結腸癌化療耐藥相關miRNA表達譜的初步研究*
2020-02-06 03:35:40張錦松陳兩洲
中國病理生理雜志 2020年1期
陳 敏,張錦松,徐 霖,陳兩洲,宋 學
(北京中醫藥大學廈門醫院普外科, 福建 廈門 361009)
隨著人們生活水平提高、生活方式和飲食結構的改變,結直腸癌的發病率在逐年上升。目前其發病率在惡性腫瘤中已上升至第3位[1]。在中國,結直腸癌發病率已經從70年代的12/千萬上升到56/千萬,年平均增長率為4.2%,顯著高于國際水平[2]。結直腸癌最有效的治療方法是根治性手術。但有些患者在手術前就已出現腫瘤遠處轉移或者在根治性手術之后出現腫瘤復發。對于這些患者,化療可以抑制腫瘤進一步生長,從而延長患者生存時間。然而,化療耐藥仍是橫亙于前的難題。有些類型的腫瘤細胞天生對某些化療藥耐藥,而且絕大多數癌細胞在化療過程中會逐漸形成化療耐藥,從而影響化療療效。
微小RNA(microRNA,miRNA,miR)是內源性的非編碼RNA,大小約20~25個核苷酸,通過結合在靶基因的3′末端來降解靶mRNA或阻遏靶mRNA的翻譯。miRNAs參與細胞增殖、分化、血管生成和凋亡等多種生物過程,其調節失衡與多種疾病有關,包括結直腸癌。異常的miRNAs表達與結直腸癌的發病和進展密切相關[3]。在結直腸癌發生和發展過程中, miRNAs可起到原癌基因或抑癌基因的作用。miRNAs與結直腸癌的化療耐藥也是息息相關。通過對miRNAs功能的識別,有望發現新的治療方法來逆轉化療耐藥。但是miRNAs在化療耐藥中的機制十分復雜。因此, 在整體上了解miRNAs調節腫瘤對化療耐藥的作用機制至關重要。應用基因芯片技術從整體上闡述miRNAs與結直腸癌化療耐藥相關性的文章較少,因此本研究利用基因芯片技術, 篩選人結腸癌細胞系及其耐長春新堿細胞系表達有明顯差異的miRNAs,并用RT-qPCR技術驗證結果;預測可能受其調控的基因,并對這些基因進行生物信息學分析;從整體上對miRNAs與結腸癌化療耐藥的關系進行了詳細的闡述,為研究miRNAs在化療耐藥中的作用提供線索。
材 料 和 方 法
1 細胞株
人結腸癌細胞系HCT8及其耐長春新堿細胞系HCT8/v購自上海奧陸生物有限公司。
2 主要試劑
長春新堿(vincristine,VCR)和生理鹽水購自上海阿拉丁生化科技股份有限公司;MTT購自Sigma-Aldrich;PBS、DMEM培養基、0.25%胰酶和胎牛血清(fetal bovine serum,FBS)購自Life Tech。所用的芯片為Agilent miRNA芯片,由上海康成生物技術有限公司提供技術支持。所用引物由上海康成生物技術有限公司根據設計合成,序列見表1。

表1 RT-qPCR使用引物列表Table 1. The primer list using in RT-qPCR
F: forward; R: reverse; GSP: gene-specific primer.
3 主要方法
3.1細胞培養及耐藥性檢測 實驗細胞于DMEM培養液中,在37℃、5%CO2的細胞培養箱中培養。MTT法驗證細胞的耐藥性。
3.2RNA的提取和質量控制 按照TRIzol方法抽提總RNA。應用NanoDrop 1000評估RNA完整性和質量。變性瓊脂糖凝膠電泳測定RNA的完整性及確定不存在污染。樣品A260/A280的比率接近 2.0(1.8~2.1),A260/A230的比率大于1.8。電泳RNA的28S和18S條帶光滑,亮度強。上條帶的強度是下條帶的2倍左右。在28S條帶上方無彌散帶或條出現。
3.3標記、雜交、清洗和掃描 采用Agilent的miRNA Complete Labeling and Hyb Kit進行標記。將500 ng的RNA添加到1.5 mL微離心管中。加入2 μL的CIP Master Mix,并在37 ℃下孵育反應30 min。樣品中加入2.8 μL的DMSO,并在100 ℃下孵育5至10 min。立即轉移到冰水浴。加入4.5 μL的Ligation Master Mix,并在16 ℃孵育2 h。使用帶加熱器的真空集中器加熱到45~55 ℃,完全干燥樣本。在18 μL無核酸酶水中重新懸浮干燥的樣品。加入4.5 μL的10× Gene Expression Blocking Agent和2×Hi-RPM Hybridization Buffer。在100 ℃孵育5 min,并立即轉移到冰浴5 min。將干凈的墊片裝入Agilent SureHyb室底座,標簽朝上,與室底座的矩形部分對齊。墊片沾濕,并評估氣泡的流動性。將組裝的室置于設置為55 ℃的雜交爐中。雜交旋轉器的轉速設置為在20 r/min,在55 ℃下雜交20 h。隨后按程序完成清洗,將片放在片支架中。芯片經過洗滌后,使用Agilent Microarray Scanner (Agilent p/n G2505C)掃描。
3.4芯片數據分析 使用Agilent Feature Extraction 11.0.1.1軟件獲得芯片圖,并讀值,得到原始數據。使用GeneSpring GX 14.9軟件(Agilent Technologies)對原始數據進行Quantile標準化和隨后的數據處理。原始數據標準化后經過篩選高質量探針(某探針在6個樣品中至少有3個被標記為detected)進行進一步分析。兩組樣品間具有統計學意義的差異表達miRNAs通過火山圖篩選。兩個樣品間差異表達miRNAs通過Fold Change篩選。使用R腳本進行層次聚類。
3.5RT-qPCR驗證 TRIzol法提取樣品的總RNA并進行質量檢測。檢測合格后,使用RNA進行cDNA合成。合成的cDNA用于RT-qPCR。各樣品的目的miRNA和內參照(U6)分別進行real-time PCR反應。數據采用2-ΔΔCt法進行分析。引物序列見表3。
3.6靶基因預測和生物信息學分析 選取部分有顯著差異表達的miRNAs進行靶基因預測和生物信息學分析。應用TargetScan 7.1和miRDB 5進行靶基因預測,對2個數據庫預測的基因取交集。利用GO和KEGG數據庫對交集內的靶基因進行生物信息學分析。
結 果
1 HCT8/v對長春新堿耐藥性的檢測
各種濃度的長春新堿均對HCT8/v細胞無抑制作用,結果如圖1所示。

Figure 1. The viability of HCT8/v cells treated with vincristine (VCR) at different concentrations. Mean±SD.n=3.
圖1 HCT8/v細胞對長春新堿耐藥性的測定
2 細胞中RNA純化和質檢
人結腸癌細胞系HCT8及其耐長春新堿細胞系HCT8/v的總RNA經NanoDrop 1000檢測A260/A280的比率均介于1.8~2.1之間,A260/A230的比率均大于1.8。經瓊脂糖凝膠電泳鑒定28S和18S兩條帶清晰,沒有拖尾現象,提示RNA的質量合格。結果如圖2所示。
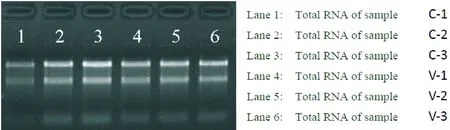
Figure 2. Results of denatured agarose gel electrophoresis. C=HCT8; V=HCT8/v.
圖2 變性瓊脂糖凝膠電泳結果
3 芯片掃描結果
HCT8/v和HCT8細胞的miRNA經基因芯片檢測比較, 改變2倍以上的miRNA數目為342個, 其中基因表達上調190個, 基因表達下調152個,見圖3。部分差異表達的基因列表見表2、3。
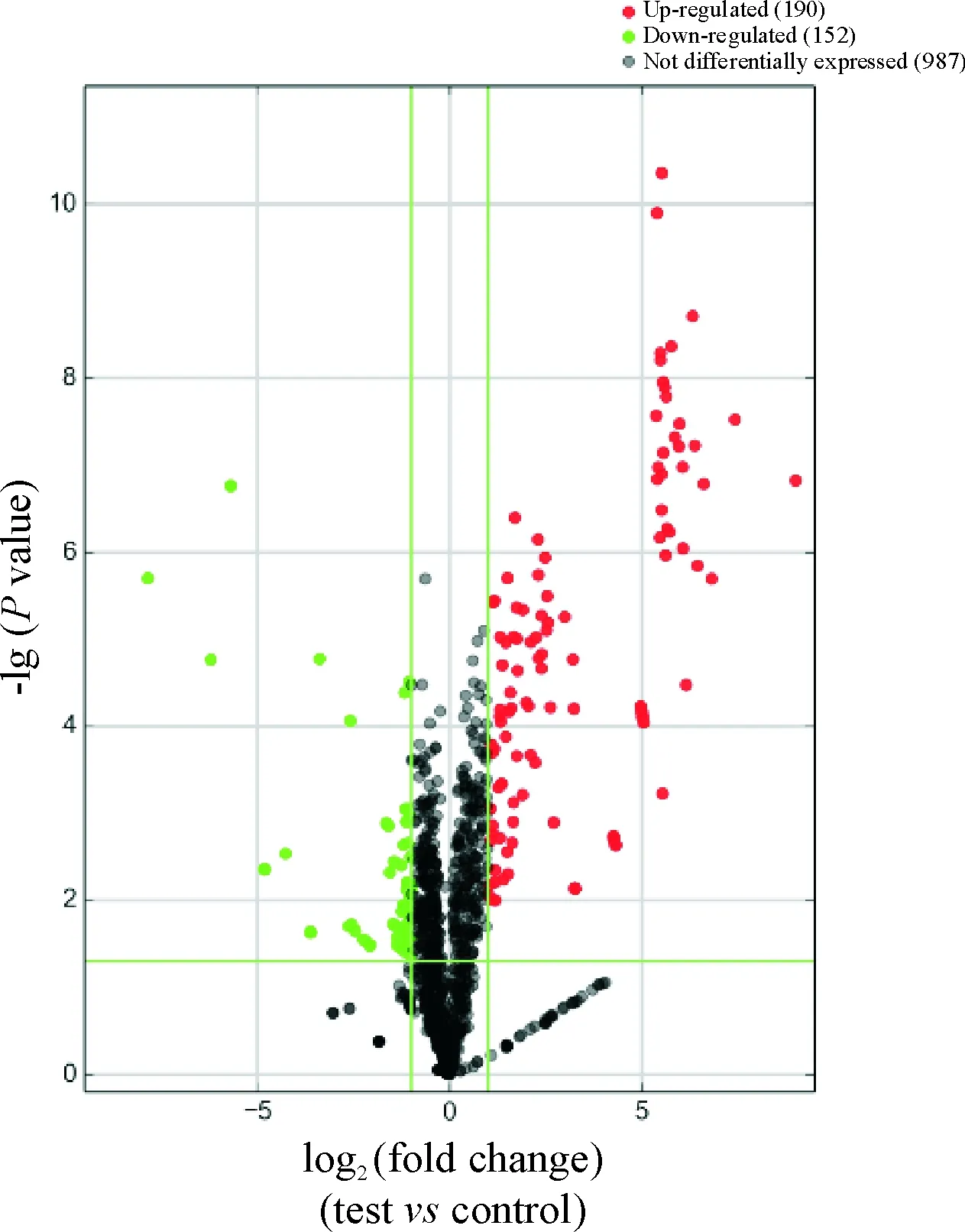
Figure 3. Volcanic map of differentially expressed miRNAs between HCT8 group and HCT8/v group. Green dot represents a cut of ≥2 times; red dot represents an increase of ≥2 times; green horizontal line above representsP≤0.05, green horizontal line below representsP≥0.05.
圖3 HCT8組和HCT8/v組間表達差異miRNAs的火山圖
4 RT-qPCR驗證芯片結果
挑選5個有顯著差異表達的miRNAs進行RT-qPCR驗證,其中miR-125-5p、miR-181c-5p和miR-153-3的表達情況和芯片檢測結果一致,miR-130a-3p和miR-149-3p的表達與芯片檢測結果不一致,見圖4。
5 靶基因預測
對上述部分miRNAs的靶基因進行預測,TargetScan 7.1預測到4 150個靶基因,miRDB 5預測到3 311個靶基因,2個數據庫共同預測到的基因有1 799個。
5 KEGG通路富集分析
KEGG分析結果顯示,這些miRNAs的靶基因主要涉及94個信號通路:其中最為富集的是軸突導向(axon guidance)通路,有39個基因的表達發生變化;其次分別是胰島素信號通路(insulin signaling pathway)及磷脂酶D信號通路(phospholipase D signaling pathway),分別有31及32個基因發生變化;其余變化顯著的依次為腎細胞癌(renal cell carcinoma)、動物自噬(autophagy-animal)、FoxO信號通路(FoxO signaling pathway)、ErbB信號通路(ErbB signaling pathway)、結直腸癌(colorectal cancer)、mTOR信號通路(mTOR signaling pathway)和肝細胞癌(hepatocellular carcinoma)。
6 GO通路富集分析
生物過程(biological processes,BP)分析結果顯示,這些miRNAs的靶基因主要涉及1 446個生物學過程:其中最為富集的是正向調節細胞過程(positive regulation of cellular process),有604個基因的表達發生變化;其次分別是正向調節生物過程(positive re-gulation of biological process)及正向調節代謝過程(positive regulation of metabolic process),分別有654及415個基因發生變化;其余變化顯著的依次為正向調節細胞代謝過程(positive regulation of cellular metabolic process)、神經系統發育(nervous system development)、正向調節大分子代謝過程(positive regulation of macromolecule metabolic process)、正向調節氮化合物代謝過程(positive regulation of nitrogen compound metabolic process)、負向調節細胞過程(negative regulation of cellular process)、正向調節基因表達(positive regulation of gene expression)和調節細胞代謝過程(regulation of cellular metabolic process)。

表2 下調≥2倍的miRNAs (部分)Table 2. The miRNAs down-regulated by ≥2 times (part)
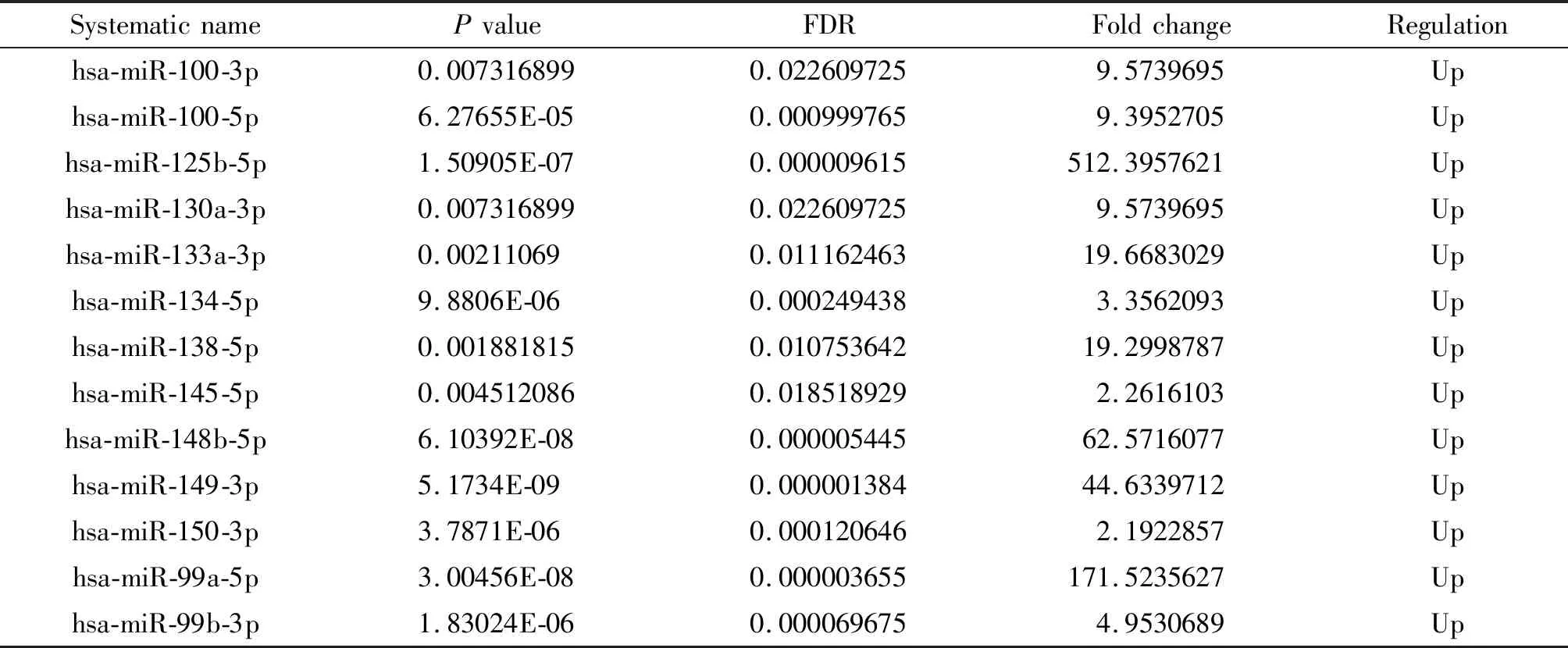
表3 上調≥2倍的miRNAs (部分)Table 3. The miRNAs down-regulated by ≥2 times (part)

Figure 4. The results of RT-qPCR.
圖4 RT-qPCR實驗結果
細胞組分(cell component,CC)分析結果顯示,這些miRNAs的靶基因主要涉及268個細胞組分:其中最為富集的是細胞內(intracellular),有1 351個基因的表達發生變化;其次分別是細胞內部分(intracellular part)及細胞內細胞器(intracellular organelle),分別有1 331及1 184個基因發生變化;其余變化顯著的依次為細胞器(organelle)、細胞質(cytoplasm)、膜旁細胞器(membrane-bound organelles)、細胞內有界細胞器(intracellular membrane-bound organelles )、神經元部分(neuron part)、細胞內細胞器部分(intracellular organelle part)和突觸小泡(synaptic vesicle)。
分子功能(molecular function,MF)分析結果顯示,這些miRNAs的靶基因主要涉及209個分子功能:其中最為富集的是蛋白結合(protein binding),有1 120個基因的表達發生變化;其次分別是酶結合(enzyme binding)及結合(binding),分別有281及1 330個基因發生變化;其余變化顯著的依次為調節區域核酸結合(regulatory region nucleic acid binding)、RNA聚合酶II調控區DNA結合(RNA polymerase II regulatory region DNA binding)、轉錄調控區DNA結合(transcription regulatory region DNA binding)、序列特異性雙鏈DNA結合(sequence-specific double stranded DNA binding)、RNA聚合酶II調控區序列特異性DNA結合(RNA polymerase II regulatory region sequence-specific DNA binding)、轉錄調控區序列特異性DNA結合(transcription regulatory region sequence-specific DNA binding)、RNA聚合酶II轉錄因子活性及序列特異性DNA結合(RNA polymerase II transcription factor activity, sequence-specific DNA binding)。
讀者可向作者索取KEGG分析和GO分析的詳細圖則和相關資料。
討 論
化療耐藥是導致化療失敗的主要原因。如何逆轉化療耐藥是世界難題。miRNAs與腸癌細胞化療耐藥的關系十分密切[4-5]。對于腸癌化療耐藥,從miRNAs入手研究也許能開辟一條新的道路。
我們通過基因芯片和RT-qPCR首次證明miR-125-5p、miR-181c-5p和miR-153-3p與結腸癌化療耐藥相關,這些miRNAs在其他惡性腫瘤化療耐藥的過程中也起重要作用[6-7],其調節結腸癌化療耐藥的機制有待進一步研究。
GO主要從功能、參與的生物途徑及細胞中的定位對基因進行分析。通過GO富集分析可以了解這些靶基因富集在哪些生物學功能上、哪些生化途徑中、哪些細胞結構上。信號通路指的是多個蛋白質間相互作用來調節細胞的功能和代謝的過程。KEGG分析結果可見,靶基因最為富集的是軸突導向通路,有39個基因富集。軸突導向通路主要在重新排列生長錐和軸突的局部細胞骨架和質膜中發揮作用,其也可以通過局部翻譯和轉錄來控制基因的表達,與細胞遷移相關[8]。有學者一次性研究并確定這條通路上的16個基因表達異常與胰腺癌相關[9],而研究這條通路上單個基因表達異常與腫瘤關系的文章更是不勝枚舉。從GO的有向無環圖的結果來看,在分子功能方面其顯著富集的最基本旁路是RNA聚合酶II調控區序列特異性DNA結合旁路,有98個基因富集。這條旁路的蛋白能有選擇性地和非共價地和DNA的特定序列(一個調控區域,能通過RNA聚合酶II控制一個基因或順反子轉錄)相互作用。在生物過程方面:其顯著富集的最基本的過程是:(1)正向調節基因表達(positive regulation of gene expression):即增加基因表達的頻率、速率或程度的生物過程。基因表達是個過程,在這個過程里,基因的編碼序列轉化成一個成熟的基因產物(蛋白或RNA)。包括RNA轉錄子和成熟的RNA產物(mRNA或circRNA)生產過程,以及mRNA或circRNA轉化成蛋白的過程。也包括蛋白的成熟過程(當需要從非活性前體形式變成活性形式時);(2)正向調節細胞代謝過程(positive regulation of cellular metabolic process),即單個細胞通過激活或增加化學反應和信號旁路的頻率、速率或程度來改變化學物質的生物過程;(3)神經系統發育(nervous system development),即神經組織從形成到成熟的生物過程。在細胞組分方面,其顯著富集的最基礎結構是細胞內細胞器組成部分(即細胞核、線粒體、質體、液泡、囊泡、核糖體和細胞骨架等的組成部分,但不包括質膜)和細胞內有界細胞器(即在細胞內的,有單或雙層膜的,具有獨特形態和功能的結構,包括細胞核、線粒體、質體、液泡和囊泡,不包括質膜)。從GO的解析來看,耐藥相關的基因主要富集的旁路是RNA聚合酶II調控區序列特異性DNA結合旁路,主要通過正向調節發揮作用,位置主要是在細胞內有界細胞器上。
本研究利用芯片技術確定了化療耐藥相關的miRNAs,并通過RT-qPCR首次驗證了miR-125-5p、miR-181c-5p和miR-153-3p在化療耐藥細胞中的異常表達。我們選取部分表達差異顯著的miRNAs進行靶基因預測,并進行生物信息學分析,進一步確定了與化療耐藥密切相關的基因所在的細胞位置,涉及的生物過程,所用的信息通道,為進一步研究結腸癌化療耐藥提供了堅實的分子基礎。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
科學大眾(2020年12期)2020-08-13 03:22:22
航空世界(2020年10期)2020-01-19 14:36:20
