DNMT1 和DNMT3a 在結直腸癌中的表達、臨床意義及相關性研究
2020-02-21 10:55:24高曉斌武雪亮趙軼峰聶雙發劉小飛竇金山張迎春
中國醫藥導報 2020年1期
關鍵詞:研究
高曉斌 武雪亮 趙軼峰 聶雙發 梁 峰 劉小飛 竇金山 張迎春
1.河北北方學院附屬第一醫院普通外科,河北張家口 075000;2.河北北方學院附屬第一醫院小兒外科,河北張家口 075000;3.河北北方學院研究生學院,河北張家口 075000
據最新流行病學調查顯示,2014 年我國居民結直腸癌發病例數位居第五,嚴重威脅國民健康[1]。結直腸癌的早期診斷、規范化治療和預后監測,對患者生存時間、生活質量有著重要影響。從分子生物學水平,尋找精準度高、特異性強的標志物已成為結直腸癌診療領域的研究熱點,同時為腫瘤的靶向治療提供新的技術支撐[2-4]。
DNA 甲基轉移酶(DNMTs)是人類表觀遺傳學中催化并維持DNA 基化結構和功能的重要酶家族[5]。DNMTs包含3個重要成員:DNMT1、DNMT2 和DNMT3,其中DNMT1 是目前研究最為廣泛也最為深入的酶類,其在DNA 修復和正常甲基化功能過程中發揮關鍵作用;DNMT2 主要負責tDNA 的甲基轉移,DNMT3 內含3 個亞基,3a、3b 和3L,3a、3b 與啟動子區CpG 島、DNA 的異常甲基化關系密切,而3L 系調節蛋白[6]。研究證實,DNMT1、DNMT3a、DNMT3b 的活性增強或表達上調能介導DNA 異常甲基化,與人類諸多腫瘤的發生、發展密切相關[7-10],但在結直腸癌中的研究較少。本研究通過對結直腸癌和正常組織中DNMT1、DNMT3a 行實時熒光定量逆轉錄聚合酶鏈反應(qRTPCR)和免疫組織化學檢測分析,旨在探討二者在結直腸癌中的表達情況及與相關臨床病理因素間的關系,為結直腸癌的早期診斷和預后監測提供可靠的分子生物學標志物,為結直腸癌的靶向診療提供科學參考。
1 資料與方法
1.1 一般資料
共納入2017 年2 月~2018 年2 月河北北方學院附屬第一醫院(以下簡稱“我院”)普通外科手術切除并經病理確診的結直腸癌組織(癌組)和癌旁正常組織(正常組)標本,各50 例,同時收集患者完整的臨床資料。本研究經我院醫學倫理委員會審核批準。
1.2 主要試劑和儀器
RNA 逆轉錄試劑購自日本Takara 公司;PCR 試劑盒、PrimeScriptTMRT reagent 試劑盒購自日本Takara公司;DNMT1、DNMT3a 和β-actin 引物由生工生物(北京)有限公司設計合成;全自動電化學發光免疫分析儀購自瑞士羅氏公司E2010;DEPC 水購自美國Sigma 公司;兔抗人DNMT1 單克隆抗體、兔抗人DNMT3a 多克隆抗體均購自美國Santa Cruz 公司;DAB顯色試劑盒購自北京中杉公司。
1.3 實驗方法
1.3.1 qRT-PCR 按TRIzol RNA 試劑盒說明書提取總RNA,用PrimeScriptTMRT reagent 試劑盒逆轉錄合成cDNA 2 μL 逆轉錄產物進行PCR 擴增,PCR 擴增引物如下,DNMT1 正向引物:5′-CGATGAGGAAGTCGATGATAACAT-3′,反向引物:5′-ATGCGATTCTTGTTCTGTTTCTTCT-3′;DNMT3a 正向引物:5′-GCCCAAGGTCAAGGAGATTATTG-3′,反向引物:5′-TCTGCCGCACCTCGTACAC-3′;β-actin 作為內參,正向引物:5′-AGGAAGCTTCCAGACACGC-3′,反向引物:5′-CGCACTGCCATTTTAGCTCC-3′。PCR 反應條件:95℃變性10 min,進行40 個循環:95℃變性15 s,60℃退火2 s,72℃延伸40 s,然后根據SYBRGreen 染色法行相對定量分析,熒光定量PCR 實驗數據處理使用2-△△Ct法。
1.3.2 免疫組織化學法 將癌組織標本連續切片4 μL貼附于經多聚賴氨酸處理的玻片上,80℃烘烤50 min,光鏡下觀察切片中DNMT1、DNMT3a 蛋白的表達和分布情況,每張切片選取5 個高倍視野(400×)。DNMT1 蛋白陽性主要分布于細胞核中,而DNMT3a蛋白陽性主要分布于細胞質和細胞膜中,均呈棕黃色顆粒。判斷標準:按染色強度,無著色為0 分,淡黃為1 分,深黃2 分,棕褐或棕黃3 分;陽性細胞計數,<10%為0 分,10%~<25%為1 分,25%~<50%為2 分,50%~<75%為3 分,≥75%為4 分。兩者相加<2 分為陰性(-),2~3 分為弱陽性(+),4~5 分為中等強度陽性(++),6~7 分為強陽性(+++),由兩名具備高級職稱的病理醫師經雙盲法獨立評分。
1.4 統計學方法
采用SPSS 17.0 統計軟件進行統計分析,計量資料以均數±標準差()表示,采用t 檢驗,計數資料以百分率表示,采用χ2檢驗。相關性分析應用Spearman檢驗。以P <0.05 為差異有統計學意義。
2 結果
2.1 結直腸癌和癌旁正常組織中DNMT1 mRNA、DNMT3a mRNA 表達
以目標基因與β-catenin 的比值作為qRT-PCR定量分析結果,癌組中DNMT1 mRNA 的表達水平(0.36±0.07)明顯高于正常組(0.09±0.01),差異有統計學意義(t=10.951,P=0.000);癌組中DNMT3a mRNA 的表達水平(0.33±0.06)明顯高于正常組(0.11±0.02),差異有統計學意義(t=8.724,P=0.000)。
2.2 結直腸癌和癌旁正常組織中DNMT1 和DNMT3a 蛋白的表達
免疫組化結果顯示,癌組中DNMT1 蛋白陽性表達為84.00%(42/50),明顯高于其在正常組中的表達[24.00%(12/50)],差異有統計學意義(χ2=21.545,P=0.000);癌組中DNMT3a 蛋白陽性表達為78.00%(39/50),明顯高于其在正常組中的表達[30.00%(15/50)],差異有統計學意義(χ2=18.433,P=0.000)。見圖1。
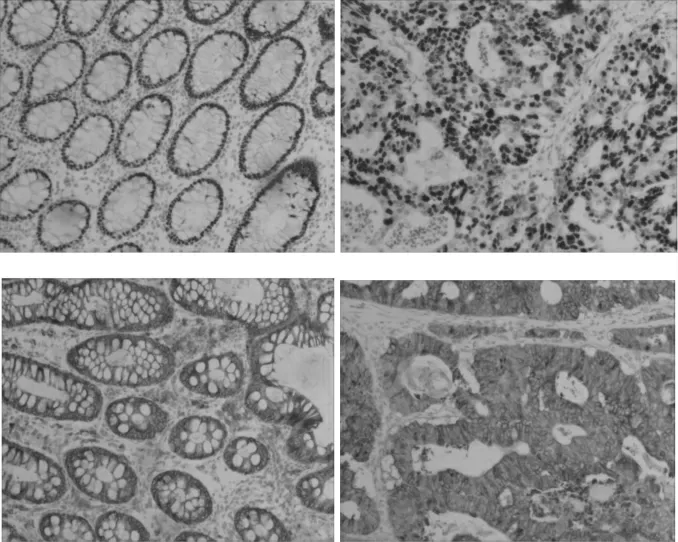
圖1 DNMT1、DNMT3a 在結直腸正常組織和癌組織中的表達(SP,200×)
2.3 DNMT1 和DNMT3a 蛋白表達與結直腸癌患者臨床病理特征的關系
DNMT1 和DNMT3a 蛋白表達均與腫瘤的浸潤深度、TNM 分期、淋巴結轉移及肝轉移相關(均P <0.05)。TNM 分期越高,二者的表達水平越高(均P <0.05),伴淋巴結轉移、肝轉移的患者二者的表達水平明顯高于未轉移的患者(均P <0.05),而二者的表達水平與患者腫瘤大小、分化程度均無相關性(均P >0.05)。見表1。
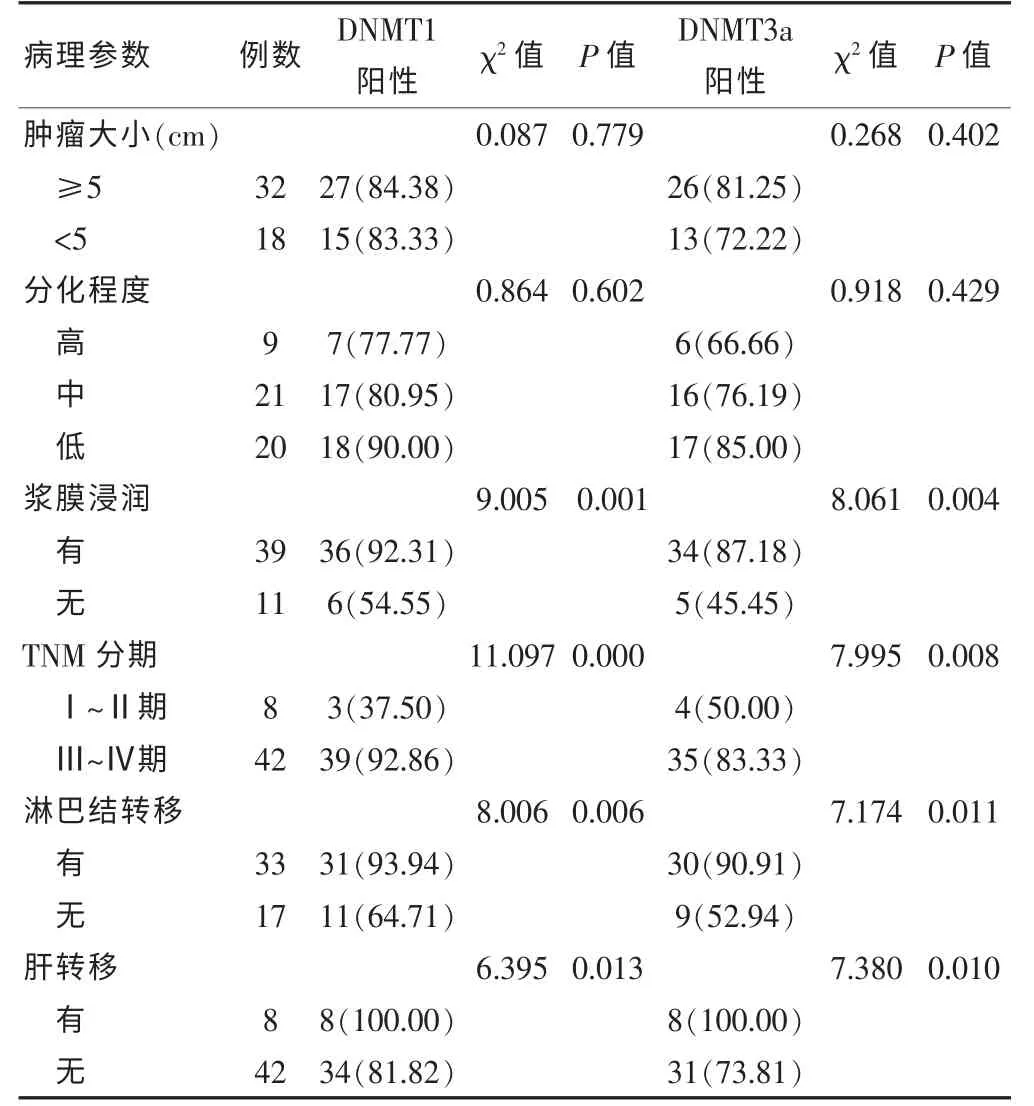
表1 DNMT1 和DNMT3a 蛋白表達與結直腸癌患者臨床病理特征的關系[例(%)]
2.4 結直腸癌組織中DNMT1 和DNMT3a 蛋白表達的相關性
DNMT1 和DNMT3a 蛋白表達呈正相關(r=0.455,P=0.000)。見圖2。
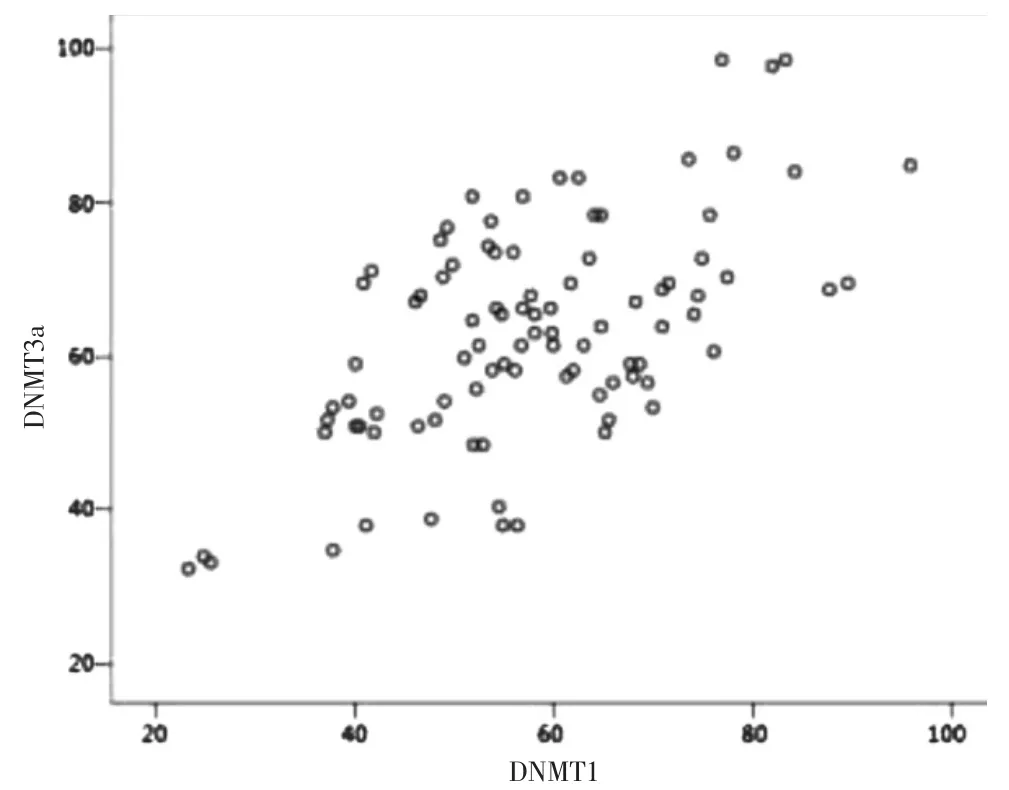
圖2 結直腸癌組織中DNMT1 和DNMT3a 蛋白的相關性
3 討論
結直腸癌的發生、發展是多個基因調控的多步驟、多階段的復雜病變過程,其中抑癌基因的失活和致癌基因的激活發揮關鍵作用,其病變機制涉及表觀遺傳學的改變,包括DNA 甲基化、染色體重塑、組蛋白修飾和lncRNA 調控[11-14],其中DNA 甲基化是目前研究最為深入的機制。
DNMT1 是由Bestor 等[15]于1988 年在真核生物中克隆出胞嘧啶特異DNA 轉移酶,即DNMT1。DNMT1 定位于人染色體19p13.2~19p13.3,內含1616 個氨基酸殘基,相對分子量為183×103,包括3 個結構域,即N 端的某些特定蛋白識別靶區域、C 端的催化端和相對的未知區域[16-17]。DNMT1 主要介導甲基化模式的建立與維護,而DNMT3a 主要負責從頭甲基化,二者高表達可以誘導甲基化模式異常、致癌基因激活和抑癌基因沉默,從而導致腫瘤發生。
研究發現DNMT1 在人類諸多腫瘤的發生、發展中發揮重要作用。DNMT1 是DNA 甲基化的關鍵作用酶,其活性的增加能促進DNA 異常甲基化。Li 等[18]通過對胰腺癌組織和相應正常組織行免疫印跡和免疫組化法檢測,發現約80%、78.7%的癌組織中DNMT1呈高表達狀態,表明DNMT1 的異常高表達與胰腺癌的發生密切相關。Yang 等[19]應用采用免疫組織化學法檢測54 例胃癌患者石蠟切片中DNMT1 和DNMT3a的表達,發現DNMT1、DNMT3a 在胃癌組織中的過表達分別為54 例(64.8%)、38 例(70.4%),且二者與TNM 分期和淋巴結轉移顯著相關。馮悅靜等[20]應用酶聯免疫吸附試驗法檢測136 例肺癌患者和147 名健康對照者中DNMT1 和DNMT3a 的蛋白表達,發現二者蛋白表達水平均顯著高于正常對照組,進一步logistic 回歸結果證實,二者蛋白的高表達能增加肺癌的發病率。
本研究顯示DNMT1 和DNMT3a 在結直腸癌組織中表達明顯高于正常組,表明在腸黏膜癌變的過程中,細胞核中的DNMT1 和細胞質、細胞膜上的DN MT3a 水平逐步上調,啟動DNA 異常甲基化過程,相應的調控基因表達受抑制,從而導致腫瘤的發生;進一步研究顯示,DNMT1 和DNMT3a 表達水平與結直腸癌的浸潤深度、TNM 分期、淋巴結和肝轉移均明顯相關,且二者表達呈明顯正相關,提示DNMT1 和DNMT3a的表達與結直腸癌的臨床病理參數相關,二者共同參與建立、維持整個DNA 甲基化模式的過程,二者表達越高,病變越嚴重、預后越差。DNMT1 和DNMT3a同時作用的甲基化酶效應是各自單獨作用的近5 倍,DNMT3a 介導從頭甲基化的啟動后,半甲基化DNA底物激活DNMT1 活性使之完成后續的甲基化修飾;而DNMT1 能夠催化未修飾的DNA 發生甲基化,DNMT3a 的從頭甲基化活性則是通過甲基化的DNA 與DNMT1 酶位于N 端的變構位點的靶向結合而被激活的。
綜上,DNMT1 和DNMT3a 的異常表達對結直腸癌的發生和進展有重要作用。本研究僅從基因和蛋白水平分析二者在結直腸癌組織中的表達情況,其具體的異常甲基化作用機制及相關通路的研究尚不明確,這也是筆者下一步的研究方向。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
