IFN-γ在早孕母胎界面對吲哚胺2,3-雙加氧酶(IDO)表達(dá)的正性調(diào)節(jié)研究
2020-02-20 10:54:26涂媛媛喻海芬趙淑云翁宇紅李仕祥錢志瑤黃官友
中國免疫學(xué)雜志 2020年2期
關(guān)鍵詞:界面
涂媛媛 喻海芬 趙淑云 翁宇紅 王 珺 李仕祥 錢志瑤 黃官友
(貴州醫(yī)科大學(xué)附屬醫(yī)院,貴陽 550004)
胎兒如何逃脫母體免疫系統(tǒng)的攻擊,具體機(jī)制尚不明確。母胎界面由母體的蛻膜組織及胚胎的滋養(yǎng)細(xì)胞組成,胚胎作為半同種移植物不被母體排斥,與母胎界面的免疫耐受狀態(tài)相關(guān)。干擾素(interferon-γ,IFN-γ)由分泌蛋白質(zhì)組成,存在于各種脊椎動物體內(nèi),具有抗病毒、抗細(xì)胞增殖、抗腫瘤、免疫調(diào)節(jié)等多種活性[1]。吲哚胺-2,3雙加氧酶(indoleamine-2,3-dioxygenase,IDO)是沿犬尿酸途徑中分解代謝催化色氨酸的限速酶,存在于哺乳動物的組織與細(xì)胞中,在淋巴組織和胎盤中廣泛表達(dá),正常狀態(tài)下呈低水平表達(dá),在炎癥過程或感染中,IDO表達(dá)顯著增加,另外IFN-γ和脂多糖(lipopolysacch-arides,LPS)也可誘導(dǎo)IDO表達(dá)[2,3]。雌激素是一種性激素,對生殖發(fā)育至關(guān)重要,主要有三種類型:雌酮、雌二醇、雌三醇。當(dāng)雌二醇水平逐漸升高約100倍時,雌三醇水平從早期妊娠到晚期妊娠增加約1 000倍,產(chǎn)后則急劇下降[4]。在懷孕期間,胎盤細(xì)胞產(chǎn)生性激素,特別是雌激素。雌激素在免疫系統(tǒng)中也有作用,但其機(jī)制尚不完全清楚。雌激素主要通過影響胸腺的T淋巴細(xì)胞、B淋巴細(xì)胞及細(xì)胞因子表達(dá)而調(diào)節(jié)免疫系統(tǒng),此外雌激素還可促使自身免疫性B細(xì)胞增多而克服免疫耐受,通過改變細(xì)胞因子來誘導(dǎo)B細(xì)胞活化,降低B細(xì)胞凋亡。由于Th1型細(xì)胞因子參與急性同種移植排斥反應(yīng),Th2型細(xì)胞因子參與免疫耐受。傳統(tǒng)理論認(rèn)為母胎界面中Th1/Th2的平衡向Th2偏倚有利于母胎免疫耐受,向Th1偏倚與復(fù)發(fā)性流產(chǎn)等密切相關(guān)。近年的研究表明,胚胎植入部位有包括IFN-γ在內(nèi)的高水平的Th1型細(xì)胞因子[5,6],而妊娠期有一個非常顯著的特征是胎盤組織通過產(chǎn)生大量的IDO來維持母胎耐受。在免疫微環(huán)境中,早期IDO的表達(dá)可能與IFN-γ有關(guān),而晚期可能與TGF-β有關(guān)[7]。由于Th1型細(xì)胞因子參與急性同種移植排斥反應(yīng),Th2型細(xì)胞因子參與免疫耐受。傳統(tǒng)理論認(rèn)為母胎界面中Th1/Th2的平衡向Th2偏倚有利于母胎免疫耐受,向Th1偏倚與復(fù)發(fā)性流產(chǎn)等密切相關(guān)[8-13]。本文分析早孕絨毛和脫膜組織中IFN-γ上調(diào)IDO表達(dá)的可能性。
1 材料與方法
1.1材料
1.1.1標(biāo)本 選擇2015年10月至2017年6月,在貴州醫(yī)科大學(xué)附屬醫(yī)院收治的接受人工流產(chǎn)治療的孕6~9周正常妊娠婦女38例及人工流產(chǎn)獲得的絨毛及蛻膜組織。納入標(biāo)準(zhǔn):健康正常妊娠婦女;妊娠6~9周;妊娠期間未接受過藥物治療。排除標(biāo)準(zhǔn):合并全身性疾病者;接受稽留流產(chǎn)或孕酮和米非司酮人流前處理者。研究經(jīng)貴州醫(yī)科大學(xué)附屬醫(yī)院倫理委員會批準(zhǔn),且患者均簽署知情同意書。
1.1.2主要試劑 細(xì)胞裂解緩沖液(RIPA)(BL502A)和蛋白酶抑制劑(P8340)均購自美國Sigma公司;一抗和二抗去除液(HCY055)、三羥甲基氨基甲烷-甘氨酸電泳液粉劑(56-40-6)及BCA-100蛋白定量試劑盒(P0012)均購自碧云天公司(877-616-CELL2355)、(D3H2)XP?Rabbit mAb、10×TBST(T1081)、IFN-γ(877-616-cell 2355)和兔抗IDO抗體均(86630)購自美國Cell Signaling Technology公司;PVDF膜(IPVH00010PCRE)、ECL試劑(c_311032402_10)和HRP標(biāo)記山羊抗兔IgG(M_25420100_10)均購自PerkinElmer公司;預(yù)染蛋白相對分子質(zhì)量Marker(BL706A)和5×SDS-PACE蛋白上樣緩沖液(BL502A)均購自美國Biosharp公司;兔抗GAPDH抗體(A01020美國Gnne Tex公司);SDS-PACE制備試劑盒(325OGR500)(北京索萊寶科技有限公司)。
1.2方法
1.2.1IFN-γ和IDO在早孕絨毛及蛻膜中的表達(dá) 用磷酸緩沖鹽溶液(phosphate buffer saline,PBS)洗滌早孕絨毛和蛻膜殘留的紅細(xì)胞,取絨毛和蛻膜組織各100 mg,置于研缽并加入適量液氮,碾碎組織成粉末,加入蛋白酶抑制劑的裂解液充分裂解,置于4℃離心機(jī),3 000 r/min離心30 min,取上清并采用BCA檢測蛋白濃度。加緩沖液煮沸變性,將變性的樣本加入聚丙烯酰胺凝膠中電泳,轉(zhuǎn)聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF),封閉,孵育一抗,4℃過夜,洗膜加二抗,孵育1 h,ECL顯影,圖片瀏覽軟件(Imagel J)處理圖片,甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)為內(nèi)參,IFN-γ和IDO的表達(dá)采用目的蛋白條帶與內(nèi)參吸光度值的比值表示(標(biāo)準(zhǔn):Western blot實驗得到的目的蛋白灰度值,除以GAPDH內(nèi)參灰度值得到的目的蛋白的相對表達(dá)量)。
1.2.2Western blot檢測不同濃度IFN-γ培養(yǎng)對IDO蛋白表達(dá)的影響 無菌收集絨毛和蛻膜組織后,立即放入0.1 m無菌磷酸鹽緩沖液(phosphate buffer saline pH7.2,PBS pH7.2)中且15 min內(nèi)在盒內(nèi)運送到實驗室,然后用PBS洗滌去除紅細(xì)胞。在Western blot分析前,將組織標(biāo)本保存在-80℃處,或切成小塊(濕重≤1 mg)進(jìn)行培養(yǎng)。培養(yǎng)樣品在收集后40 min內(nèi)進(jìn)行處理。將小塊絨毛和蛻膜用F-12/DMEM(21041025加拿大Invitrogen公司)沖洗2次,離心5 min后,在培養(yǎng)基中加入10%FBS(10100147吉布科;澳大利亞)和1%青霉素,在37℃溫箱(美國Thermo公司)與5% CO2共同孵育。給予不同濃度的IFN-γ(1、10、100 ng/ml)分別以48 h、72 h培養(yǎng)胚胎絨毛組織與蛻膜組織,對照組用PBS水替代,Western blot檢測IDO在兩種組織中的表達(dá)情況。
1.3統(tǒng)計學(xué)方法 采用SPSS22.0軟件對實驗數(shù)據(jù)進(jìn)行統(tǒng)計,t檢驗和雙變量的直線相關(guān)性分析IFN-γ和IDO在早孕絨毛及蛻膜組織中的表達(dá)等計量資料,P<0.05為差異有統(tǒng)計學(xué)意義。
2 結(jié)果
2.1IFN-γ、IDO在早孕絨毛及蛻膜組織中的表達(dá)及相關(guān)性 Western blot檢測顯示早孕絨毛組織和蛻膜組織中(圖1A)均可見分子量為46 kD和17 kD的蛋白條帶,與人的IDO和IFN-γ蛋白分子量大小一致,提示絨毛和蛻膜中均有IDO和IFN-γ的表達(dá)。分別對IDO、IFN-γ在兩種組織中的表達(dá)情況進(jìn)行統(tǒng)計分析,結(jié)果顯示,IDO與IFN-γ蛋白在絨毛組織中呈正相關(guān)表達(dá),如圖1B所示(r=0.987 1,P<0.05);IDO與IFN-γ蛋白在脫膜組織中呈正相關(guān)表達(dá),如圖1C所示(r=0.979 1,P<0.05)。分別以IDO、IFN-γ在絨毛組織與蛻膜組織中的表達(dá)量進(jìn)行統(tǒng)計分析,如圖2,結(jié)果顯示,IDO在蛻膜組織中的表達(dá)高于在絨毛組織中的表達(dá)(P=0.002);IFN-γ在蛻膜組織中的表達(dá)同樣高于在絨毛組織中的表達(dá)(P=0.001)。圖3為各數(shù)據(jù)匯總統(tǒng)計分析圖。
2.2IFN-γ誘導(dǎo)絨毛和蛻膜中IDO的表達(dá) 通過IFN-γ誘導(dǎo)絨毛和蛻膜中IDO的表達(dá),以確定IFN-γ是否能影響IDO的表達(dá)。將絨毛和蛻膜分別切成小塊,孵育6 h,然后加入不同濃度(1、10、100 ng/ml)的IFN-γ,培養(yǎng)48 h和72 h后分別進(jìn)行Westren blot分析,以檢測IDO的表達(dá)。根據(jù)IDO、IFN-γ擴(kuò)增帶灰度值與GAPDH帶灰度值的比值進(jìn)行統(tǒng)計分析,數(shù)據(jù)為平均值±Se,相關(guān)實驗結(jié)果重復(fù)3次后選取1例為文章示例圖。圖2、3表示在IFN-γ濃度分別為1、10、100 ng/ml時,對絨毛組織孵育48、72 h時, IDO表達(dá)情況與不同IFN-γ濃度之間的關(guān)系,P值分別為0.048、0.036,r值分別為0.952 1,0.964 1;圖4、5為IFN-γ濃度分別為1、10、100 ng/ml時,對蛻膜組織孵育48、72 h時,IDO表達(dá)情況與不同IFN-γ濃度之間的關(guān)系,P值分別為0.036、0.034,r值分別為0.964 1、0.966 4。結(jié)果顯示,在培養(yǎng)的絨毛和蛻膜組織中加入IFN-γ培養(yǎng)48 h及72 h后,IDO蛋白有較高表達(dá),均高于對照組,且隨著IFN-γ濃度的升高而增加(圖2~5)。由此可知,IFN-γ可上調(diào)母胎界面中IDO的表達(dá)來維持免疫耐受。

圖1 IDO與IFN-γ在絨毛、蛻膜組織中的表達(dá)Fig.1 IDO and IFN-γ expression in chorionic villi and deciduaNote:Samples of Numbers 1-8.***.P<0.001,**.P<0.01,*.P<0.05.
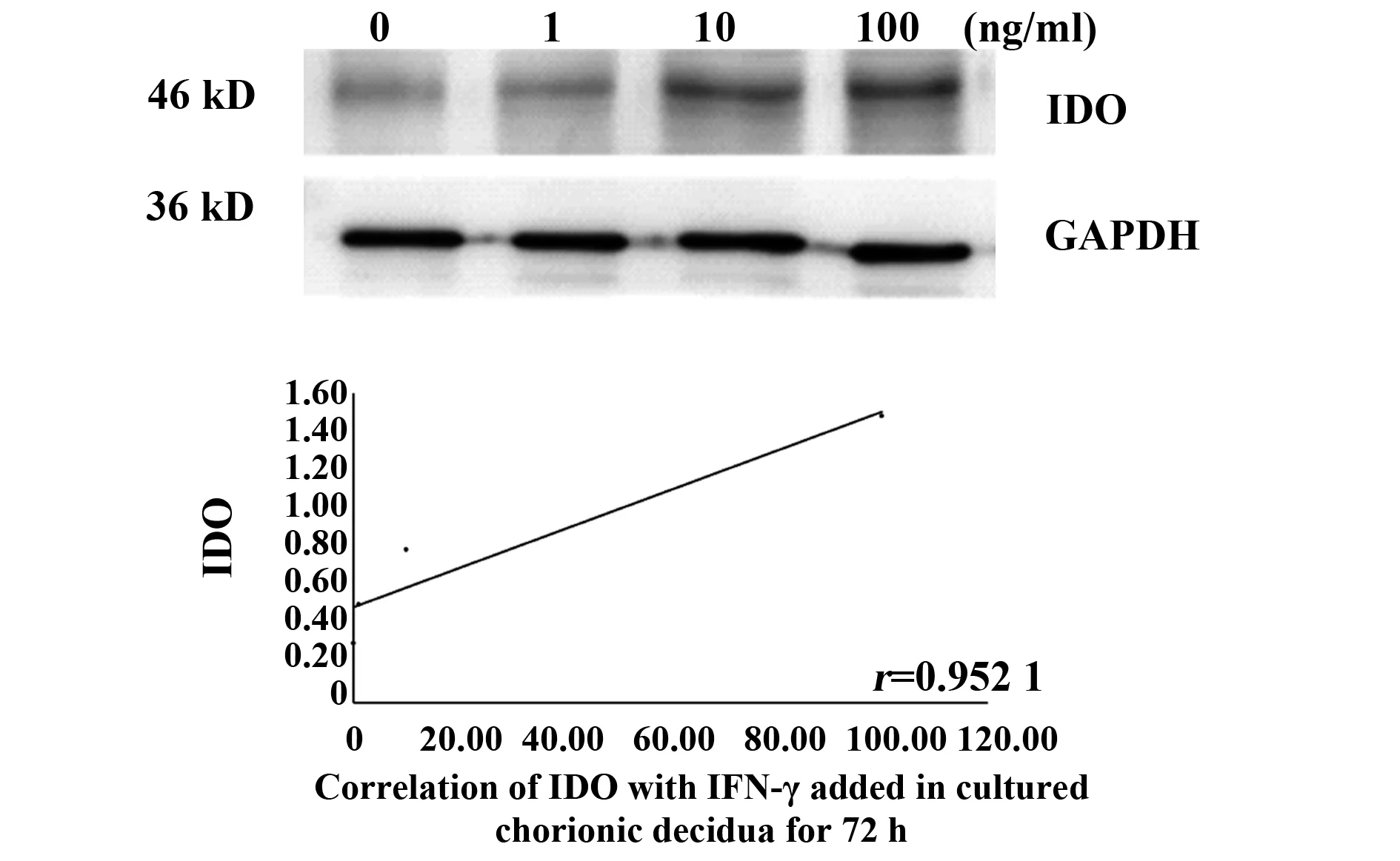
圖2 絨毛組織中不同IFN-γ濃度48 h IDO的表達(dá)情況Fig.2 Expression of IDO with IFN-γ added in cultured chorionic villi for 48 h
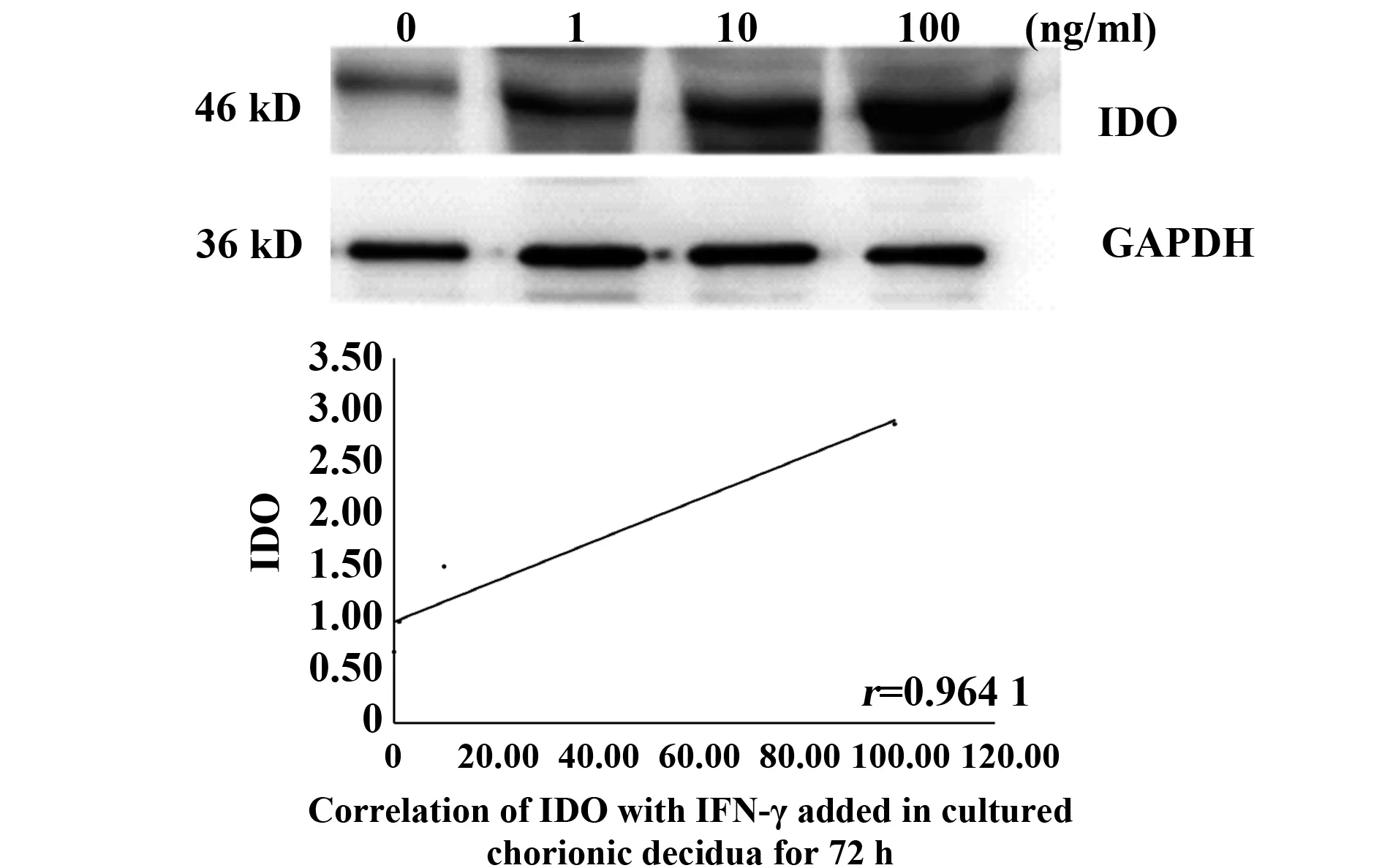
圖3 絨毛組織中不同IFN-γ濃度72 h IDO的表達(dá)情況Fig.3 Expression of IDO with IFN-γ added in cultured chorionic villi for 72 h

圖4 蛻膜組織中不同IFN-γ濃度48 h IDO的表達(dá)情況Fig.4 Expression of IDO with IFN-γ added in cultured decidua for 48 h

圖5 蛻膜組織中不同IFN-γ濃度72 h IDO的表達(dá)情況Fig.5 Expression of IDO with IFN-γ added in cultured decidua for 72 h
3 討論
本研究結(jié)果表明,在早孕絨毛和蛻膜組織中,IFN-γ和IDO的表達(dá)均呈正相關(guān),提示在該組織中,IFN-γ上調(diào)IDO的可能性較大; 在培養(yǎng)的早孕絨毛和蛻膜組織中加入不同濃度的IFN-γ,發(fā)現(xiàn)所加入的IFN-γ的濃度與IDO的表達(dá)均呈正相關(guān),揭示在早孕絨毛和蛻膜組織中,IFN-γ上調(diào)IDO的表達(dá)。Kudo等[14]曾報道在培養(yǎng)的人胎盤絨毛組織中加入不同濃度的IFN-γ,發(fā)現(xiàn)所加入的IFN-γ的濃度與IDO mRNA的表達(dá)呈劑量依賴關(guān)系,但該文的研究僅限于mRNA水平,且僅限于細(xì)胞培養(yǎng)(只研究培養(yǎng)的胎盤絨毛,未研究蛻膜組織),并沒有報道未培養(yǎng)的早孕絨毛和蛻膜組織中IFN-γ和IDO的表達(dá)的關(guān)系。我們在Kudo 等報道的基礎(chǔ)上做了進(jìn)一步的研究。本研究結(jié)果表明,不僅僅是絨毛組織,蛻膜組織亦可通過IFN-γ上調(diào)IDO的表達(dá)來致免疫耐受。
IFN-γ誘導(dǎo)的一氧化氮敏感性效應(yīng)機(jī)制有可能是誘導(dǎo)IDO的作用機(jī)制之一[15,16]。IFN-γ的抗衣原體活性超越色氨酸耗竭,包括干擾細(xì)胞能量代謝,導(dǎo)致后代減少,但也降低了細(xì)胞的抗菌敏感性[17]。IFN-γ通過信號轉(zhuǎn)導(dǎo)和轉(zhuǎn)錄激活因子1的激活,顯著增加IDO的表達(dá)[18]。IFN-γ能顯著激活角質(zhì)形成細(xì)胞的IDO表達(dá),而非成纖維細(xì)胞[19]。Sakural等[20]發(fā)現(xiàn)IFN-γ在實驗性自身免疫性腦脊髓膜炎發(fā)病的臨床前期達(dá)到高峰,緩解期降低。而IDO的升高在時相上正好在IFN-γ之后。IFN-γ誘導(dǎo)IDO表達(dá)的特點是表達(dá)強度較高及表達(dá)持續(xù)的時間較為短暫。因而,IFN-γ誘導(dǎo)IDO的表達(dá)有助于宿主限制微生物的生長及預(yù)防急性炎癥對宿主潛在的損害,但I(xiàn)FN-γ誘導(dǎo)IDO表達(dá)的特點不能解釋IDO在體內(nèi)的長期效應(yīng),即維持包括妊娠和自身耐受等免疫穩(wěn)態(tài)[2]。
眾所周知,母胎界面有Th1 型細(xì)胞因子和Th2 型細(xì)胞因子。Th1 型細(xì)胞因子包括IFN-γ、TNF-α、TNF-β、IL-2及IL-12。Th2型細(xì)胞因子,包括IL-4、IL-5、IL-6 及IL-10。正常情況下,母胎界面的細(xì)胞因子向Th2 型偏倚,即母胎界面Th2 型細(xì)胞因子較Th1 型細(xì)胞因子占優(yōu)勢[21]。傳統(tǒng)認(rèn)為,受孕后,母體對胎兒的識別及妊娠的維持是通過占優(yōu)勢的Th2 型細(xì)胞因子來實現(xiàn)的,母胎界面的Th2 型細(xì)胞因子是維持同種異體耐受的核心,Th2 型偏倚的重要意義在于維持妊娠和抑制母體和胎兒之間的排斥反應(yīng),即維持母胎耐受[22,23]。我們的研究結(jié)果提示,Th1 型細(xì)胞因子(IFN-γ)亦可通過調(diào)節(jié)IDO的表達(dá)致母胎耐受,從而保護(hù)胎兒免受母體的排斥。因而,本研究進(jìn)一步完善了母胎耐受的機(jī)制。
總之,本研究發(fā)現(xiàn)早孕絨毛和蛻膜組織中IFN-γ和IDO的表達(dá)呈正相關(guān),并在體外培養(yǎng)的該組織中發(fā)現(xiàn)IFN-γ可上調(diào)IDO的表達(dá)。本研究結(jié)果表明,在早孕母胎界面IFN-γ可通過上調(diào)IDO的表達(dá)來致母胎耐受。
猜你喜歡
艦船科學(xué)技術(shù)(2022年16期)2022-09-22 02:15:00
北京航空航天大學(xué)學(xué)報(2021年6期)2021-07-20 07:23:54
當(dāng)代陜西(2020年13期)2020-08-24 08:22:02
制造技術(shù)與機(jī)床(2017年5期)2018-01-19 02:49:17
制造技術(shù)與機(jī)床(2017年11期)2017-12-18 06:47:29
金秋(2017年4期)2017-06-07 08:22:16
蘇州科技大學(xué)學(xué)報(自然科學(xué)版)(2017年1期)2017-03-20 15:25:18
中國材料進(jìn)展(2016年10期)2016-12-26 06:50:20
濰坊學(xué)院學(xué)報(2016年2期)2016-12-01 13:00:11
新聞傳播(2015年11期)2015-07-18 11:15:04
