2000
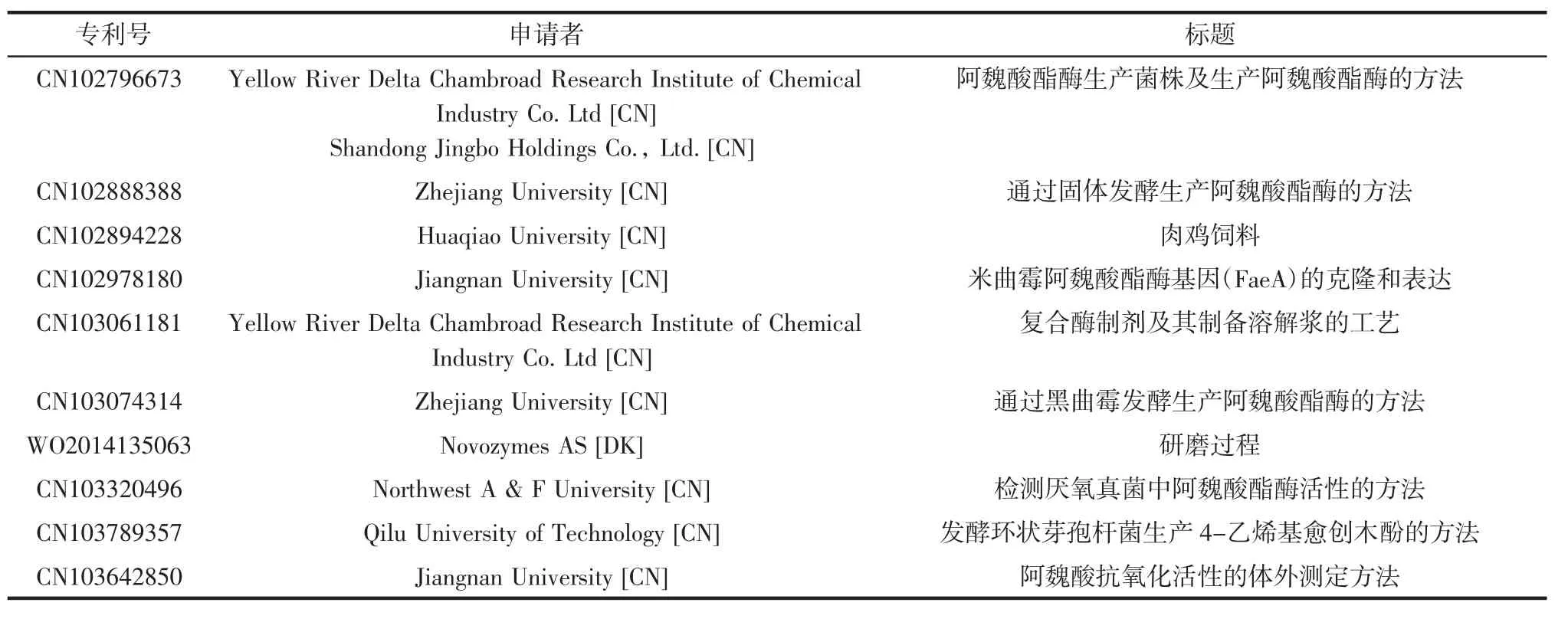
續表2 自2000年以來阿魏酸酯酶申請和授予專利[9]Continue table 2 Ferulic acid esterase applications and patents granted since 2000
專利中大部分包括微生物來源的FAE,其多肽序列,編碼多肽的核苷酸,核酸構體,載體和包含該核苷酸的宿主細胞以及生產和使用該多肽的方法。利用現代生物技術優化微生物基因,以達到提高阿魏酸酯酶產量的目的。目前不同微生物來源的FAE被異源克隆或以其他方式克隆后,發酵不同農業副產物產阿魏酸酯酶能力的研究較少。Record等[25]將一種新構建的含有黑曲霉FAE A編碼序列的cDNA載體轉入有蛋白酶基因表達缺陷的黑曲霉菌株中。以葡萄糖為碳源(60 g/L),阿魏酸酯酶活性為1.236 U/mL并且該酶易于純化。Levasseur等[26]將來自黑曲霉BRFM13的FAE B基因插入到gpd啟動子控制下的表達載體中,使其在蛋白酶基因表達缺陷的黑曲霉菌株表達。轉化后的阿魏酸酯酶活性達到1.079 U/mL,比由甜菜漿誘導的相同來源的非轉化黑曲霉菌株的產量高16倍。
Levasseur等[27]構建了一種由AnFaeA和里氏木霉組成的嵌合酶。轉入里氏木霉Rut30和CL847菌株中分別表達,發現Rut30產阿魏酸酯酶的活力比CL847菌株中的低。Abokitse等[28]研究來自溫性厭氧菌的TtFaeA轉入大腸桿菌后發酵小黑麥糠釋放阿魏酸。反應物先用木聚糖酶37℃處理16 h,表達后的阿魏酸酯酶55℃作用5 h。研究發現該酶熱穩定性高達80℃,可釋放30%的阿魏酸。Wong等[6]將來自黑曲霉的AnfaeA基因克隆并轉入到釀酒酵母中,發酵柳枝稷釋放阿魏酸約3.01 μg/100 mg(總釋放阿魏酸2 mg/g)。細菌來源的阿魏酸酯酶能從小黑麥麩皮,干燥的麥麩以及蒸汽爆炸處理后的小麥秸稈中釋放出大量的阿魏酸。Wu等[5]將來自纖維堆囊菌(Sorangium cellulosum)的阿魏酸酯酶ScFae1和ScFae2轉入大腸桿菌細胞純化后發酵小黑麥麩皮,將0.45 U/mL克隆表達后的酶與5.3 U/mL木聚糖酶液態發酵5%黑小麥麩皮懸浮液,37℃發酵3 h。阿魏酸的最大釋放量為96%。
2 釋放阿魏酸的生產工藝及影響因素
現有微生物發酵釋放阿魏酸方式主要有固體發酵與液體發酵兩種:固態發酵(solidstate fermentation,SSF)和液態發酵(submerged fermentation,SmF)。液態發酵比固態發酵過程要復雜,實際工廠生產設備投資較大。固態發酵[9]包括將固體懸浮在液體中的深層發酵和幾乎沒有游離水的濕固體材料上培養微生物的工藝過程。所用原料一般為經濟易得、富含營養物質的工農業副、廢產品,如麩皮、薯粉、大豆餅粉、高粱、玉米粉等。并且具有生產效率高、工藝簡單、操作粗放、能耗少、廢液少、產物分離較容易等優點倍受重視,有可能產生一些在液體培養中不產生的酶和其他有益代謝產物。張憬等[29]利用液態深層發酵法生產同時含有阿魏酸酯酶和阿拉伯木聚糖酶的混合酶制劑,協同作用于去淀粉麥麩制備阿魏酸和低聚糖,經3次降解后阿魏酸總得率為55.46%。由于小麥麩皮細胞壁是由纖維素、半纖維素和蛋白質等多種高聚物組成,半纖維素(主要為阿拉伯木聚糖)和木質素通過阿拉伯糖和阿魏酸等橋鏈作用形成致密的網狀結構,很難降解。由此可以推測阿魏酸酯酶、木聚糖酶和纖維素酶3種酶共同作用,同時切斷木聚糖間的酯鍵長鏈和降低纖維素間的阻礙作用,阿魏酸釋放率將會得到進一步的提高。
微生物固態發酵釋放阿魏酸的影響因素有發酵底物基質的顆粒大小、水分活度、溫度、接種濃度、pH值等。Samad K A等[30]用芽孢桿菌發酵香蕉莖稈廢物并應用共培養法釋放阿魏酸,研究幾種影響釋放阿魏酸因素的影響力。結果表明:pH值>共培養類型>接種量>攪拌>發酵時間>溫度>料液比。pH值,攪動,共培養類型和接種量對阿魏酸生產具有顯著影響。在交互性因素中,pH值和溫度與發酵時間的相互作用對阿魏酸產量的影響最大。
在植物細胞壁中阿魏酸主要以酯鍵的形式連接在阿拉伯木聚糖的阿拉伯糖側鏈殘基上,纖維素與木聚糖之間的空間阻礙會影響阿魏酸酯酶的作用能力,從而影響酶與底物的有效接觸,所以單一使用阿魏酸酯酶水解釋放阿魏酸的效率不理想。已有報道表明,阿魏酸酯酶可以協同纖維素酶以及木聚糖酶提高植物纖維質中阿魏酸的釋放量[31]。
龔燕燕等[32]用阿魏酸酯酶與木聚糖酶協同作用降解麥麩制備阿魏酸的研究中發現,木聚糖酶和阿魏酸酯酶協同作用,木聚糖酶先將細胞壁中的半纖維素降解為相對分子質量相對較小的阿魏酸寡聚多糖,改變細胞壁的物理化學性質有利于阿魏酸酯酶作用于阿魏酸寡聚多糖,能促進釋放FA,當木聚糖酶添加量為300 U時,FA的釋放率從19.48%上升至70.10%。Topakas等[33]的報道也證明阿魏酸酯酶是釋放阿魏酸的關鍵酶,木聚糖的添加能促進阿魏酸酯酶進一步釋放阿魏酸,兩者具有良好的協同作用,能有效降解小麥麩皮,提高阿魏酸的釋放率。
曾妍等[34]用阿魏酸酯酶與纖維素酶協同降解麥麩釋放阿魏酸,單獨使用阿魏酸酯酶30 U時阿魏酸的釋放率僅為4.1%,單獨使用纖維素酶并不釋放阿魏酸,當兩種酶協同作用時,阿魏酸的釋放率明顯提高,經單因素試驗確定雙酶協同作用的最佳條件為:阿魏酸酯酶的最適添加量為30U協同纖維素酶的最適添加量為70 U,此時阿魏酸釋放率達到22.9%,是阿魏酸單獨作用時的5.58倍,水解時間為10 h,水解溫度為40℃,水解pH值為5.0,料液質量體積比為1∶30(g/mL),此時阿魏酸的釋放率為23.6%。
Wang 等[31]從嗜酸乳酸桿菌(Lactobacillus acidophilus)中分離得到的一種新型阿魏酸酯酶,單獨使用水解100 mg去淀粉麥麩并不釋放阿魏酸,分別與木聚糖酶和α-L-阿拉伯糖苷酶協同作用最大可以得到12.4 nmol和3.64 nmol的阿魏酸,3種酶共同作用最大可以得到15.7 nmol的阿魏酸,證明以木聚糖酶為主,α-L-阿拉伯呋喃糖苷酶為輔可以協同作用阿魏酸酯酶促進阿魏酸的釋放。
Wu Hongli等[35]用木聚糖酶(AnXyn11A)和阿魏酸酯酶協同降解麥麩,得到產率較高的阿魏酸和低聚木糖。AnXyn11A在pH 5.0和60℃時最大活性為240 U/mL。AnFaeA在pH 5.0和45℃時最大活性為21 U/mL的。AnXyn11A和AnFaeA協同作用于去淀粉麥麩(DSWB),釋放的FA與單獨酶作用下(僅釋放FA的16.8%)相比增加至70%。
Wu等[5]用在大腸桿菌中表達的新型阿魏酸酯酶ScFAE2發酵黑小麥麩皮即得阿魏酸為2.22 mg/g;Benoit I等[36]用來自黑曲霉的阿魏酸酯酶發酵玉米麩皮獲得阿魏酸的得率是31.22 mg/g;Xie C等[37]用來自猴頭菌的阿魏酸酯酶發酵麥麩即得阿魏酸4 mg/g;Schmidt等[38]用米根霉(Rhizopus oryzae)固態發酵的方法發酵米糠,從未發酵前33 mg/g到發酵后含量為765 mg/g。
3 結束語及展望
對農林廢棄物中阿魏酸的研究一直是糧油加工、生物工程等學科的研究熱點之一。微生物采用固態發酵降解麥麩、米糠等廢棄物釋放阿魏酸比液態發酵有更好的可控條件,但是由于不同微生物來源的阿魏酸酯酶的酶特異性不同,所以對發酵條件如固態基質的選擇、溫度、pH值、料液比等的最優選擇亟待解決。
目前,微生物直接發酵生產阿魏酸得率不佳,表明距離具體工業化生產仍有一定的差距所以在發酵過程中可采取如蒸汽爆破等輔助手段來提高阿魏酸的釋放率。所以要使農林廢棄物中阿魏酸在工業上得到充分生產應用,不僅要在現有研究的基礎上應用現代生物技術對產阿魏酸酯酶的基因進行改良還要找到生產過程中每一個步驟的有效契合點,對工藝參數進行分析、構建、優化和創新。找到合適的發酵條件、培養條件,建立一套實用性強、節約成本的生產方法。
