鋁脅迫造成橡膠苗死亡的機制研究
2020-02-22 03:19:20張婷婷劉子凡安鋒謝貴水
熱帶作物學報 2020年12期
張婷婷 劉子凡 安鋒 謝貴水

摘? 要:鋁毒是酸性土壤中抑制植物生長和產量的主要因子,但其對橡膠幼苗生長的影響研究甚少。本研究設置50、100、200 mmol/L 3種Al3+處理濃度,采用水培試驗研究了鋁脅迫下橡膠幼苗葉片葉綠素含量、葉片相對含水量、根系活力、根系導水率、H2O2和O2?含量變化。結果表明:葉片SPAD值、葉片相對含水量、根系活力、根系導水率隨脅迫時間的延長而逐漸降低,H2O2和O2?含量則逐漸增加,且濃度越高對各個指標的影響越顯著。鋁毒害可造成根系活性氧(H2O2和O2?)的累積,進而造成根系活力和導水率下降,葉片SPAD值和相對含水量下降,最終導致橡膠幼苗死亡。
關鍵詞:鋁毒;橡膠苗;SPAD值;根系活力;根系導水率;活性氧
中圖分類號:S718.43? ? ? 文獻標識碼:A
Abstract: Aluminum toxicity is a main factor that inhibits plant growth and productivity in acid soils, leaf relative water content, root activity, root hydraulic conductivity, H2O2, and O2? content were studied by hydroponic experiments. The results showed that the leaf SPAD value, leaf relative water content, root activity and root hydraulic conductivity gradually decreased with the prolonged stress time, and the contents of H2O2 and O2? gradually increased, the higher the concentration, the more significant impact of each index. Taking together, the results showed that aluminum could cause the accumulation of active oxygen species (H2O2 and O2?) in the rubber tree root system, which in turn led to a decreased in root activity and water hydraulic conductivity, resulting in a decrease in leaf SPAD value and relative water content, and eventually the death of rubber tree saplings.
Keywords: aluminum toxicity; rubber tree saplings; SPAD value; root activity; root hydraulic conductivity; active oxygen species
DOI: 10.3969/j.issn.1000-2561.2020.12.010
鋁(Al)在地殼中含量極為豐富,自然情況下生物可利用性不高[1]。但在酸性環境下(pH<5.0),土壤中難溶的鋁轉變為游離的鋁離子對植物生長造成損害,截至目前,我國酸性土壤的面積約占全國可耕土壤面積的21%[2]。鋁毒成為抑制作物生長的主要因素[3-4]。前人已有研究表明,鋁脅迫對桉樹(Eucalyptus robusta)[5]、茶樹(Camellia sinensis)[6-7]、玉米(Zea mays)[8]、水稻(Oryza sativa)[9]、大豆(Glycine max)[10]、大麥(Hordeum vulgare L)[11]、馬尾松(Pinus massoniana)[12]等作物生長有抑制作用。鋁毒害最先作用于與其直接接觸的植物根系[13-14],最明顯的特征是:根尖膨大、根系伸長受阻、根系活力及根系導水率降低[15-16]。而葉綠素在植物光合作用中充當著相當重要的角色,光合作用的強弱直接取決于葉綠素含量的高低[17]。由于植物體內葉綠素對鋁脅迫的敏感性[18],植物受鋁毒的損害程度常可通過植物體內的葉綠素含量反映[19]。有研究發現,鋁毒害可以降低葉片葉綠素含量、抑制植物光合作用,進而對植物的生長產生影響[20],但只有超過相應的閾值才會抑制植物的生長[21]。
橡膠樹(Hevea brasiliensis)原產于巴西亞馬遜河流域[22],是目前唯一大面積商業化種植生產天然橡膠的樹種,在中國主要種植于南方的云南、廣東以及海南[23],該區土壤環境多為酸性,因此橡膠樹在其生長過程中難以避免地出現鋁毒害的現象,張晗等[24]、安鋒等[25]通過盆栽試驗研究了鋁脅迫對橡膠幼苗葉綠素熒光、膜脂過氧化、丙二醛、過氧化物酶等相關生理指標的影響,發現200~400 mmol/L高濃度鋁脅迫處理會導致橡膠苗抗氧化防御系統被破壞,顯著降低了活性氧清除相關酶的活性以及葉綠素含量和熒光特性;橡膠苗細胞質膜透性、MDA、葉片游離脯氨酸和可溶性糖含量顯著提高,并在處理3~5 d后死亡,推測鋁脅迫下活性氧的積累可能造成橡膠苗活性氧清除系統破壞、根系導水率下降、葉片干枯進而造成橡膠幼苗死亡,但并未對鋁脅迫下橡膠幼苗根系活性、根系導水率以及活性氧(H2O2和O2?)含量進行直接測定。為此,本研究設置不同濃度鋁脅迫,測定了鋁脅迫下橡膠幼苗葉片相對含水量、根系活性、根系導水率和根系活性氧(H2O2和O2?)含量的變化,以期闡明鋁脅迫造成橡膠幼苗死亡的機制,為橡膠園土壤改良、橡膠樹耐鋁品種的選擇和栽培提供依據。
1? 材料與方法
1.1? 材料
橡膠幼苗為‘熱研7-33-97無性系組培苗,由中國熱帶農業科學院橡膠研究所提供。
1.2? 方法
1.2.1? 試驗設計? 試驗于2019年6至10月在海南省海口市中國熱帶農業科學院橡膠研究所恒溫培養箱內進行。恒溫培養箱設白天溫度25 ℃、晚上溫度20 ℃,光照12 h,強度400 μmol/m2·s。取沙培2蓬葉的橡膠組培苗,洗凈,移至裝有5 L營養液的桶中預培養,桶的規格為:上部直徑29 cm,下部直徑21 cm,高20 cm,預培養的營養液為:含有40 μmol/L AlCl3·6H2O(防止鋁處理可能出現的休克作用[26])pH 5.5的Hoagland營養液[配方:2.8 mg/L H3BO3,3.4 mg/L MnSO4·H2O,0.1 mg/L CuSO4·5H2O,0.22 mg/L ZnSO4·7H2O,0.1 mg/L (NH4)6Mo7O24·4H2O,6.72 mg/L Na2EDTA,5.58 mg/L FeSO4·7H2O,9.4 mg/L Ca(NO3)2·4H2O,5.2 mg/L MgSO4·7H2O,6.6 mg/L KNO3,1.2 mg/L NH4H2PO4,現配現用],連續增氧通氣,每3 d更換一次營養液,培養5 d。
每個處理16株苗,設置3種Al3+濃度處理,并以不加Al3+ pH為5.5和4.2的T1、T2為對照,因為在前期大田調研發現,膠園土壤pH為5.5,極端條件下為4.2。此外,也為排除培養液pH變化對橡膠幼苗生長的影響(詳見表1),分別在處理6、24、48、72、96 h后測定相關生理生化指標。
1.2.2? 項目測定? 每個處理取苗4株,剪下所有葉片和根系,測定其鮮重和烘干后干重。計算出相對含水量。葉綠素含量用SPAD-502葉綠素儀(Konica-Minota, Tokyo, 日本)測定,用SPAD值表示。根系活力測定采用TTC法[27];根系導水率的測定采用壓力室法[28];過氧化氫(H2O2)含量采用北京索萊寶生物公司提供的過氧化氫檢測試劑盒(微量法)進行測定;超氧陰離子(O2?)含量采用羥胺氧化法[29]測定。
1.3? 數據處理
采用SAS 9.4軟件的Duncan新復極差法對測定數據進行單因素方差分析(P<0.05),用Origin 2016和Excel軟件作圖。
2? 結果與分析
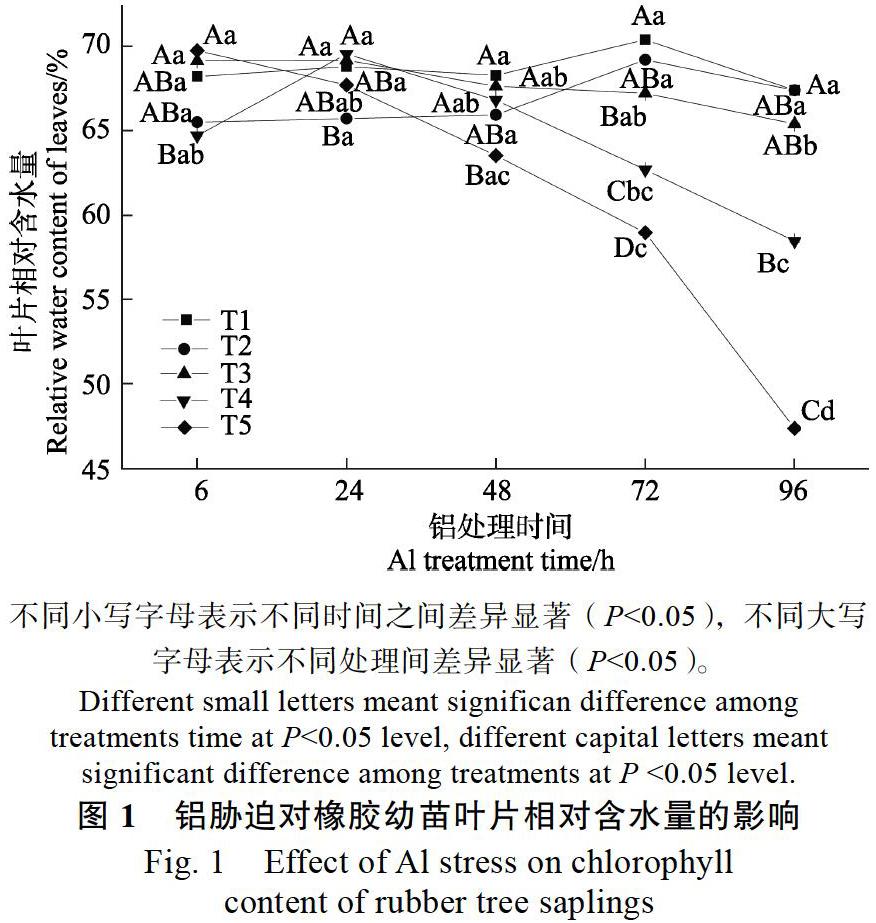
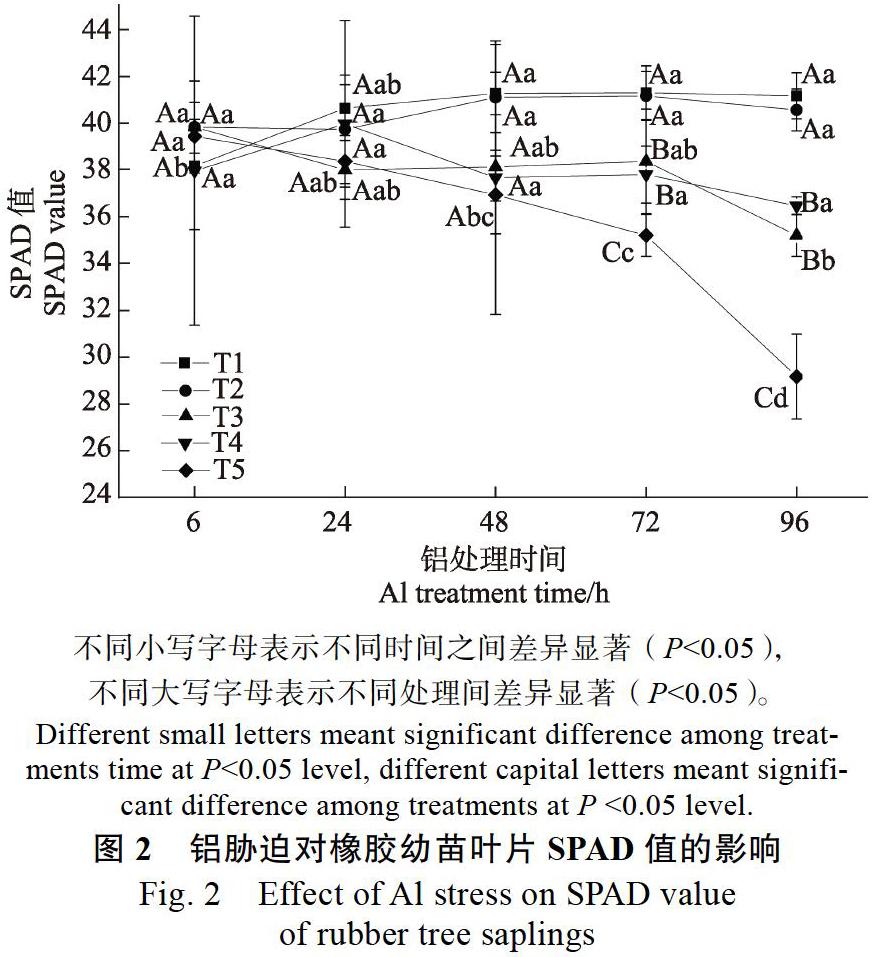
2.1? 鋁脅迫對橡膠苗葉片SPAD值及相對含水量的影響
由圖1、圖2可以看出,與T1處理相比,T2處理對橡膠幼苗葉片SPAD值及葉片相對含水量無顯著影響,說明處理液酸度由5.5降至4.2不會影響橡膠幼苗葉片SPAD值和相對含水量。而在不同濃度鋁脅迫下橡膠幼苗葉片SPAD值及葉片相對含水量均會出現不同程度的下降。各濃度鋁處理下,脅迫24 h前對橡膠幼苗葉片相對含水量和SPAD值的影響均不顯著,但鋁脅迫下均隨著處理時間的延長呈下降趨勢。T3鋁處理96 h時,葉片相對含水量顯著降低;T4脅迫72 h,橡膠幼苗葉片相對含水量就顯著降低;而脅迫濃度為T5時,處理48 h便可顯著降低橡膠幼苗葉片相對含水量。鋁處理對葉片SPAD值的影響略有不同,脅迫濃度為T4時,處理96 h顯著降低SPAD值;T5脅迫72 h時,顯著降低葉片SPAD值。其中,鋁離子脅迫濃度為T5時,對橡膠幼苗葉片SPAD值和葉片相對含水量的影響最為顯著,在鋁脅迫48 h后顯著降低,葉片SPAD值在T5鋁脅迫處理48、72、96 h時,分別比同一pH值T2未加鋁處理6 h時的葉片SPAD值降低了6.3%、10.7%和26.0%,而葉片相對含水量比同一pH值T2未加鋁處理6 h時降低了9.0%、15.0%、32.0%。說明高濃度鋁脅迫會降低橡膠幼苗葉片SPAD值及葉片相對含水量,且抑制作用隨時間的延長而逐漸加強。
2.2? 鋁脅迫對橡膠苗根系活力和根系導水率的影響
葉片相對含水量和SPAD值的下降可能由于根系活力的下降引起,為此我們比較了不同濃度鋁脅迫對橡膠苗根系活力的影響。由表2可以看出,與T1相比,T2溶液處理對根系活力有一定的促進作用或沒有顯著影響。而鋁脅迫處理則會使橡膠幼苗根系活力均顯著下降。而且T4、T5濃度下根系活力顯著低于T3,但T4和T5濃度之間差異不顯著。在根系活力隨鋁脅迫處理時間的變化方面,T1、T2、T3、T4鋁脅迫處理24 h時,橡膠幼苗根系活力較處理6 h有所提高,增幅分別為50.43%、50.49%、33.24%、17.05%,表明相同處理時間鋁毒害對橡膠幼苗根系活力的影響強于培養液酸度的變化,而隨后T3、T4隨著處理時間的延長根系活力受到抑制而降低; T3鋁處理96 h橡膠苗根系活力顯著降低;T4處理72 h后橡膠幼苗根系活力顯著降低;T5鋁脅迫下橡膠幼苗根系活力隨時間的延長前期無顯著差異,但在處理96 h后顯著下降。T3、T4、T5鋁離子處理幼苗96 h,其橡膠幼苗根系活力分別為T2無鋁脅迫的41.80%、15.20%和9.60%。其說明T3、T4和T5三種濃度鋁脅迫在6 h前可能已經造成橡膠苗根系活力下降,鋁脅迫對橡膠幼苗根系活力的抑制隨濃度增加而加強,并且隨著處理時間的延長整體呈下降趨勢。
根系活力的下降可能會導致根系導水率的下降。由表3可知,鋁脅迫對根系導水率的影響與根系活力的影響基本一致。與T1相比,T2溶液對根系導水率有一定提高或沒有顯著變化。而與鋁脅迫對根系活力影響一致,從脅迫6 h開始,T3、T4、T5鋁脅迫均造成橡膠幼苗根系導水率的顯著下降,并且隨著處理時間的延長,T3、T4均未表現出顯著差異,只有T5在處理96 h根系導水率顯著降低。鋁脅迫處理96 h時,T3、T4、T5鋁脅迫處理的橡膠幼苗根系導水率分別是pH 4.2不加鋁處理的29.13%、6.25%、0.60%;T5處理96 h橡膠幼苗根系導水率比處理6 h時下降了96.52%,根系已基本完全喪失吸水能力。說明與根系活力變化一致處理在6 h前已經造成橡膠幼苗根系導水率顯著降低,并且在T5處理96 h后根系已基本完全死亡,喪失吸水能力。
2.3? 鋁脅迫對橡膠苗根系H2O2和O2?含量的影響
根系活力和導水率的下降可能由于根系活性氧的累積引起,為此我們研究了不同濃度鋁處理下根系H2O2和O2?含量的變化。由圖3的結果可以看出,低pH和鋁脅迫均會使橡膠幼苗根系H2O2含量增加,T2處理24 h時使根系H2O2含量顯著增加,但隨著處理時間的延長,T2處理的根系H2O2含量與T1差異不顯著;而T3、T4、T5鋁脅迫處理均會使橡膠苗根系H2O2含量始終顯著高于不加鋁pH 5.5 T1溶液;說明酸度降低也會造成H2O2的累積,但酸性條件下鋁脅迫也會造成H2O2的累積。T5鋁脅迫下處理24 h根系H2O2含量就顯著高于T2;而處理24 h后,隨著處理時間的延長各種濃度鋁脅迫下橡膠幼苗根系H2O2含量均顯著高于T2,但T4、T5鋁脅迫下橡膠幼苗根系H2O2含量無顯著差異。在T3、T4、T5鋁脅迫處理96 h時,橡膠幼苗根系H2O2含量是T2的1.52、1.79、1.91倍。表明鋁脅迫會造成膠幼苗根系H2O2累積,對植物可能產生毒害影響。
鋁脅迫對根系O2?的影響見圖4,可以看出,T2處理24 h時,橡膠幼苗根系超氧陰離子含量(O2?)與T1差異不顯著且高于T3鋁脅迫處理;而處理48 h時,T2處理的根系O2?含量增加并顯著高于T1;至處理96 h時,根系O2?含量與T1無顯著差異。說明培養液酸度由5.5降低到4.2也會影響根系O2?含量的變化。此外,不同濃度鋁脅迫下根系O2?含量隨著處理時間的延長呈上升趨勢,在鋁脅迫處理96 h前,各種濃度鋁脅迫下橡膠幼苗根系O2?含量差異不顯著,但在鋁脅迫處理96 h時,T3、T4、T5根系O2?含量顯著高于T1、T2;而且T4、T5的根系O2?含量顯著高于T3,T4、T5根系O2?含量差異不顯著,說明鋁脅迫也會造成橡膠苗根系O2?的累積,而且高濃度比低濃度鋁對橡膠苗根系O2?含量的影響更大。
3? 討論
活性氧(ROS)累積是植物在逆境脅迫下作出的氧化應激反應,ROS累積過多會使植物的細胞質膜被破壞而影響到植物生長和發育[30],而植物體清除活性氧依賴于植物自身的超氧化物歧化酶(SOD)、過氧化氫酶(CAT)和過氧化物酶(POD)等抗氧化酶組成的ROS清除系統。在先前的研究中,我們測定了不同濃度鋁脅迫處理下橡膠幼苗SOD、POD、CAT的活性,發現SOD、CAT和POD的活性隨著處理液中鋁濃度的增加而顯著下降,并且隨著處理時間的延長呈現出先升高后下降的趨勢[24-25, 31]。說明短時間低濃度處理可使SOD、POD和CAT的活性升高,清除植物體內多余的ROS,但隨著鋁處理濃度的增加和時間的延長,植株體內的抗氧化酶活性失調,使得ROS產生和清除失衡導致ROS在植株體內累積,最終造成植株生長發育受阻。H2O2和O2?是ROS在植物體內不同的存在形式,本試驗直接測定了鋁脅迫下橡膠幼苗H2O2和O2?含量的變化,結果顯示:與pH 4.2不添加鋁離子的處理相比,鋁脅迫可以顯著增加橡膠幼苗根系H2O2和O2?含量,并且隨著處理時間的增長,根系H2O2和O2?含量呈上升趨勢,其中,200 mmol/L Al3+脅迫處理橡膠幼苗96 h,其根系H2O2和O2?的含量分別是pH 4.2不加鋁離子處理的1.94和1.94倍。這一實驗結果證實了隨著處理時間的延長,ROS會在橡膠苗根系中累積,有可能破壞植物體內的抗氧化酶清除系統,使得ROS不能被及時清除而在植株體內大量累積,可能對植物產生毒害。前人對大豆在鋁脅迫下ROS積累的研究以及夜間高溫下鈣對番茄幼苗ROS含量影響的報道也證明了這一結論[32, 33]。
植物根系是直接與鋁離子接觸的部位,植物根系根尖部位受鋁毒損害最為嚴重[34]。而根系導水率是植物吸收和傳導水分能力的一個重要指標[35],關系著植株能否正常生長發育。有研究表明,ROS累積可造成水通道蛋白的門控效應,造成水通道蛋白活性下降,根系水分的吸收運輸受到抑制,從而降低植物根系導水率[36]。而根系導水率的降低會致使葉片缺水,氣孔開度減小從而抑制植物生長[37-39]。本研究結果表明:不同濃度鋁脅迫處理,可顯著降低橡膠幼苗的根系活力,其中200 mmol/L鋁離子脅迫96 h對橡膠幼苗根系活力的抑制作用最強,僅為pH 4.2不添加鋁處理的9.60%。這與前人報道的鋁脅迫造成烤煙以及大豆的根系活力下降的研究結果相符[40-41]。鋁毒害造成根系活力的下降可顯著降低橡膠幼苗根系導水率,與pH 4.2不加鋁處理相比,不同濃度的鋁脅迫橡膠幼苗在6 h時根系導水率已經顯著下降,并且隨著處理時間的延長,有進一步下降的趨勢;其中,鋁離子濃度為200 mmol/L處理24、48、72和96 h時橡膠幼苗根系導水率分別比處理6 h下降了31.61%、41.39%、13.70%和96.52%,說明處理時間越長對根系導水率的抑制作用是疊加的、不可逆的,最終造成根系導水能力的喪失和橡膠苗的死亡。這些試驗結果與其他重金屬脅迫下根系導水率下降的報道相同[42]。值得注意的是,Gunse等[43]對玉米根系水分轉運特性的研究發現,鋁脅迫處理幾分鐘或幾小時后會導致根系導水率降低,而本實驗結果發現,鋁脅迫處理在6 h前可能已經造成橡膠幼苗根系導水率以及根系活性降低,但鋁脅迫造成橡膠幼苗根系導水率和根系活性降低的確切時間仍需進一步的實驗驗證。
葉綠素含量是植物能否進行正常生長發育的決定因素之一,SPAD值可以反映葉片葉綠素含量及其變化趨勢[44]。而葉片相對含水量變化可作為判斷植物非生物脅迫耐受度,植物細胞含水量降低是因為細胞的失水,細胞失水會造成植物生長受阻[45]。本試驗結果表明:200 mmol/L鋁脅迫處理48 h就能使橡膠葉片SPAD值和相對含水量顯著降低,早于50 mmol/L的96 h和100 mmol/L的72 h;鋁脅迫處理48 h后隨著脅迫時間的延長對橡膠幼苗葉片SPAD值和相對含水量有顯著的抑制作用,表明鋁離子濃度越高對橡膠苗SPAD值和相對含水量抑制作用越強。這與Chen[46]報道的鋁脅迫造成植物嫩芽SPAD值和相對含水量降低的結果一致。此外,本研究結果也證明了張晗等[24]、安鋒等[25]報道的橡膠幼苗對鋁毒害的耐受濃度為100~200 mmol/L。
綜上,鋁脅迫會造成橡膠幼苗根系H2O2和O2?的累積和ROS清除系統的破壞,抑制橡膠幼苗根系活性,使得根系導水率下降沒有充足的水分向上運輸到葉片,造成橡膠苗葉片相對含水量降低、葉片缺水枯黃,葉綠素含量降低,植株不能正常進行光合作用,進而影響整個植株的正常生長和發育,造成橡膠苗死亡。
參考文獻
羅? 亮, 謝忠雷, 劉? 鵬, 等. 茶樹對鋁毒生理響應的研究[J]. 農業環境科學學報, 2006, 25(2): 305-308.
趙天龍, 解光寧, 張曉霞, 等. 酸性土壤上植物應對鋁脅迫的過程與機制[J]. 應用生態學報, 2013, 24(10): 3003-3011.
應小芳, 劉? 鵬, 徐根娣. 土壤中的鋁及其植物效應的研究進展[J]. 生態環境, 2003, 12(2): 237-239.
Halman J M, Schaberg P G, Hawley G J, et al. Calcium and aluminum impacts on sugar maple physiology in a northern hardwood forest[J]. Tree Physiology, 2013, 33(11): 1242- 1251.
徐圓圓, 陸明英, 蔣維昕, 等. 鋁脅迫下不同耐鋁型桉樹無性系根和葉抗氧化特征的差異[J]. 浙江農林大學學報, 2016, 33(6): 1009-1016.
于翠平, 潘志強, 陳? 杰, 等. 鋁對茶樹生長與生理特性影響的研究[J]. 植物營養與肥料學報, 2012, 18(1): 182-187.
吳瓊鴦, 鄭偉偉, 羅? 亮, 等. 鋁對茶樹根系生理的影響[J]. 湖北農業科學, 2005(3): 80-82.
劉保國, Vesna Hadzi-Taskovic Sukalovic. 鋁對玉米生長和硝酸還原酶活性的影響[J]. 植物生理學報, 1998, 24(4): 347-353.
石貴玉. 鋁對水稻幼苗生長和生理的影響[J]. 廣西植物, 2004, 24(1): 77-80.
田? 聰, 張? 爍, 粟? 暢, 等. 鋁脅迫下大豆根系有機酸積累的特性[J]. 大豆科學, 2017, 36(2): 256-261.
Clarkson D T. Effect of aluminum on the uptake and metabolism of phosphorus by barley seedlings[J]. Plant Physiology, 1996, 41(1): 165-172.
張盛楠, 劉亞敏, 劉玉民, 等. 馬尾松幼苗生長及生理特性對鋁脅迫的響應[J]. 西北植物學報, 2016, 36(10): 2022-2029.
Kollmeier M, Felle H H, Horst W J. Genotypical differences in aluminum resistance of maize are expressed in the distal part of the transition zone. Is reduced basipetal auxin flow involved in inhibition of root elongation by aluminum[J]. Plant Physiology, 2000, 122(3): 945-956.
Rengel Z. Disturbance of cell Ca2+ homeostasis as a primary trigger of Al toxicity syndrome[J]. Plant, Cell and Environment, 1992, 15: 931-938.
唐劍鋒, 林咸永, 章永松, 等. 小麥根系對鋁毒的反應及其與根細胞壁組分和細胞壁對鋁的吸附?解吸性能的關系[J]. 生態學報, 2005, 25(8): 1890-1897.
孟長軍. 鋁脅迫對白苦瓜幼苗生長狀況和生理特性的影響[J]. 中國農業科技導報, 2018, 20(8): 23-28.
初曉輝, 張艾青, 段新慧, 等. 鋁脅迫對多花黑麥草生長和生理的影響[J]. 草原與草坪, 2017, 37(6): 48-56.
曹? 林, 吳玉環, 章? 藝, 等. 外源水楊酸對鋁脅迫下菊芋光合特性及耐鋁性的影響[J]. 水土保持學報, 2015, 29(4): 260-266.
張云華, 孟慶玲, 計成林, 等. 鋁脅迫對轉C4磷酸烯醇式丙酮酸羧化酶基因水稻光合作用和活性氧代謝的影響[J]. 生物技術通報, 2014(7): 81-85.
劉? 強, 胡? 萃, 柳正葳, 等. 鋁脅迫對煙草葉片光能利用、光保護系統及活性氧代謝的影響[J]. 華北農學報, 2017, 32(1): 118-124.
劉? 強, 尹? 麗, 龍婉婉, 等. 鋁脅迫對蓼科植物生長和光合、蒸騰特性的影響[J]. 廣西植物, 2011, 31(2): 227-232.
李明謙, 張鳳良, 李小琴, 等. 橡膠樹無性系苗期生長和光合生理特性的比較[J]. 西南林業大學學報(自然科學), 2020, 40(2): 1-5.
貝美容, 蔡? 雋, 戚春林, 等. 2種新型種植材料(苗木)橡膠幼樹的施肥效應[J]. 熱帶作物學報, 2019, 40(11): 2105-2111.
張? 晗, 安? 鋒, 袁? 坤, 等. 不同鋁水平脅迫對橡膠樹幼苗若干生理指標的影響[J]. 熱帶作物學報, 2014, 35(10): 1992-1996.
安? 鋒, 李昌珍, 張婷婷, 等. 鋁脅迫對橡膠苗生理和葉綠素熒光特性的影響[J]. 應用生態學報, 2018, 29(12): 4191-4198.
徐鏡波, 袁曉凡, 郎佩珍. 過氧化氫酶活性及活性抑制的紫外分光光度測定[J]. 環境化學, 1997, 16(1): 73-76.
郝再彬, 蒼? 晶, 徐? 仲. 植物生理實驗[M]. 哈爾濱: 哈爾濱工業大學出版社, 2004.
胡田田, 康紹忠. 局部灌水方式對玉米不同根區土-根系統水分傳導的影響[J]. 農業工程學報, 2007, 23(2): 11-16.
李忠光, 龔? 明. 植物中超氧陰離子自由基測定方法的改進[J]. 云南植物研究, 2005, 27(2): 211-216.
Vale G, Mehennaoui K, Cambier S, et al. Manufactured nanoparticles in the aquatic environment-biochemical responses on freshwater organisms: A critical overview[J]. Aquatic Toxicology, 2016, 170(23): 162-174.
彭? 艷, 李? 洋, 楊廣笑, 等. 鋁脅迫對不同小麥SOD、CAT、POD活性和MDA含量的影響[J]. 生物技術, 2006, 16(3): 38-42.
楊? 喆, 王芳妹, 王? 瀚, 等. H2O2參與大豆根尖鋁毒調節的研究[J]. 大豆科學, 2011, 30(1): 105-108.
李天來, 高曉倩, 劉玉鳳. 夜間低溫脅迫下鈣對番茄幼苗根系活力及活性氧代謝的調控作用[J]. 西北農業學報, 2011, 20(8): 127-132.
沈玉芳, 曲? 東, 王保莉, 等. 干旱脅迫下磷營養對不同作物苗期根系導水率的影響[J]. 作物學報, 2005, 31(2): 214-218.
楊啟良, 張富倉, 劉小剛, 等. 環境因素對植物導水率影響的研究綜述[J]. 中國生態農業學報, 2011, 19(2): 456-461.
Boursiac Y, Prak S, Boudet J, et al. The response of Arabidopsis root water transport to a challenging environment implicates reactive oxygen species-and phosphorylation-dependent internalization of aquaporins[J]. Plant Signal Behavior, 2008, 3(12): 1096-1098.
Veselova S V, Farhutdinov R G, Veselov S Y, et al. The effect of root cooling on hormone content, leaf conductance and root hydraulic conductivity of durum wheat seedlings (Triticum durum L.)[J]. Journal of Plant Physiology, 2005, 162(1): 21-26.
Lovelock C E, Ball M C, Feller I C, et al. Variation in hydraulic conductivity of mangroves: Influence of species, salinity, and nitrogen and phosphorus availability[J]. Physiologia Plantarum, 2006, 127(21): 457-464.
Martre P, North G B, Nobel P S. Hydraulic conductance and mercury-sensitive water transport for roots of opuntia acanthocarpa in relation to soil drying and rewetting1[J]. Plant Physiology, 2001, 126(1): 352-362.
李淮源, 劉柏林, 鄧世媛, 等. 鋁脅迫對烤煙生長和光合特性的影響[J]. 煙草科技, 2015, 48(9): 9-13.
俞慧娜, 劉? 鵬, 徐根娣. 大豆生長及葉綠素熒光特性對鋁脅迫的反應[J]. 中國油料作物學報, 2007, 29(3): 257-265.
孔祥瑞, 曲? 東, 周莉娜. 硫營養對重金屬脅迫下玉米和小麥根系導水率的影響[J]. 西北植物學報, 2007, 27(11): 2257-2262.
Gunse B, Poschenrieder C, Barcelo J. Water transport properties of roots and root cortical cells in proton- and Al-stressed maize varieties[J]. Plant Physiology, 1997, 113(2): 595-602.
賈? 朋, 錢? 磊, 羅樹凱, 等. 洋紫荊葉綠素含量的分布特征及與葉片氮含量的關系[J]. 中國城市林業, 2019, 27(11): 1-7.
Rahman A, Mostofa M G, Nahar K, et al. Exogenous calcium alleviates cadmium-induced oxidative stress in rice (Oryza sativa L.) seedlings by regulating the antioxidant defense and glyoxalase systems[J]. Brazilian Journal of Botany, 2016, 39(2): 393-407.
Chen L S. Physiological responses and tolerance of plant shoot to aluminum toxicity[J]. Journal of Plant Physiology and Molecular Biology, 2006(2): 143-155.
