光強對耐蔭差異普通鈍葉草(Stenotaphrum helferi)種質的形態與生理差異分析
2020-02-22 07:15:35楊娟王志勇郭海林丁西朋楊虎彪吳楊廖麗
熱帶作物學報 2020年12期
關鍵詞:形態
楊娟 王志勇 郭海林 丁西朋 楊虎彪 吳楊 廖麗


摘? 要:對2個耐蔭性具有顯著(P<0.05)差異的普通鈍葉草(Stenotaphrum helferi)品系S17(耐蔭強)與S34(耐蔭弱)設置3個不同程度(自然光照、70%自然光照、25%自然光照)遮光處理,對其形態及生理生化差異進行分析。結果表明,經遮蔭處理后耐蔭性強的S17相對葉長、相對葉寬、相對綠色覆蓋度、相對草坪密度、相對坪用質量、相對地上干物重及相對光合色素含量均顯著(P<0.05)高于耐蔭性弱的S34,且重度脅迫顯著(P<0.05)高于輕度脅迫;遮蔭后S17的相對葉綠素a/b逐漸增加,而S34逐漸降低;重度遮蔭后,相對可溶性糖及相對可溶性蛋白含量顯著(P<0.05)低于輕度脅迫,其中S17顯著(P<0.05)高于S34。本研究對今后開展普通鈍葉草耐蔭新品種選育和QTL定位提供參考。
關鍵詞:普通鈍葉草;耐蔭性;形態;生理
中圖分類號:S688.4? ? ? 文獻標識碼:A
Abstract: Three shading treatment, natural light, 70% natural light and 25% natural light, was applied to two Stenotaphrum helferi accessions with significant difference in shade tolerance. The morphological, physiological and biochemical characteristics of S. helferi were analyzed. The relative leaf length, relative leaf width, relative green coverage, relative turf density, relative turf quality, relative dry matter weight and relative photosynthetic pigment contents of S17 with strong shade tolerance were significantly higher than those of S34 with shade sensitive, and those of severe stress were significantly higher than those of light stress. The relative chlorophyll a/b of S17 increased gradually, while S34 decreased gradually after shade. The content of relative soluble sugar and relative protein was significantly lower than that of nature light, and S17 with strong shade tolerance was higher than S34 with shade sensitive. This study would provide a reference for future breeding and QTL mapping of new shade-tolerant cultivars.
Keywords: Stenotaphrum helferi; shade tolerance; morphology; physiology
DOI: 10.3969/j.issn.1000-2561.2020.12.012
光照是植物生長發育的重要環境因子之一,影響植物的整個生長進程。城市綠化中的高大建筑與植物,常使草坪大面積處于弱光生境中。弱光環境雖然會對草坪草的正常生長造成壓力,但大多數草坪草可以通過改變自身形態結構和代謝進程來適應逆境。比如遮蔭會使草坪垂向生長,匍匐枝減少,葉片變得細長且薄,葉面積增大[1],葉重增加[2-3]。遮蔭后到達植物葉片的光量子通量密度降低,導致植物生長發育中的生理生化過程發生改變,增加葉片的光合色素含量[4],使植物能進行正常光合作用。當光合效率提高,植物的呼吸效率則會減少,植物內部的滲透調節機制會發生相應改變,更多的可溶性糖被積累[5-6],保持了植物內部的滲透平衡,更好地適應弱光環境。
普通鈍葉草(Stenotaphrum helferi)為禾本科多年生草本植物,分布于熱帶或者亞熱帶地區。該草種耐蔭性強[7-8],植株低矮,匍匐莖產量高[9],管理要求粗放,成本投入較低[8],具有耐水淹、耐旱、繁殖快、生命力強、耐踐踏等優點[10-13],但其抗寒性較差[14-15],則常在南方被應用于生態環境綠化中[16]。目前國內外已對鈍葉草的耐蔭性種質篩選及形態表觀進行研究[10, 17],但鈍葉草耐蔭性的響應機制方面研究較少。不同草坪草在弱光脅迫中會表現不同的生物學特征[18-19],同一植物的不同品種之間也會存在差異[20]。因此,開展選育耐蔭品種及其分析其應對弱光脅迫形態與生理上的響應機制,為后續植物耐蔭機制的深入探究提供一定理論依據。本研究擬從形態與生理上開展對普通鈍葉草耐蔭性的響應機制初步研究,以期為篩選優質鈍葉草耐蔭品種及分子響應機制提供一定理論依據。
1? 材料與方法
1.1? 材料
1.1.1? 試驗地概況? 試驗地點在海南省海口市海南大學試驗基地大棚,地理坐標20°03′N,110°19′E,海拔2.8 m;屬熱帶海洋性季風氣候,試驗時間為2019年4至7月,期間大棚內最低溫度為22 ℃,最高溫度為39 ℃,相對濕度約85%。
1.1.2? 試驗材料? 試驗材料為普通鈍葉草品系S17(耐蔭強)與S34(耐蔭弱),是在實驗室前期篩選出的耐蔭性具有顯著差異的2個鈍葉草品系,分別采于廣西、云南,現保存于海南大學農科實驗基地。
1.1.3? 材料的培養及處理? 試驗采用盆栽方式,土壤基質配制比例體積比河沙∶田園土∶椰糠為1∶1∶1,混合均勻后裝入塑料盆(高25 cm,直徑22.5 cm)中,每盆栽種5節生長健壯、整齊一致的鈍葉草匍匐莖段,待培養約3個月成坪后方可用于后續試驗。
試驗為二因素隨機區組試驗,A因素為2個不同品系,B因素為不同程度遮光脅迫。各處理用不同規格的遮蔭網搭建(遮蔭棚長1.8 m,寬1.2 m,高1.2 m),用照度計(型號:TES-1334A)測定光照強度。其中對照為100%自然光,處理1(T1)的光照度為70%自然光;處理2(T2)的光照度為對照的25%自然光。生長期間對其進行統一的水肥管理。分別在處理后的20 d(5月8日)、40 d(5月28日)、60 d(6月17)和80 d(7月8日)對形態和生理生化指標進行測定。
1.2? 方法
測定指標參照張靜等[21]、廖麗等[22]試驗方法進行,分別在4個時期進行葉長、葉寬、綠色覆蓋度、草坪密度、坪用質量、地上部鮮重、葉綠體色素可溶性糖及可溶性蛋白的測定,具體方法如下:
葉長、葉寬(cm):米尺測量;綠色覆蓋度和草坪密度(%):目測法,指單位面積上草坪植物綠色葉片的數量和單位面積上草坪植物個體及枝條的數量,采用百分比制;坪用質量:目測法,參照NTEP(The National Turfgrass Evaluation Program,美國國家草坪評比項目)標準,以草坪密度、質地、顏色、均一性等為指標進行綜合評分。最好質量為9分,6分為可以接受的草坪質量,0分為草坪死亡;地上部鮮重(g):剪取處理80天后的地上部分稱重;葉綠素和胡蘿卜素含量(mg/g):分光光度計測定;可溶性糖含量(μg/g):蒽酮比色法;可溶性蛋白含量(μg/g):考馬斯亮藍染色法。
1.3? 數據處理
相對葉長、相對葉寬、相對綠色覆蓋度、相對草坪密度、相對地上部鮮重、相對葉綠素含量、相對類胡蘿卜素含量、相對葉綠素a/b、相對可溶性糖含量及相對可溶性蛋白含量計算方法:遮光處理后的平均值/自然光照下的平均值× 100%。試驗數據采用SPSS(IBM SPSS Statistics 20)軟件進行方差分析,Excel 2016軟件進行分析處理。
2? 結果與分析
2.1? 不同遮光強度對普通鈍葉草葉片性狀的影響
圖1為S17與S34的相對葉長與相對葉寬的變化。由圖1A中可知,在輕度脅迫時,隨著脅迫時間的增加,S17的相對葉長先增加后下降,再逐漸增加,在80 d時達到峰值,顯著(P<0.05)高于此時期的S34;S34的相對葉長則是先增加,20 d后下降,40 d后再增加,60 d后顯著(P<0.05)下降的趨勢,在20 d時為最大峰值。
其中,40 d與80 d時S17的相對葉長顯著(P<0.05)高于S34。重度脅迫時,S17與S34的變化一致,均為先增加,20 d后下降,40 d后再增加,60 d后又下降的趨勢,其中S17的相對葉長在60 d時達到峰值,顯著(P<0.05)高于同處理其他3個時期,而S34在20 d時便達到峰值。在重度脅迫40 d前,2個品系的相對葉長差異不顯著,在處理60~80 d時S17則顯著(P<0.05)高于S34。
如圖1B中所示,S17在2個不同程度遮蔭脅迫下的相對葉寬變化趨勢均為先增加后降低,S34在2個不同程度處理下的變化趨勢也一致,呈先增加,20 d后降低,40 d時再增加,60 d后又降低的趨勢,且2個品系均在60 d時達到峰值。在輕度脅迫20 d時,S17的相對葉寬顯著(P< 0.05)低于S34,到40 d時,S17便顯著(P<0.05)高于S34,40 d后,2品系逐漸趨于穩定,無顯著差異,其中S17的相對葉寬要略高于S34。重度脅迫下,S17的相對葉寬均高于S34,且80 d時顯著(P<0.05)高于S34。
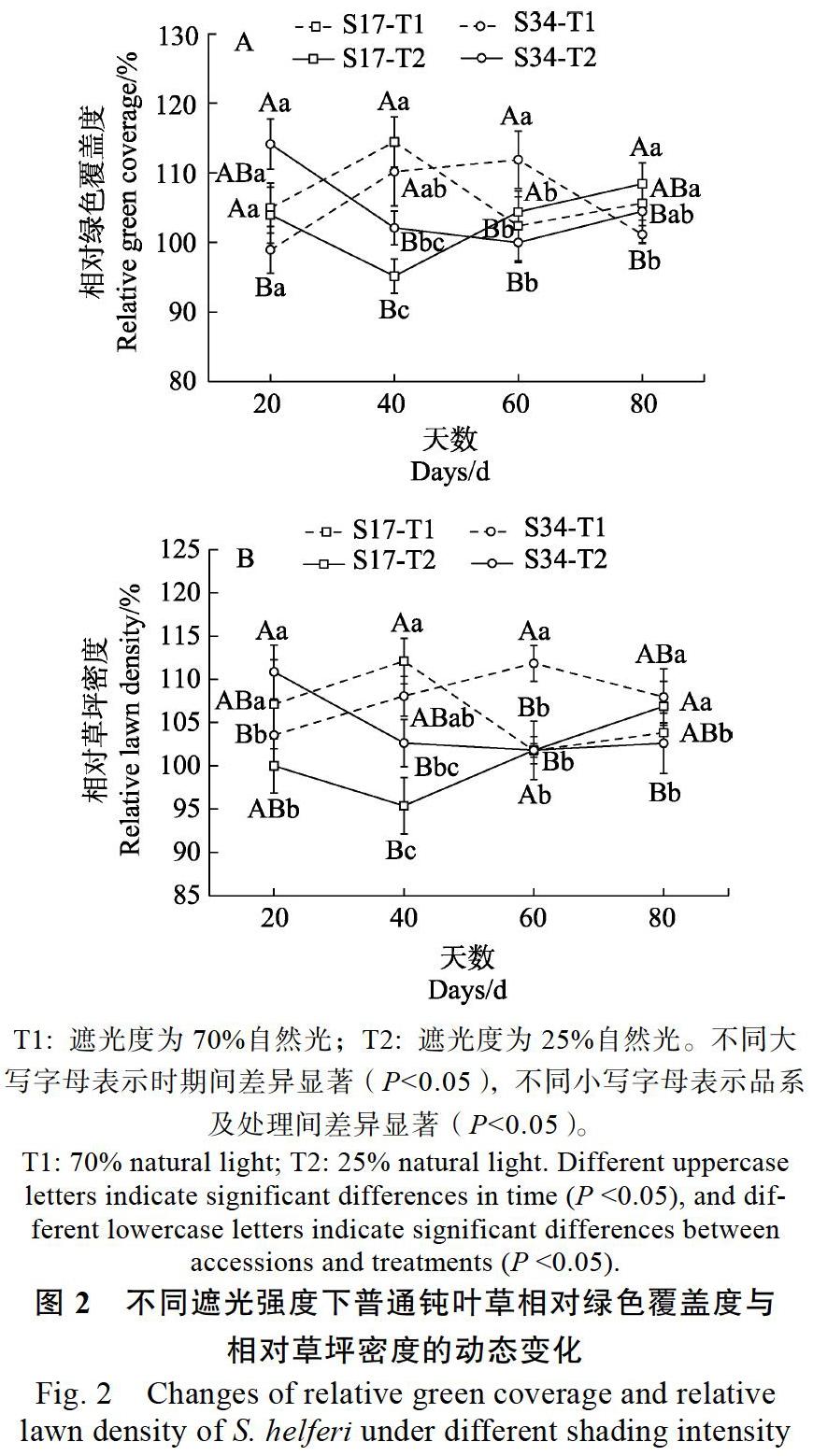
2.2? 不同遮光強度對普通鈍葉草成坪狀況的影響
如圖2A中顯示,輕度脅迫時,S17的相對綠色覆蓋度呈先上升,40 d后下降,至60 d時再上升的趨勢,處理40 d時達最大值;S34則呈先上升,60 d后逐漸下降的趨勢,且60 d的相對綠色覆蓋度顯著(P<0.05)高于同時期的S17。重度脅迫時,S17呈先上升,而20 d后呈“V”型變化,最低值在處理40 d時,顯著(P<0.05)低于同處理的其它時期;S34在重度脅迫20 d時的相對綠色覆蓋度顯著(P<0.05)高于同處理的其它時期,20 d后開始逐漸下降,60 d時有逐漸上升的趨勢。
2個品系的相對草坪密度(圖2B),在輕度脅迫時S17呈先上升,40 d后下降,60 d后再上升趨勢,處理40 d時達最大峰值;S34在輕度脅迫60 d前一直呈上升趨勢,60 d后開始逐漸下降。其中,在40 d前S17的相對草坪密度要高于S34,且在20 d時顯著(P<0.05)高于S34,而60~80 d期間S34要高于S17,且S34在60 d的相對草坪密度顯著(P<0.05)高于S17。重度脅迫時,S17的變化趨勢為先上升,20 d后下降,40 d時再上升趨勢,處理40 d時為最低值;而S34的相對草坪密度在重度遮光20 d時顯著(P<0.05)高于同處理的其它時期,20 d后開始下降,40 d后趨于穩定,波動幅度較小。
2.3? 不同遮光強度對普通鈍葉草坪用質量的影響
如圖3,在輕度遮光脅迫時,S17的相對坪用質量逐漸增加,S34則是先增加后下降,其中40~60 d期間相對穩定,60 d后逐漸下降。2個品系經輕度遮光60 d前,S34各時期的相對坪用質量均高于S17,且40 d與60 d的顯著高于同時期的S17,到80 d時S17才略高于S34。在重度脅迫時,S17同樣呈逐漸增加趨勢,且60~80 d期間的相對坪用質量顯著(P<0.05)高于同處理的20 d與40 d;S34則呈先增加,60 d后下降的趨勢。其中2個品系在經重度遮光60 d前各時期的相對坪用質量無顯著差異,60 d后S34則顯著(P<0.05)下降,到80 d時顯著(P<0.05)低于S17。
2.4? 不同遮光強度對普通鈍葉草地上部重量的影響
從圖4可看出,2個品系在不同程度遮蔭脅迫下的相對地上部鮮重均存在顯著(P<0.05)差異。耐蔭性強的S17在重度脅迫下的相對地上部鮮重顯著(P<0.05)高于2個處理下的S34及輕度脅迫下的S17;輕度脅迫下S34的相對地上部鮮重顯著(P<0.05)高于重度脅迫下的S34及輕度脅迫下的S17。則重度遮光有利于耐蔭性強的S17地上部生長,輕度遮光更有利于耐蔭性弱的S34地上部生長。
2.5? 不同遮光強度對普通鈍葉草葉綠體色素的影響
如圖5A中顯示,輕度脅迫時S17的相對葉綠素(a+b)含量隨脅迫時間增加呈先上升,40 d后下降并趨于平穩的變化趨勢;S34呈先上升,40 d后下降趨勢,且80 d時的相對葉綠素(a+b)含量顯著(P<0.05)低于同時期的S17以及S34的同處理的前3個時期。重度脅迫時,S34的相對葉綠素(a+b)含量在前20天持續增加,20 d后則開始下降,60 d后又稍增長趨勢;S17的相對葉綠素(a+b)含量則隨遮蔭天數的增加一直保持上升趨勢,且在40 d、60 d及80 d時的相對葉綠素含量均顯著(P<0.05)高于同時期重度脅迫下的S34。
2個品系經遮光處理后的相對類胡蘿卜素含量均呈先上升后下降趨勢(圖5B)。輕度脅迫時,S17與S34在40~60 d期間的相對類胡蘿卜素含量均顯著(P<0.05)高于各自在20 d與80 d的相對類胡蘿卜素含量,其中S17在20 d、40 d與80 d時的相對類胡蘿卜素含量顯著(P<0.05)高于此時的S34。重度脅迫時,2個品系的相對類胡蘿卜素含量峰值均在60 d,總體可以看出,重度遮光下S17的相對類胡蘿卜素含量在各時期均顯著(P<0.05)高于同處理的S34。
如圖5C所示,輕度脅迫時,隨遮蔭天數的增加,S17的相對葉綠素a/b在處理20 d后逐漸下降,20~80 d時均顯著(P<0.05)低于20 d的相對葉綠素a/b。而S34則呈先增加,60 d后下降趨勢,且在60 d時達到峰值,顯著(P<0.05)高于20 d與40 d的相對葉綠素a/b。其中,在脅迫20 d時S17的相對葉綠素a/b要顯著(P<0.05)高于S34,而S17的相對葉綠素a/b逐漸降低,到60~80 d均顯著(P<0.05)低于S34。重度脅迫時,S17呈先增加,20 d后降低,40 d時再增加,60 d后又降低的趨勢,其中20 d時為峰值,顯著(P<0.05)高于重度脅迫下S17的其他時期。S34則逐漸增加,80 d的相對葉綠素a/b顯著(P<0.05)高于S34的前3個時期。其中20 d時S17顯著(P<0.05)高于S34,而S34的相對葉綠素a/b逐漸增加到80 d則顯著(P<0.05)高于同脅迫程度的S17。
2.6? 不同光照強度對普通鈍葉草滲透調節物質的影響
耐蔭差異的普通鈍葉草經遮光處理后的可溶性糖含量除輕度脅迫60 d時S17的可溶性糖含量外,其他時期均低于對照(表1)。隨遮蔭時間的增加,S17在2個不同程度脅迫下的相對可溶性糖含量的變化趨勢均呈先增加后降低趨勢,均在60 d時達到最大峰值,顯著(P<0.05)高于同程度處理的其他3個時期。而S34在輕度脅迫下的變化趨勢為先增加,再降低后又增加的趨勢,在40 d時達最大峰值。重度脅迫時則呈先增加后降低,再增加后又降低趨勢,且20 d的相對可溶性糖含量顯著(P<0.05)高于后3個時期。其中輕度脅迫60 d時S17的相對可溶性糖含量顯著(P<0.05)高于S34,重度脅迫20 d時,S34相對可溶性糖含量顯著高于S17,而后S34逐漸降低,到40 d時顯著(P<0.05)低于S17,到60 d后2品系雖無顯著差異,但S17在60 d與80 d的相對可溶性糖含量均高于S34。
隨遮蔭天數的增加,S17的相對可溶性蛋白含量在2個不同程度遮光處理下,均呈先上升后下降趨勢,且均在60 d時達最大峰值,顯著(P<0.05)高于同一處理的其他時期。2個不同程度遮蔭脅迫下S34的相對可溶性蛋白含量變化趨勢一致,均呈先上升后下降,再上升而后又下降的趨勢,同樣60 d時的相對可溶性蛋白含量顯著(P<0.05)高于同一處理的其他時期。在輕度脅迫20 d時,S17的相對可溶性蛋白顯著(P<0.05)低于S34,而后S17逐漸增加,脅迫到40 d到60 d時均顯著(P<0.05)高于S34。重度脅迫20 d與80 d時,2個品種的相對可溶性蛋白含量無顯著差異(P>0.05),而在40 d與60 d時S17要顯著(P<0.05)高于S34。
3? 討論
草坪草在弱光環境中會表現出不同的生物學特征,但同一草坪草的不同品種之間也會存在差異。選育耐蔭性強的鈍葉草品種,了解其應對弱光脅迫形態與生理上的響應,為鈍葉草耐蔭性的深入探究提供一定的理論依據。
本研究發現不同光照強度對耐蔭性差異顯著的普通鈍葉草品系的葉長、葉寬產生不同影響。數據顯示,耐蔭性強的S17的葉長與葉寬均高于自然光照組,其相對葉長與相對葉寬在遮光處理60 d前總體呈增長趨勢,且重度遮光各時期的葉片性狀顯著高于輕度遮光。說明當光強減弱,植株所能利用的光能減少時,耐蔭性強的品系會通過改變葉片形狀,增加葉面積以捕獲更多光能,提高光合效率,增加蒸騰面積,這一結果與Trenholm等[23]結論一致。耐蔭性弱的S34的相對葉長與相對葉寬在遮蔭后期80 d時要顯著低于處理20 d時,可看出總體呈下降趨勢,且隨遮光程度的增加導致S34相對葉寬逐漸降低。說明耐蔭性較弱的品系在脅迫前期能適度增加葉長及葉寬以適應逆境,長時間的遮蔭會顯著抑制其生長,這與Cai等[24]及陳斌等[25]結果相同。其中耐蔭性弱的品系隨著光強減弱,葉片逐漸變細,則可將葉寬作為鈍葉草耐蔭品種篩選的評價指標之一
植物在有限光照條件下通過光合作用產生的有限的光合產物,耐蔭性強的植物會將有限的能量更多地分配給地上部分,以獲取更多的光量子來適應遮蔭環境[25-26]。在草坪草中主要是以增加葉面積以獲得更多光能,從而地上部的莖葉要優先獲得能量[27],而其中草坪的密度、匍匐莖減少與光照強度減弱及遮蔭時間導致的光合產物、物質積累減少密切相關。本研究中2個鈍葉草品系的綠色覆蓋度、草坪密度及處理40 d后的草坪質量都高于自然光照組,其中耐蔭性強的S17在長時間遮蔭下能持續保持穩定的相對綠色覆蓋度與相對草坪密度,相對草坪質量隨遮蔭天數增加逐漸增加,且各時期重度遮光的相對草坪質量顯著高于輕度遮光組。這與Trenholm[28]研究中的優良鈍葉草品種結果一致。 而S34的相對綠色覆蓋度及相對草坪密度及相對草坪質量在輕度脅迫時能隨遮蔭時間增加而增加,60 d后才稍下降,且后期的成坪狀況優于S17。這說明輕度遮蔭相較于2個品系來說,耐蔭性弱的S34能更有效的進行光合作用,而在長時間的重度遮蔭時,耐蔭性弱的品系將更多的光能產物用于增加葉面積,則枝條數減少,密度下降,坪用質量相對也下降,而耐蔭性強的品系則能在重度遮光環境中充分利用有限光能,生物量能均勻分配,從而保證良好的草坪質量。
遮蔭能提高植物葉片的葉綠素含量,在草坪草應用方面,一定程度上增加了草坪的綠色景觀效果[1]。本研究中,耐蔭性強的S17鈍葉草的葉綠素與類胡蘿卜素含量隨遮蔭程度及天數的增加而逐漸增加,且各時期重度脅迫顯著高于輕度脅迫及同一遮蔭處理時的S34。說明植株通過增加光合色素的含量來提高光合作用能力,促進物質的積累和葉片的生長,以獲得更多的光能,保證其適應性。而耐蔭性弱的S34的葉綠素與類胡蘿卜素含量隨遮蔭程度增加而增加,而隨著遮蔭時間的增加則呈先上升后下降變化趨勢。則可能是因為草坪草的葉綠素是處于一個不斷合成和分解的動態變化中,耐蔭性弱的品系隨著遮光程度的增加,遮光時間的延長,使其葉綠素含量出現一個波動的過程[29]。葉綠素a/b的比值是體現植物適應弱光環境的一個重要指標。S17的葉綠素a/b比值隨遮蔭程度增加及遮蔭時間的延長逐漸降低,S34則逐漸增加。遮蔭使鈍葉草的光合色素含量增加,而葉綠素a/b的變化趨勢則相反,則是由于弱光脅迫導致植物陰生色素葉綠素b含量增加,能有效吸收弱光環境中的藍紫光。則說明耐蔭性強的品系能在長時間弱光脅迫中增加自身葉綠素b以保證地上部生長,而耐蔭性弱的品系由于自身對弱光環境的敏感導致只能適應一定時長。
植物葉片中的可溶性糖及可溶性蛋白作為滲透調節物質參與植物應對逆境脅迫時的一系列生理響應機制中,二者的合成均受到光的調控。適度的光強有利于其含量的增加[30],也有研究顯示可溶性糖及可溶性蛋白含量會隨光照強度的減弱逐漸降低[31-32]。本研究中耐蔭性強的S17的相對可溶性糖含量和相對可溶性蛋白質含量遮蔭60 d內逐漸上升,且在各時期重度脅迫的相對可溶性糖含量和相對可溶性蛋白質顯著低于輕度脅迫。而耐蔭性弱的S34遮蔭后的相對可溶性糖含量和相對可溶性蛋白質變化較為波動。其中S17在同一遮蔭強度下各時期的相對可溶性糖含量及相對可溶性蛋白含量均高于耐蔭性弱的S34。在弱光逆境中,光合速率高,呼吸消耗減少,則可溶性糖更多被用于物質的積累,說明鈍葉草在遮蔭條件下具有較強的適應性。結合研究中形態指標變化規律,耐蔭性強的鈍葉草品系可溶性糖與蛋白含量與形態的變化趨勢大致呈負相關,說明遮蔭為其提供了較好的滲透調節平衡條件,能夠積累較多的可溶性糖、蛋白,以用于物質的量合成與積累,使其對弱光逆境有良好的適應性,從而具有較高的草坪質量。而耐蔭性較弱的品系在弱光逆境中,能保持一段時間的正常生長,而在長時間脅迫下自身的調節機制以不足以保持其正常生長,從而在后期生長中各項指標逐漸下降。
對不同光照強度下2個耐蔭性差異顯著的普通鈍葉草品系的形態及生理指標的分析表明,耐蔭性強的S17在重度遮光脅迫時,隨遮蔭時間的延長,其葉片性狀、成坪狀況、坪用質量、光合色素含量及滲透調節物質含量都要優于輕度遮光下的S17,而耐蔭性弱的S34,在輕度遮光脅迫中能保持良好生長,而在長時間重度遮光條件下,生長狀況受到明顯抑制。說明2個耐蔭性不同的普通鈍葉草品系都有不同的光適應性,不同程度的弱光逆境對2個品系都起到了促進作用,在輕度遮光時,耐蔭性弱的S34的總體生長狀況要略優于耐蔭性強的S37。在重度遮光時,耐蔭性強的S17的生長狀況顯著優于S34,且長期遮蔭后尤為顯著。
參考文獻
周興元, 曹福亮, 陳志明, 等. 遮蔭對幾種暖地型草坪草成坪速度及其景觀效果的影響[J]. 草原與草坪, 2003, 23(2): 26-29.
范? 彥, 周壽榮. 川西地區三種野生草坪地被植物耐蔭性的研究[J]. 中國草地, 1999, 21(5): 3-5.
嚴? 潛. 吉祥草對光照強度適應性的研究[D]. 長沙: 湖南農業大學, 2007.
張? 爽. 槭葉草對干旱和遮陰的適應性研究[D]. 長春: 吉林農業大學, 2014.
德力格爾, 李媛媛, 張天俊, 等. 遮陰對成坪期草地早熟禾和紫羊茅生理指標的影響[J]. 草原與草坪, 2015, 35(5): 50-54, 59.
姜? 丹, 陳雅君, 劉? 丹, 等. 光氮互作對草地早熟禾碳氮代謝的影響[J]. 中國草地學報, 2005, 27(6): 49-53.
Glenn B. Effect of shade intensity and duration on warm-season turfgrass growth and development[D]. California: University of Florida, 2015.
Yu X, Kimball J A, Milla-Lewis S R. High density genetic maps of St. Augustinegrass and applications to comparative genomic analysis and QTL mapping for turf quality traits[J]. BMC Plant Biology, 2018, 18(1): 346.
Wang Z, Raymer P, Chen Z. Isolation and characterization of microsatellite markers for Stenotaphrum Trin. using 454 sequencing technology[J]. HortScience, 2017, 52(1): 16-19.
黃? 娟, 夏漢平, 蔡錫安. 遮光處理對三種鈍葉草的生長習性與光合特性的影響[J]. 生態學雜志, 2006, 25(7): 759-764.
胡化廣, 張振銘. 大穗結縷草對鹽脅迫響應及臨界鹽濃度的研究[J]. 北方園藝, 2010(3): 80-83.
徐? 特. 不同暖季型運動草坪綜合質量的對比研究[J]. 當代體育科技, 2018, 8(18): 225-226.
席嘉賓, 羅? 耀, 張巨明. 幾種常綠草坪草對干旱脅迫的生理響應[J]. 草原與草坪, 2011, 31(2): 28-32.
喬雪云, 李海梅. 三種暖季型草坪草的抗寒性研究[J]. 北方園藝, 2012(13): 64-67.
Kimball J A , Tuong T D , Arellano C , et al. Linkage analysis and identification of quantitative trait loci associated with freeze tolerance and turf quality traits in St. Augustinegrass[J]. Molecular Breeding, 2018, 38(5): 67.
楊秀云, 武小鋼. 遮蔭條件下草坪的建植與管理[J]. 北方園藝, 2004(1): 36.
Gambrell N J. Screening St. Augustinegrass [Stenotaphrum secundatum (Walt.) Kuntze] for USDA zone 7[D]. Clemson: Clemson University, 2014.
羅? 耀, 席嘉賓, 譚筱弘, 等. 9種暖季型草坪草耐陰性綜合評價及其指標的篩選[J]. 草業學報, 2013, 22(5): 239-247.
Caminos T S, Estévez S J. Effects of shade on the persistence of cool-season grasses to form turfgrass[J]. Agricultura Técnica, 2007, 67(4): 372-383.
Okeyo D O, Fry J D, Bremer D J, et al. Stolon growth and tilling of experimental Zoysiagrasses in shade[J]. HortScience, 2011, 46(10): 1418-1422.
張? 靜, 廖? 麗, 白昌軍, 等.竹節草對NaCl脅迫臨界濃度的初步研究[J]. 草地學報, 2014, 22(3): 661-664.
廖? 麗, 蔣仁嬌, 劉建秀, 等. 竹節草種質資源抗寒性初步評價研究[J]. 熱帶作物學報, 2016, 37(2): 234-240.
Trenholm L E, Nagata R T. Shade tolerance of St. Augustinegrass cultivar[J]. Hort Technology, 2005, 15(2): 267-272.
Cai X, Trenholm L E, Kruse J, et al. Response of “Captiva” St. Augustinegrass to shade and potassium[J]. HortScience, 2011, 46(10): 1400-1403.
陳? 斌, 李洪瑤, 劉筱瑋, 等. 不同光照強度對新娘草葉片形態建成及超微結構的影響[J]. 草業科學, 2019, 28(7): 175-185.
Jahns P, Miehe B. Kinetic correlation of recovery from photo-inhibition and zeaxanthin epoxidation[J]. Planta, 1996,198(2): 202-210.
德力格爾, 李媛媛, 張天俊. 遮陰對成坪期草地早熟禾和紫羊茅生理指標的影響[J]. 草原與草坪, 2015, 35(5): 50-54, 59.
Trenholm L E. Effect of shade on performance of St. Augustine cultivars[C]. Meeting of the Florida State Horticultural Society. 2008.
尹淑霞. 幾種冷季型草坪草的耐蔭性研究[D]. 蘭州: 甘肅農業大學, 2000.
劉怡凡, 張蓓蓓, 王? 荷, 等. 光照強度對黑麥草葉片可溶性蛋白含量及生長發育的影響[J]. 寶雞文理學院學報(自然科學版), 2018, 38(4): 49-53, 66.
張麗平, 周亞圣, 劉君成, 等. 遮蔭對刈割互花米草生物量及滲透調節物質的影響[J]. 西南林業大學學報(自然科學), 2019, 39(4): 142–148.
李志剛, 侯扶江, 安? 淵. 不同光照強度對三種牧草生長發育的影響[J]. 中國草地學報, 2009, 31(3): 55-61.
猜你喜歡
中國綠色畫報(2016年11期)2017-02-18 15:21:32
中州學刊(2016年11期)2017-01-06 02:17:50
中國綠色畫報(2016年7期)2016-12-26 16:37:18
藝術科技(2016年10期)2016-12-14 20:02:46
文藝生活·下旬刊(2016年11期)2016-12-12 08:14:33
商情(2016年40期)2016-11-28 11:50:53
藝術科技(2016年9期)2016-11-18 18:35:07
人間(2016年27期)2016-11-11 15:56:02
中國科技博覽(2016年13期)2016-07-13 02:25:11
科技視界(2016年7期)2016-04-01 12:24:45
