麥角硫因機(jī)體內(nèi)的分布與代謝和其在疾病中的作用研究進(jìn)展
2020-01-08 05:59:08潘虹余郭麗瓊林俊芳
食品科學(xué) 2019年23期
潘虹余,郭麗瓊,2,林俊芳,2,
(1.華南農(nóng)業(yè)大學(xué)食品學(xué)院,廣東 廣州 510642;2.廣東省微生態(tài)制劑工程技術(shù)研究中心,廣東 廣州 510640)
1909年,有學(xué)者在研究麥角真菌Claviceps purpurea時(shí)分離出一種獨(dú)特的白色結(jié)晶含硫化合物,后來(lái)被確定為2-硫基-L-組氨酸三甲基內(nèi)鹽[1],隨后其被命名為麥角硫因(ergothioneine,EGT)。EGT可由非酵母真菌、放線菌目和某些藍(lán)細(xì)菌合成[1]。動(dòng)物機(jī)體自身不能合成EGT,只能從食物中攝取并積累在組織和細(xì)胞中。EGT具有很強(qiáng)的抗氧化性,在機(jī)體內(nèi)扮演著重要的角色。隨著人們對(duì)抗氧化劑研究的不斷深入,EGT作為一種天然的生物活性物質(zhì)受到人們廣泛的關(guān)注,國(guó)外已有相關(guān)報(bào)道,而國(guó)內(nèi)對(duì)其研究還處于探索階段[2]。目前為止,鮮有研究探討EGT在人類受試者中的藥代動(dòng)力學(xué)[2]。
1 EGT的結(jié)構(gòu)、性質(zhì)和生物合成
EGT是一種無(wú)味、無(wú)色化合物,分子式為C9H16N3O2S,25 ℃時(shí)水溶解度極限值為0.9 mol/L[3],分子結(jié)構(gòu)中含有咪唑-2-硫酮基團(tuán)。EGT在溶解狀態(tài)下存在硫醇和硫酮兩種結(jié)構(gòu)的互變異構(gòu)體(圖1)。在生理pH值條件下EGT主要以硫酮形式存在于水溶液中,因此與其他天然存在的硫醇(例如谷胱甘肽、N-乙酰半胱氨酸)相比,EGT是非常穩(wěn)定的抗氧化劑,在生理pH值下不易發(fā)生自氧化。在pH值為7時(shí),EGT的標(biāo)準(zhǔn)氧化還原電勢(shì)為-60 mV,其他天然存在的硫醇氧化還原電勢(shì)在-200~-320 mV之間[4]。硫酮硫醇互變異構(gòu)體和高氧化還原電位是EGT具有更高穩(wěn)定性的原因,這種特性可防止EGT在生理pH值下像其他硫醇(如谷胱甘肽(glutathione,GSH))一樣進(jìn)行自氧化[4]。
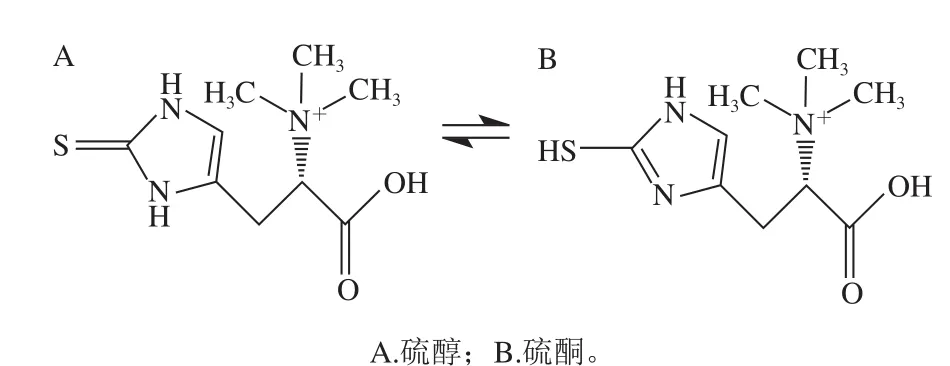
圖1 EGT的硫醇和硫酮的互變異構(gòu)體Fig. 1 Structures of thione-thiol tautomers of EGT
Seebeck首次探討了合成EGT的基因簇(egt A/B/C/D/E)在分枝桿菌屬中的應(yīng)用[10],并且在許多真核生物和細(xì)菌中發(fā)現(xiàn)了EGT合成的關(guān)鍵基因,表明能夠產(chǎn)生EGT的微生物不僅限于上文所述[10-11]。Seebeck[10]推測(cè)這5 個(gè)基因簇(egt A/B/C/D/E)主要負(fù)責(zé)分枝桿菌中EGT的生物合成(圖2)。EGT的生物合成從L-組氨酸開(kāi)始,通過(guò)中間體前體組氨酸三甲基內(nèi)鹽,分別從半胱氨酸和蛋氨酸獲取硫和甲基以形成最終產(chǎn)物EGT[12]。EGT生物合成中存在兩個(gè)主要反應(yīng),即氧化C—S鍵的形成和C—S鍵的裂解反應(yīng),使硫原子從半胱氨酸或c-谷氨酰半胱氨酸完全轉(zhuǎn)移至組氨酸的咪唑側(cè)鏈[13]。2018年,Osawa等[14]在大腸桿菌中成功表達(dá)了恥垢分枝桿菌中的EGT合成酶基因egt A、egt B、egt C、egt D、egt E,使重組菌的EGT產(chǎn)量達(dá)到了24 mg/L。
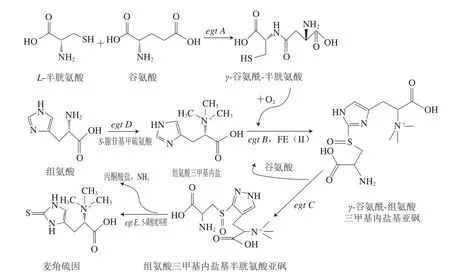
圖2 EGT在分枝桿菌中的生物合成路徑[10]Fig. 2 Biosynthetic pathway of EGT in mycobacteria[10]
2 EGT在體內(nèi)的攝取、分布和代謝
2.1 EGT的攝取和分布
盡管EGT存在于許多細(xì)胞中,但其本身不能跨越細(xì)胞質(zhì)膜,需要特定的載體將其運(yùn)輸至細(xì)胞內(nèi)[15]。2005年,Gründemann等[16]在動(dòng)物組織中發(fā)現(xiàn)了一種Na+依賴型EGT轉(zhuǎn)運(yùn)蛋白——有機(jī)陽(yáng)離子轉(zhuǎn)運(yùn)蛋白1型(organic cation transporter novel type-1,OCTN1),并使用液相色譜-質(zhì)譜差異著色法確定了EGT是OCTN1的關(guān)鍵底物。OCTN1的特異性表現(xiàn)在除了EGT外對(duì)其他有機(jī)陽(yáng)離子攝取和轉(zhuǎn)運(yùn)率均很低,包括與EGT結(jié)構(gòu)相似的化合物,例如甲巰咪唑和組氨酸三甲基內(nèi)鹽[17-18]。OCTN1對(duì)EGT的轉(zhuǎn)運(yùn)米氏常數(shù)(Km)很低(約為21 mmol/L),說(shuō)明EGT是其特殊的結(jié)合底物[17-18]。
特定轉(zhuǎn)運(yùn)蛋白的存在表明動(dòng)物(包括人類)可以從飲食中攝取EGT并通過(guò)在組織中積累而獲益。EGT有自己的選擇性轉(zhuǎn)運(yùn)蛋白用于腸道的攝取和輸送到各組織[19]。早期研究揭示了EGT在組織中的積累具有性別差異[20](可能與睪酮水平有關(guān))和年齡差異,EGT在體內(nèi)組織中的水平具有與年齡增長(zhǎng)相關(guān)的下降趨勢(shì)[21-22]。盡管EGT在人體組織中分布較廣泛,但它目前不被認(rèn)為是必需的營(yíng)養(yǎng)成分,因?yàn)闆](méi)有研究發(fā)現(xiàn)其缺乏會(huì)引起不良反應(yīng)。最近研究發(fā)現(xiàn)EGT的基線水平在肝臟和血細(xì)胞中最高,比腦和眼睛中高約10~20 倍,表明這些組織可能是體內(nèi)EGT積累的主要部位[23]。對(duì)體內(nèi)EGT分布的研究表明,EGT優(yōu)先積聚在易受高水平氧化應(yīng)激和炎癥影響的器官、細(xì)胞和分泌物中,如肝臟、腎臟、紅細(xì)胞[2,23]、眼晶狀體和精液[15],這可能是一種適應(yīng)性的細(xì)胞保護(hù)機(jī)制[17,24]。
2.2 EGT的代謝
EGT的代謝轉(zhuǎn)換率相對(duì)緩慢,在禁食1 周的大鼠中觀察到其在血液和肝臟中的水平?jīng)]有明顯下降[25],EGT的這種高積累和緩慢代謝的特征歸因于腎臟重吸收和EGT的低尿排泄[2,26]。人體研究顯示,尿液中EGT的排泄量很小,表明EGT在口服后主要在體內(nèi)保留[2]。
Servillo等[27]研究了EGT的可能氧化產(chǎn)物,并提出組氨酸三甲基內(nèi)鹽和EGT-磺酸是EGT的穩(wěn)定氧化產(chǎn)物。Cheah等[2]測(cè)定了組織中3 種與EGT相關(guān)代謝物(組氨酸三甲基內(nèi)鹽、EGT-磺酸、S-甲基-EGT)的分布水平,發(fā)現(xiàn)僅能檢測(cè)到組氨酸三甲基內(nèi)鹽,并且其含量與各組織中的EGT濃度顯著相關(guān)[23]。這可能表明組氨酸三甲基內(nèi)鹽確實(shí)是體內(nèi)EGT的代謝產(chǎn)物。研究發(fā)現(xiàn)EGT的積累并不總是與上述氧化產(chǎn)物的水平相關(guān),例如肝臟中EGT含量最高,但檢測(cè)不出EGT-磺酸,而肺中EGT的含量處于中等水平,卻檢測(cè)出較高水平的EGT-磺酸[2]。
3 EGT的抗氧化特性
EGT的強(qiáng)抗氧化性可能與以下4 種分子活動(dòng)有關(guān):直接清除活性氧(reactive oxygen species,ROS)[28];有效螯合各種二價(jià)金屬陽(yáng)離子[29];激活抗氧化酶(如谷胱甘肽過(guò)氧化物酶和符胱什肽過(guò)氧化物酶),并抑制超級(jí)氧化激酶,如NADPH-細(xì)胞色素c還原酶;抑制各種血紅素蛋白(如血紅素和肌紅蛋白)的氧化作用。
EGT與二價(jià)金屬陽(yáng)離子例如Cu2+、Hg2+、Co2+、Fe2+形成無(wú)氧化還原活性的絡(luò)合物[30],其中最穩(wěn)定的是銅,其具有最高的絡(luò)合物形成常數(shù)[29]。這些金屬離子在體內(nèi)的結(jié)合可能有助于防止它們參與ROS的產(chǎn)生。
以前如果想買(mǎi)私募基金或者信托產(chǎn)品,只要滿足100萬(wàn)元到300萬(wàn)元人民幣不等的資金門(mén)檻就行,所謂的合格投資者審查基本上是一個(gè)形式。而資管新規(guī)實(shí)施后,無(wú)論是銀行、券商還是信托、私募基金,發(fā)行的資管產(chǎn)品將被嚴(yán)格分為公募和私募兩類。公募產(chǎn)品誰(shuí)都能買(mǎi),而私募產(chǎn)品僅供合格投資者。這里說(shuō)的合格投資者不但必須具有2年以上投資經(jīng)歷,并且需要家庭金融資產(chǎn)至少在300萬(wàn)元到500萬(wàn)元以上,或者是投資者本人三年的年均收入在40萬(wàn)元以上。要求顯然比以前提高不少。換句話說(shuō),目前的很多高凈值客戶以后只夠格購(gòu)買(mǎi)公募產(chǎn)品了。想買(mǎi)定制化、投資非標(biāo)或者非上市股權(quán)的私募產(chǎn)品,需滿足合格投資者的要求。
EGT具有調(diào)節(jié)炎癥[31]、防止紫外線輻射誘導(dǎo)的損傷[32-33]以及其他保護(hù)細(xì)胞活動(dòng)的功能[34]。與其他硫醇相比,EGT還顯示出更高的速率(速率常數(shù)為2.3×107L/(mol·s)從而使單線態(tài)氧失活[35]。相比之下,EGT與超氧化物和H2O2不能很快地進(jìn)行反應(yīng),與超氧化物反應(yīng)的速率常數(shù)通常低于103L/(mol·s)[36]。EGT對(duì)由亞硝基鐵氰化鈉(硝普鈉)產(chǎn)生的NO·誘導(dǎo)的細(xì)胞死亡沒(méi)有影響,表明它與NO·的反應(yīng)很緩慢或沒(méi)有反應(yīng)[37]。
研究表明,EGT可能作為適應(yīng)性抗氧化劑在體內(nèi)發(fā)揮抗氧化作用。EGT在健康動(dòng)物中的抗氧化作用有限,但是在由組織損傷引起的自由基活性物質(zhì)水平升高、存在ROS/活性氮(reactive nitrogen species,RNS)產(chǎn)生的毒素或涉及氧化應(yīng)激引起的疾病程度加重時(shí),EGT會(huì)發(fā)揮強(qiáng)有力的抗氧化作用[15-16]。通過(guò)檢測(cè)血漿和尿液中的氧化損傷標(biāo)準(zhǔn)生物指標(biāo)水平,發(fā)現(xiàn)EGT的攝入可減少由不同物質(zhì)引起的氧化損傷,包括尿囊素(尿酸鹽氧化)、8-羥基脫氧鳥(niǎo)苷(DNA損傷)、8-iso-PGF2a(脂質(zhì)過(guò)氧化)、蛋白質(zhì)羰基化和C-反應(yīng)蛋白(蛋白質(zhì)氧化),但減少程度不明顯[2]。EGT的這種特性可能具有重要意義,即在不干擾ROS/RNS在健康組織中重要作用的情況下發(fā)揮抗氧化作用[38-40];當(dāng)氧化損傷變得嚴(yán)重時(shí),EGT則在體內(nèi)組織中大量積累并發(fā)揮抗氧化作用[41]。
研究發(fā)現(xiàn)某些在體外具有強(qiáng)大抗氧化活性的物質(zhì)在體內(nèi)不發(fā)揮作用,例如許多類黃酮[42]。EGT在細(xì)菌、真菌[5-6,14,43]、秀麗隱桿線蟲(chóng)和動(dòng)物體內(nèi)也具有抗氧化作用。例如,缺乏EGT轉(zhuǎn)運(yùn)蛋白的秀麗隱桿線蟲(chóng)表現(xiàn)出較高的蛋白質(zhì)羰基化水平(蛋白質(zhì)氧化損傷的生物指標(biāo))[42,44];在缺乏EGT的斑馬魚(yú)中,8-羥基-2-多氧鳥(niǎo)苷和8-二氫鳥(niǎo)嘌呤(一種核酸氧化損傷的生物標(biāo)志物)的水平增加[42,45]。
4 EGT的藥代動(dòng)力學(xué)
4.1 皮膚保護(hù)功能
EGT在皮膚細(xì)胞和離體皮膚的表皮細(xì)胞中呈現(xiàn)時(shí)間和濃度依賴性積累[46]。與具有較高氧化自由基吸收能力但生物利用度低和易被快速清除的抗氧化劑(如白藜蘆醇)相比,EGT具有較好的生物利用度[47]。EGT不僅可以防止氧化損傷[32],還可對(duì)經(jīng)紫外線照射的細(xì)胞進(jìn)行DNA修復(fù),防止單鏈DNA斷裂[48],抑制細(xì)胞凋亡的發(fā)生,并使細(xì)胞活力增加[46,48]。細(xì)胞吸收、積累和利用L-EGT的能力使EGT及其受體/轉(zhuǎn)運(yùn)蛋白OCTN1成為皮膚抗氧化防御系統(tǒng)的重要組成部分[46]。
4.2 抗抑郁癥
最近的研究發(fā)現(xiàn),口服攝入的EGT可通過(guò)血腦屏障轉(zhuǎn)運(yùn)到大腦中,并在小鼠體內(nèi)發(fā)揮類似抗抑郁藥的作用。對(duì)小鼠進(jìn)行強(qiáng)迫游泳實(shí)驗(yàn)(forced swimming test,F(xiàn)ST)和懸尾實(shí)驗(yàn)(tail suspension test,TST),發(fā)現(xiàn)與正常飲食對(duì)照組小鼠相比,EGT顯著縮短了處理組小鼠在FST和TST中的不動(dòng)時(shí)間[49]。口服攝入EGT后,腦內(nèi)EGT濃度呈劑量依賴性增加[49]。EGT有兩種作用:抑制神經(jīng)干細(xì)胞的增殖和促進(jìn)神經(jīng)元分化[49-50]。抗抑郁藥物如氟西汀、丙咪嗪和瑞波西汀可促進(jìn)神經(jīng)新生,表明神經(jīng)新生和抗抑郁作用之間可能存在關(guān)聯(lián)[49,51]。EGT可能是通過(guò)促進(jìn)神經(jīng)元的分化來(lái)發(fā)揮其在體內(nèi)的抗抑郁作用,但這一點(diǎn)需要進(jìn)一步的研究來(lái)證實(shí)[49]。
4.3 眼部疾病
研究表明,EGT可以保護(hù)角膜內(nèi)皮細(xì)胞免受氧化應(yīng)激和未折疊蛋白反應(yīng)[52]。最近發(fā)現(xiàn)EGT在人類淚液和房水樣本中具有較高的積累水平(分別為約0.35 μmol/L和約28 μmol/L),并且證明EGT很容易在小鼠眼內(nèi)積累[23]。早期研究發(fā)現(xiàn)在患有白內(nèi)障的生物機(jī)體中,晶狀體和角膜中的EGT水平顯著降低;隨著白內(nèi)障程度的增加,EGT水平持續(xù)下降[53]。另一項(xiàng)研究發(fā)現(xiàn)用EGT治療可以有效地保護(hù)小雞免受糖皮質(zhì)激素誘導(dǎo)的白內(nèi)障損傷[54]。
4.4 心血管疾病
已知心肌缺血-再灌注損傷(ischemia-reperfusion injury,IR)可產(chǎn)生ROS/RNS,而ROS/RNS過(guò)度形成會(huì)加劇IR,導(dǎo)致氧化肌細(xì)胞損傷[55-56]。EGT在IR期間可保護(hù)心臟和肝臟組織免受損傷[57-58]。EGT保護(hù)心臟的機(jī)制之一可能是防止ROS/RNS氧化肌紅蛋白,形成具有細(xì)胞毒性的鐵蛋白肌紅蛋白;EGT還能降低組織丙二醛水平并調(diào)節(jié)熱休克蛋白70和促炎細(xì)胞因子(腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)和白細(xì)胞介素-1β(interleukin-1β,IL-1β))[58]。此外,EGT作為鐵和銅的螯合物可通過(guò)減少I(mǎi)R期間自由基的產(chǎn)生來(lái)保護(hù)機(jī)體組織,這可能是EGT的另一種保護(hù)機(jī)制[59]。在小鼠體內(nèi),EGT容易在血液中累積并進(jìn)入心臟,因此它可以發(fā)揮保護(hù)心臟的作用[2,23]。
4.5 腎臟疾病與糖尿病
最近一項(xiàng)研究發(fā)現(xiàn),敲除編碼OCTN1基因的慢性腎病(chronic kidney disease,CKD)小鼠模型出現(xiàn)腎纖維化惡化的癥狀,同時(shí)CKD患者的血液中EGT水平較低[60]。CKD會(huì)增加氧化損傷[61],EGT可能通過(guò)在腎臟中的積累來(lái)減少這種損害。
糖尿病和糖尿病并發(fā)癥與氧化和糖化應(yīng)激增加密切相關(guān),部分原因是高血糖的促氧化作用[62]。EGT能夠保護(hù)內(nèi)皮細(xì)胞和大鼠嗜鉻細(xì)胞瘤(PC12)細(xì)胞免受高葡萄糖水平引起的損傷[63-64];對(duì)患糖尿病的妊娠大鼠補(bǔ)充EGT后減少了畸形胚胎的發(fā)生率[65],這歸因于EGT對(duì)高血糖依賴性氧化應(yīng)激的調(diào)節(jié)[65]。
4.6 慢性炎癥疾病
EGT在炎癥中的作用頗有爭(zhēng)議。研究發(fā)現(xiàn),輕度類風(fēng)濕性關(guān)節(jié)炎(rheumatoid arthritis,RA)患者體內(nèi)的紅細(xì)胞和單核細(xì)胞EGT水平顯著升高,并且其與CD14+細(xì)胞中OCTN1 mRNA的表達(dá)密切相關(guān)[66]。在由膠原誘導(dǎo)的關(guān)節(jié)炎小鼠發(fā)炎關(guān)節(jié)的血液和免疫組織中也發(fā)現(xiàn)了高度表達(dá)的OCTN1[67]。與RA類似,對(duì)克羅恩病(Crohn's disease,CD)患者發(fā)炎回腸黏膜的分析中發(fā)現(xiàn)EGT水平顯著升高;相應(yīng)地,與非CD受試者相比,CD患者的OCTN1 mRNA表達(dá)也有增加[68]。
在培養(yǎng)的紅細(xì)胞中,對(duì)OCTN1基因的沉默抑制了EGT攝取,并促進(jìn)了細(xì)胞凋亡的發(fā)生[69]。在這些慢性炎癥狀態(tài)中,可能是由于EGT的抗細(xì)胞凋亡特性可介導(dǎo)免疫細(xì)胞的存活并刺激炎癥的發(fā)生,進(jìn)而導(dǎo)致EGT在體內(nèi)組織中水平的增加[68]。一些研究表明,EGT水平升高的另一個(gè)原因可能是由于細(xì)胞因子介導(dǎo)的OCTN1表達(dá)上調(diào)。用TNF-α孵育Caco-2細(xì)胞,結(jié)果顯示通過(guò)TNF-α受體-1的激活增加了OCTN1表達(dá),因此EGT的增加可能是通過(guò)上述提到的延長(zhǎng)免疫細(xì)胞存活來(lái)進(jìn)一步促進(jìn)炎癥反應(yīng)[68]。類似的,與RA相關(guān)的炎性細(xì)胞因子例如IL-1β和轉(zhuǎn)錄因子RUNX1能夠上調(diào)OCTN1的表達(dá)。然而,這種反應(yīng)是否與發(fā)病機(jī)制相關(guān)或作為細(xì)胞防御的反作用反應(yīng),以及EGT的分子作用仍未確定。
4.7 神經(jīng)退行性疾病
EGT在大腦中的功能尚不確定,但在腦和各種神經(jīng)細(xì)胞中能夠觀察到OCTN1表達(dá)[70]。EGT滲透血腦屏障的能力表明EGT能在中樞神經(jīng)系統(tǒng)和神經(jīng)元中發(fā)揮功能性作用。EGT可通過(guò)激活p38 MAPK細(xì)胞內(nèi)抗氧化途徑以保護(hù)PC12細(xì)胞免受H2O2誘導(dǎo)的細(xì)胞損傷[71],還可抑制由β-淀粉樣蛋白和順鉑誘導(dǎo)的神經(jīng)元細(xì)胞凋亡[55]。研究表明,輕度認(rèn)知受損受試者和帕金森病患者的血液中EGT水平顯著低于同年齡的健康個(gè)體[72-73]。這可通過(guò)多種機(jī)制引起,例如飲食的改變和/或OCTN1活性的變化。最近有學(xué)者提出蘑菇攝入量的增加與癡呆發(fā)病率降低之間存在相關(guān)性[74-75]。然而蘑菇含有多種可能具有生物活性的化合物,不能認(rèn)為是EGT發(fā)生作用。因此,仍然需要更多研究來(lái)討論EGT是否對(duì)神經(jīng)退行性疾病產(chǎn)生有益作用。
5 EGT的安全性
迄今為止對(duì)EGT的毒理學(xué)研究未發(fā)現(xiàn)其有副作用[76]。然而,一些人認(rèn)為在慢性炎癥中,EGT的積累會(huì)加重炎癥反應(yīng)[68]。為證明發(fā)炎和受損組織是否會(huì)自發(fā)增加EGT水平使其自身受到進(jìn)一步傷害,推測(cè)細(xì)胞可通過(guò)EGT的積累來(lái)進(jìn)行自我保護(hù),有助于保護(hù)組織免受氧化損傷的進(jìn)一步傷害。因此,可以推斷在一些人類疾病中觀察到的EGT在組織中的積累不是病理性的,而是細(xì)胞保護(hù)性的[77-80]。例如一項(xiàng)代謝組學(xué)研究確定了壓力超負(fù)荷和梗塞的小鼠心臟中EGT水平顯著增加[81],這可能反映了機(jī)體組織能通過(guò)積累EGT來(lái)保護(hù)自身免受進(jìn)一步的傷害。
2017年,歐洲食品安全局發(fā)布報(bào)告提出EGT作為膳食補(bǔ)充劑和強(qiáng)化食品攝入不會(huì)加劇RA、CD和糖尿病的易感性和病癥發(fā)展;同時(shí),其下屬的營(yíng)養(yǎng)產(chǎn)品、營(yíng)養(yǎng)及致敏科學(xué)組提出EGT的嬰兒安全每日限量為2.82 mg/kg mb,兒童為3.39 mg/kg mb,成人(包括孕婦和哺乳期婦女)為1.31 mg/kg mb[82]。因此新型合成L-EGT在安全食用水平下對(duì)成人(孕婦)、兒童和嬰兒均是安全的。
6 結(jié) 語(yǔ)
雖然近年來(lái)對(duì)EGT的研究不斷取得進(jìn)展,并有大量研究證明EGT在體外具有多種抗氧化和細(xì)胞保護(hù)作用;但是EGT的真正生理作用尚未完全闡明,這些細(xì)胞保護(hù)作用背后的分子機(jī)制仍然未知。同時(shí),由于體外研究中采用的條件與體內(nèi)不同,體外研究結(jié)果不能完全應(yīng)用于體內(nèi)。因此,仍需要大量的體內(nèi)研究來(lái)驗(yàn)證EGT的治療潛力。
此外,不僅某些真菌,一些致病微生物、病原體也會(huì)產(chǎn)生EGT并利用它來(lái)保護(hù)自身免受宿主防御(包括ROS/RNS)的傷害[43,67],例如結(jié)核分枝桿菌(人結(jié)核病的致病因子),而其EGT合成能力的喪失也會(huì)降低它們的毒力。因此,EGT生物合成途徑的抑制劑(應(yīng)該是一種可行的治療方法,因?yàn)檫@種途徑不會(huì)在人體中發(fā)生)可能是攻擊致病性合成EGT生物的新型藥物潛在來(lái)源。
由于EGT在生物體內(nèi)的作用機(jī)制及與疾病的關(guān)系還不清楚,限制了其生產(chǎn)和應(yīng)用。因此需要對(duì)EGT作為免疫調(diào)節(jié)劑和某些疾病的治療劑開(kāi)展更多的研究,探討其在體內(nèi)的作用機(jī)制,以推進(jìn)EGT在疾病治療上的應(yīng)用。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
天天愛(ài)科學(xué)(2022年9期)2022-09-15 01:12:54
天天愛(ài)科學(xué)(2022年4期)2022-05-23 12:41:48
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
當(dāng)代水產(chǎn)(2022年3期)2022-04-26 14:26:56
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
航空世界(2020年10期)2020-01-19 14:36:20
人大建設(shè)(2019年12期)2019-05-21 02:55:32
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
七彩語(yǔ)文·畫(huà)刊(2012年3期)2012-04-29 00:00:00
