增齡對大鼠睪丸自噬水平和血睪屏障完整性的影響
2020-01-09 07:36:38趙海霞馬瓊艷楊思琪張長城
中國藥理學通報 2020年1期
尤 旭,趙海霞,馬瓊艷,楊思琪,張 艷,楊 圓,葉 勇,袁 丁,張長城
(三峽大學醫學院,湖北 宜昌 443002)
隨著社會經濟的發展、生活環境的變化和人們生育觀念的改變,選擇晚育的夫婦越來越多。然而,隨著年齡的不斷增加,男性睪丸組織結構和功能會出現不同程度的衰退,生精功能逐漸下降,精子數量和質量均明顯下降,精子形態發生異常,男性不育和子代畸形率也大幅上升。研究顯示[1-2],睪丸血睪屏障完整性被破壞是導致衰老睪丸生精功能衰退的重要因素,而睪丸自噬功能受損又是導致血睪屏障完整性被破壞的重要原因[3]。然而,衰老過程中睪丸自噬水平是否發生變化及如何變化,尚無相關文獻報道。本研究探討了衰老過程中睪丸自噬水平的變化及其對血睪屏障完整性的影響。
1 材料與方法
1.1 實驗動物SPF級♂Sprague-Dawley(SD)大鼠,購于北京維通利華實驗動物技術有限公司,許可證號SCX K(京)2012-0001,分籠飼養于三峽大學實驗動物中心,室溫(23±3)℃,濕度(60±5)%,12 h陰暗交替,自由攝食和飲水。
1.2 主要材料與試劑β-actin抗體(貨號#4970)和β-catenin抗體(貨號#8814),購自Cell Signaling Technology公司;ATG5抗體(貨號10181-2-AP)和LC3抗體(貨號14600-1-AP),購自Proteintech公司;ATG7抗體(貨號500691),購自正能抗體公司;Beclin1抗體(貨號ab207612),購自Abcam公司;Occludin抗體(貨號sc-5562),購自Santa Cruz公司;羊抗鼠、羊抗兔和兔抗羊二抗及牛血清白蛋白(bovine serum albumin,BSA),購自武漢科瑞有限公司;Alexa Fluor 488驢抗兔 IgG(H+L)抗體,購自Jackson ImmunoResearch公司;4’,6-二脒基-2-苯基吲哚(4′,6-diamidino-2-phenylindole,DAPI),購自Life Technologies公司;BCA蛋白定量試劑盒,購自北京普利萊基因技術有限公司;BeyoECL Plus超敏ECL化學發光試劑盒,購自碧云天生物技術研究所;高蛋白脫脂高鈣奶粉,購自內蒙古伊利實業集團股份有限公司。
1.3 主要儀器LEICA TP 1020全自動脫水機和ULTRACUT R超薄切片機購于德國Leica公司;攤片烤片機購于上海寰熙醫療器械有限公司;BX53顯微鏡購于日本Olympus公司;A1R+激光共聚焦顯微鏡購于日本Nikon公司;JA2003電子分析天平購于上海天平儀器廠;電動勻漿器購于常州德杜精密儀器有限公司;CT15RT高速冷凍離心機購于上海天美生化儀器設備工程有限公司;PK-600S水浴鍋購于上海精宏實驗設備有限公司;51118650酶標儀購于美國Thermo公司;PowerPacTMBasic電泳儀和Mini PROTEAN3 Cell電泳槽購于美國Bio-Rad公司;Bioshine ChemiQ4800 mini化學發光凝膠成像顯影設備購于上海歐翔科學儀器有限公司。
1.4 方法
1.4.1動物分組與處理 分別購買不同月齡的♂SD大鼠,即6月齡、12月齡、18月齡和24月齡,每個月齡組大鼠各10只,稱取大鼠體重,腹腔注射20%烏來糖麻醉后處死,迅速取出兩側睪丸組織,稱重,右側睪丸組織迅速過液氮,儲存于-80 ℃超低溫冰箱中備用,左側睪丸放于4%多聚甲醛中固定。
1.4.2HE染色觀察大鼠睪丸組織形態學的變化 將睪丸取出,4%多聚甲醛中固定24~48 h后,常規梯度乙醇脫水、二甲苯處理、石蠟包埋、切片,按常規操作進行蘇木精和伊紅染色,中性樹脂封片。光學顯微鏡下觀察睪丸形態和結構的變化,每張切片隨機觀察5個視野,并拍照取圖。
1.4.3Western blot檢測大鼠睪丸組織Beclin1、ATG5、ATG7 、LC3Ⅱ、Occludin和β-catenin蛋白的表達水平 精密稱取30~40 mg新鮮睪丸組織,按照1 ∶20(質量 ∶體積)加入含磷酸化蛋白酶抑制劑的RIPA裂解液,使用電動勻漿器低溫勻漿40 s后,于冰上裂解1 h,期間每10 min渦旋一次。隨后,4 ℃條件下12 000 r·min-1離心10 min,取上清,采用BCA蛋白定量試劑盒進行蛋白濃度測定。在上清中按照4 ∶1加入上樣緩沖液,95 ℃加熱10 min滅活。每孔等量蛋白上樣,SDS-PAGE凝膠電泳并恒流轉PVDF膜。5%脫脂牛奶封閉1 h后,一抗4 ℃孵育過夜。TBST洗3次,每次5 min。隨后,二抗室溫孵育1 h,最后ECL化學發光法顯影。各組實驗均重復3次。
1.4.4免疫熒光法觀察大鼠睪丸組織Beclin1、LC3和β-catenin蛋白的表達及定位 取睪丸石蠟切片置于60 ℃烘箱烤片至蠟熔,二甲苯脫蠟與梯度乙醇脫水脫蠟,用檸檬酸修復液高壓修復抗原10~15 min。待冷卻至室溫后,5% BSA封閉1 h,孵育一抗,4 ℃孵育15 h左右。二抗37 ℃孵育60~90 min后,DAPI染色10 min,最后抗熒光淬滅劑封片,4 ℃避光保存。在激光共聚焦顯微鏡下進行觀察,每張切片隨機觀察5個視野并拍照取圖。各組實驗各重復3次。
2 結果
2.1 增齡過程中大鼠睪丸組織形態學的變化HE染色顯示,6月齡組大鼠睪丸組織生精小管形態結構完整,管內可見排列整齊的各級生精細胞;12月齡時,大鼠睪丸組織中部分生精小管形態結構開始出現一定程度的退化;18月齡時,睪丸組織形態進一步變差,上皮變薄,生精細胞層數減少,小管界膜增厚;24月齡組大鼠睪丸部分生精小管發生嚴重萎縮,小管中生精細胞缺失,結果見Fig 1。
2.2 增齡過程中大鼠睪丸血睪屏障相關蛋白β-catenin和Occludin表達水平的變化采用免疫熒光和Western blot法檢測了血睪屏障相關蛋白β-catenin表達水平的變化,結果如Fig 2和Fig 3所示,增齡過程中,大鼠睪丸組織中β-catenin 蛋白表達逐漸降低,且主要分布于生精小管管腔周圍。此外,隨著年齡增加,大鼠睪丸血睪屏障另一重要細胞連接蛋白Occludin表達水平也逐漸下降。以上結果提示,大鼠睪丸血睪屏障完整性隨增齡而逐漸被破壞。
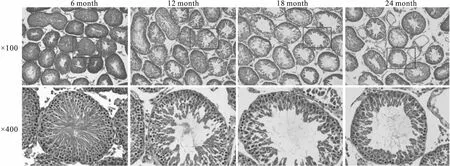
Fig 1 Morphological changes of testicular tissues in rats of different age groups
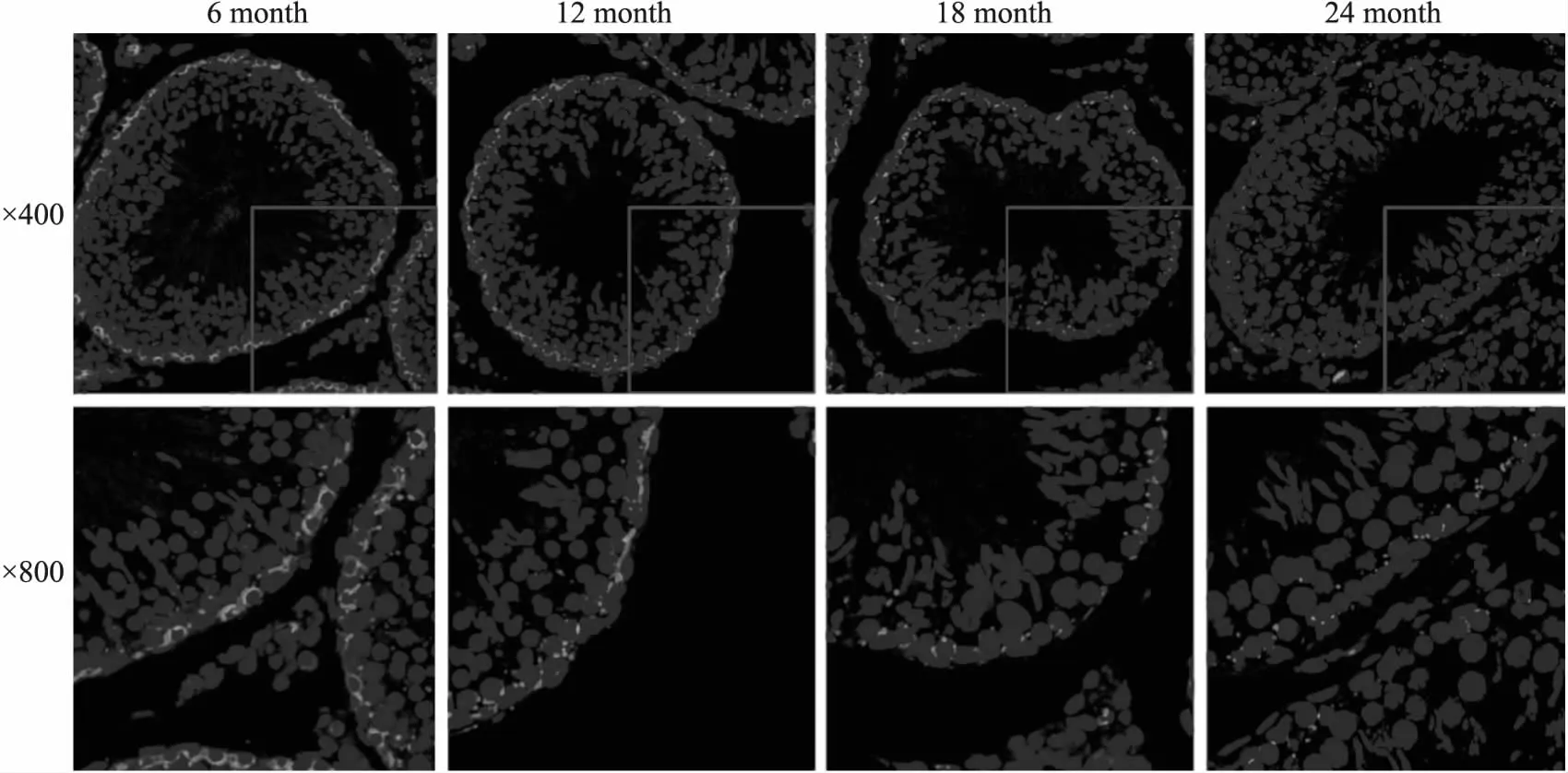
Fig 2 Expression and localization of β-catenin in testicular tissues in rats of different age groups
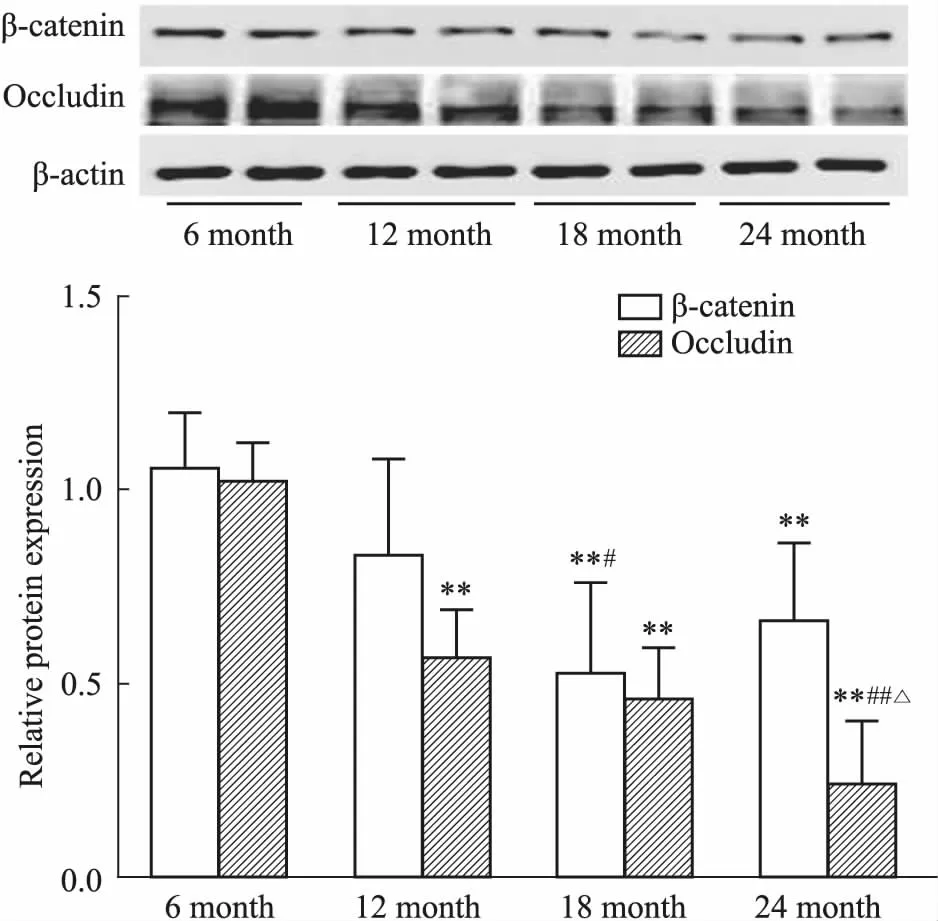
Fig 3 Expression levels of blood testis-associated protein β-catenin and Occludin in testicular tissues in rats of different age groups n=6)
2.3 增齡過程中大鼠睪丸自噬相關蛋白Beclin1、ATG5、ATG7和LC3Ⅱ表達水平的變化Beclin1、ATG5、ATG7和LC3Ⅱ是自噬過程中的幾個關鍵蛋白,可用于評價細胞內自噬功能的強弱。Western blot結果顯示,增齡過程中,大鼠睪丸組織中Beclin1、ATG5、ATG7和LC3Ⅱ蛋白表達水平逐漸下降,結果見Fig 4。
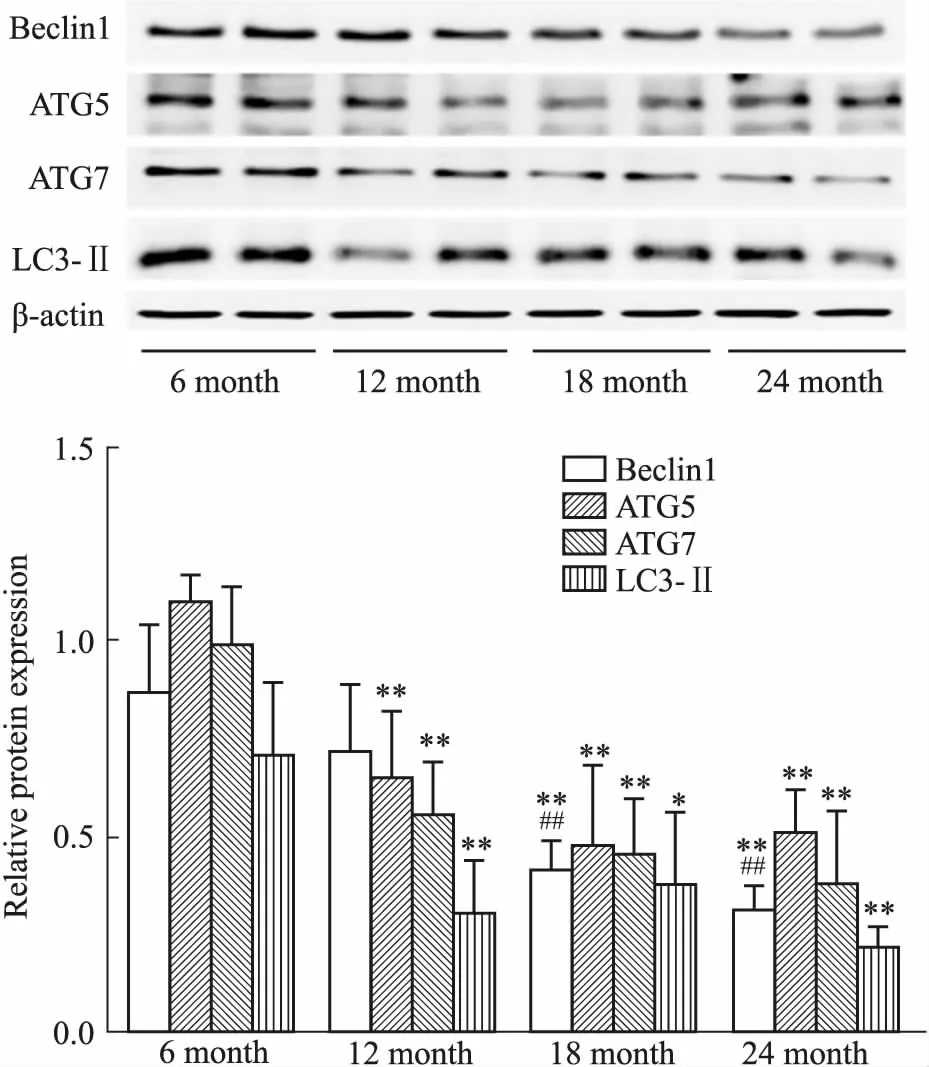
Fig 4 Changes of Beclin1, ATG5, ATG7 and LC3Ⅱ protein expression levels in testicular tissues in rats of different age groups n=6)
2.4 增齡過程中大鼠睪丸組織Beclin1和LC3蛋白表達及定位免疫熒光結果顯示,增齡過程中,大鼠睪丸組織中自噬標志蛋白Beclin1 和LC3 表達水平逐漸降低,且主要表達于睪丸生精小管上皮細胞中,結果見Fig 5。
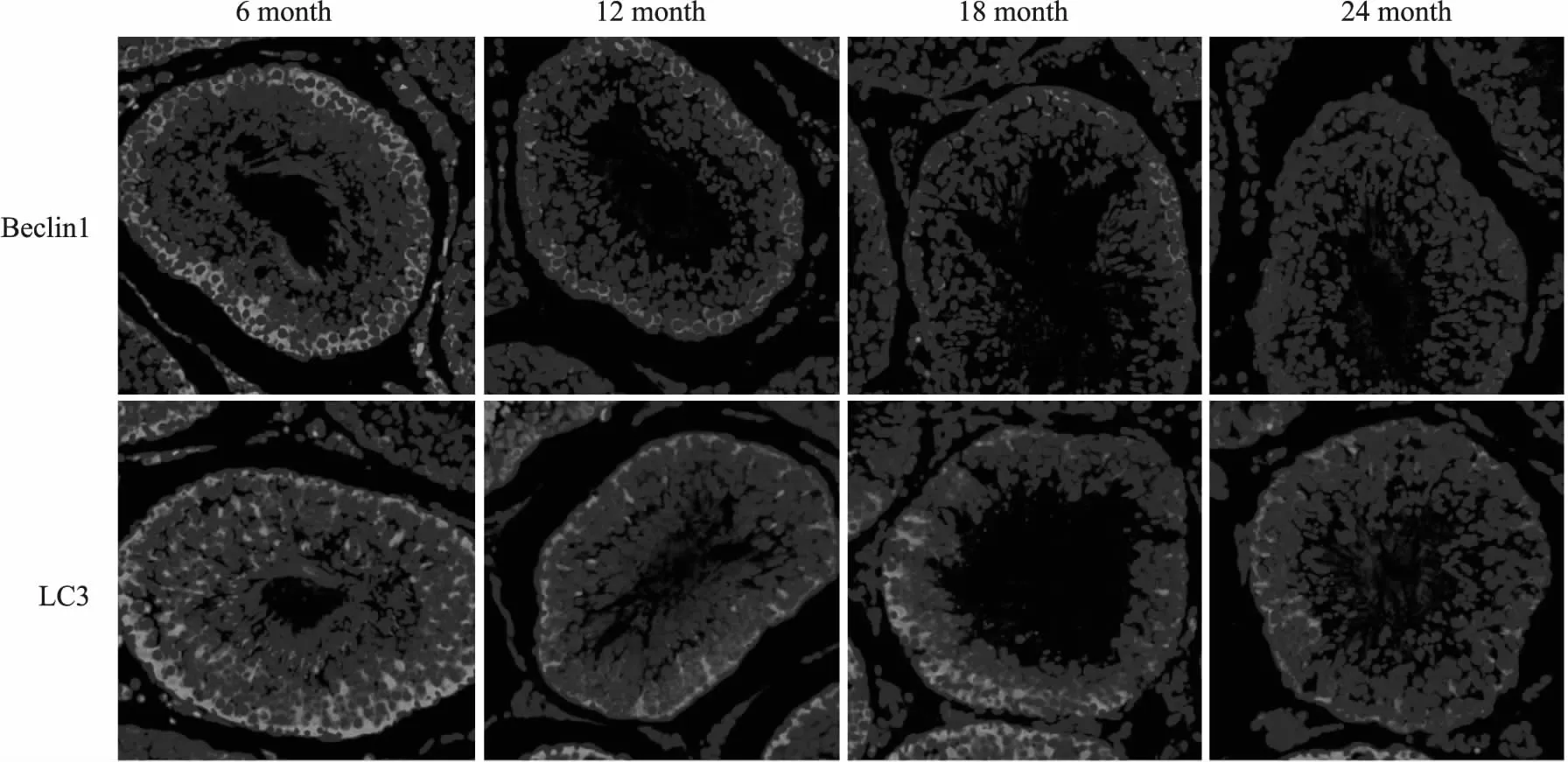
Fig 5 Expression and localization of Beclin1 and LC3 in testicular tissues in rats of different age groups (×400)
3 討論
衰老是器官功能和個體機能隨時間推移而發生退化的一個極其復雜和多因子的過程[4]。伴隨著機體老化、性腺結構和功能的退化是人類衰老的主要特征之一。睪丸是男性生殖系統的重要器官,具有合成、分泌雄激素和產生精子的能力。大量研究顯示[5-6],衰老睪丸生精小管形態結構發生明顯改變,如出現萎縮、生精細胞層數明顯減少和各級細胞排列疏松等。本研究也顯示,增齡過程中,大鼠睪丸組織形態逐漸發生改變,生精小管發生萎縮,生精細胞層數減少,細胞間間隙增大,且發生部分脫落現象,與文獻報道基本一致,提示增齡過程中大鼠睪丸生精功能逐漸發生減退。
最新研究顯示[1-2],血睪屏障的完整性被破壞是衰老睪丸生精功能減退的重要原因。Levy等[1]報道,老年大鼠睪丸血睪屏障相關超微結構發生明顯變化,血睪屏障被破壞,睪丸精子生成減少。Catriona等進一步發現,隨著大鼠年齡的增長,參與形成血睪屏障的相關細胞連接的蛋白表達減少,進而導致血睪屏障保護生殖細胞的能力下降[2]。睪丸中非信號的β-catenin作為黏附連接的adaptor蛋白,外接細胞連接的膜蛋白,內接細胞骨架纖維F-actin,是血睪屏障建立的一個不可缺少的連接蛋白[7]。β-catenin也是血睪屏障的特異性蛋白,有文獻也將其視為血睪屏障的蛋白標志物[8]。Occludin是生精上皮中支持細胞間形成緊密連接結構的基礎,其程序性的開啟與關閉保證了精子發生正常進行,若異常開放則會影響精子發生的正常進程。有文獻報道,干擾睪丸支持細胞內Occludin蛋白的功能狀態,會導致不育[9]。本研究顯示,增齡過程中大鼠睪丸中參與形成血睪屏障的相關細胞連接蛋白β-catenin和Occludin表達逐漸下降,提示增齡過程中大鼠睪丸血睪屏障的完整性逐漸被破壞,與現有文獻報道一致。
近期研究顯示,睪丸血睪屏障結構與功能的維持與睪丸中自噬水平密切相關。睪丸中的自噬對于維持精子發生的血睪屏障的形成以及某些成分的降解至關重要[10]。睪丸自噬功能受損可使構成血睪屏障的相關細胞連接蛋白(如Occludin)等下調,破壞血睪屏障的完整性,進而導致精子數量明顯減少,精子畸形率增加,生育能力明顯下降[3]。自噬過程中,LC3-Ⅱ和Beclin1 蛋白的表達強度與自噬活性密切相關,被認為是自噬體的標志分子[11]。ATG5 在自噬泡形成過程中起重要作用,可誘導自噬體的形成[12]。ATG7在自噬體和囊泡的形成過程中發揮重要作用,是激活細胞自噬必不可少的基因[13],且對于精子發生的正常進行至關重要[14]。有研究顯示,小鼠睪丸支持細胞中ATG5或ATG7被特異性敲除后,睪丸生精小管結構紊亂,血睪屏障完整性受損,精子頭部畸形且活力低下、小鼠不育[15]。本研究顯示,增齡過程中大鼠睪丸自噬相關蛋白Beclin1、ATG5、ATG7和LC3Ⅱ表達水平均出現下降,提示增齡過程中大鼠睪丸自噬水平逐漸下降。
綜上所述,增齡過程中大鼠睪丸生精功能減退,其機制與睪丸生精小管上皮細胞自噬水平下降,進而破壞血睪屏障的完整性,導致生精微環境改變有關。本實驗有助于更好地理解自噬在衰老睪丸生精功能衰退中的作用。
(致謝:本實驗完成于三峽大學醫學院國家中醫藥管理局中藥藥理科研三級實驗室。感謝醫學院實驗中心和腫瘤微環境與免疫治療湖北省重點實驗室對本課題提供的支持與幫助,感謝本課題組全體老師和同學對實驗的幫助!)
