深海貝萊斯芽孢桿菌DH82的抑菌活性物質初步分離純化及其抑菌譜檢測
2020-02-28 09:13:10王青華孫曉暉萬婧倞黃仕新徐長安
應用海洋學學報 2020年1期
王青華,唐 旭,孫曉暉,萬婧倞,黃仕新,徐長安
(自然資源部第三海洋研究所、自然資源部海洋生物資源綜合利用工程技術研究中心,福建 廈門 361005)
細菌、真菌性病害是動植物養殖、種植產業的重要災害,嚴重制約了動植物養殖、種植生產的持續發展。迄今,傳統化學藥物和抗生素的應用仍然是控制病害的主要措施,但長期、大劑量、大面積的不正當使用,造成病原菌耐藥、藥物殘留等問題日漸顯現,對人類的健康造成極大的安全隱患[1-2]。隨著人民生活水平提高,人民對無公害食品的要求日益高漲,開發和利用生防菌防治水產病害的方法受到了廣泛的關注。
深海有著極其豐富的物種多樣性,其多種復雜獨特的生境,包括熱液口、冷泉等,造就了有著獨特功能的海洋微生物,是國家重要的戰略儲備資源,同時具有巨大的研發空間和應用開發潛力[3-4]。芽孢桿菌是廣泛存在于自然界之中的非致病性細菌,繁殖能力和環境耐受性強,能產生多種抑菌物質,具有巨大的開發利用價值[5-6]。相對于陸地芽孢桿菌,深海芽孢桿菌生活在非常特殊的高溫、高壓、高污染環境里,決定了其次級代謝產物更豐富、結構新穎和具有特異的生物活性,同時賦予深海芽孢桿菌產生具有新型抑菌活性物質的能力[7]。
據報道,研究者已從深海芽孢桿菌中獲得了不少生物活性物質。張少博等(2013)從深海獨島枝芽孢桿菌(Virgibacillusdokdonensis)A493分離到新的氨基糖苷類抑菌活性物質,其抑菌譜有特異性[8];呂倩等(2014)從南海深海里篩選到Bacillusmethylotrophicus,該菌產抗真菌活性的bacillomycin Lc類脂肽[9];Li等(2016)從深海枯草芽孢桿菌(B.subtilis)B5中提取到一種新的macrolactin A衍生物,該成分能防控茶病原真菌包括茶擬盤多毛孢(Pestalotipsistheae)和膠孢炭疽菌(Colletotrichumqloeosporioides)[10];龍彬等(2014)從深海解淀粉芽孢桿菌(B.amyloliquefacien)分離出12個化合物,其中化合物9(4-羥基脯氨酸-亮氨酸)對番木瓜炭疽菌(C.papayae)和香蕉黑星菌(Phyllosticamusarum)有中等抑菌活性[11]。作者從深海微生物中篩選到一株貝萊斯芽孢桿菌DH82(BacillusvelezensisDH82),前期研究發現其產的抗菌蛋白有較好熱穩定性,不受木瓜蛋白酶、蛋白酶K、胃蛋白酶及胰蛋白酶影響。
貝萊斯芽孢桿菌是一類非常重要的有生防潛力的細菌[12]。近年來,人們把貝萊斯芽孢桿菌對陸地動、植物病原細菌類或病原真菌防治分別進行研究,很少把病原細菌和病原真菌整合在一起進行防治研究,并且針對貝萊斯芽孢桿菌產生的抑菌物質對水產病原菌的防治效果的報道更是寥寥無幾[13-15]。本研究就貝萊斯芽孢桿菌DH82產生抗菌蛋白進行初步分離及其對水產病原細菌和植物病害真菌的拮抗作用進行了研究,以期為抗菌蛋白進一步開發應用奠定基礎。
1 材料及方法
1.1 材料
1.1.1 菌株 出發菌:出發菌DH82分離自蛟龍號第150潛次中西太平洋雅浦海溝6 000 m深的海水樣品,經16S rDNA(GenBank登錄號:MK203035)和gyrB同源性序列對比分析,結合其形態學和生理生化特性,鑒定出發菌DH82為貝萊斯芽胞桿菌BacillusvelezensisDH82,在改良LB(Luria Broth)斜面固體培養基4 ℃保存。
指示病原細菌:遲緩愛德華氏菌(Edwardsiellatarda)、副溶血弧菌(Vibrioparahaemolyticus)、溶藻弧菌(V.alginolyticus)由廈門大學生物實驗室提供,在LB斜面固體培養基4 ℃保存;坎氏弧菌(V.campbellii)、嗜水氣單胞菌(Aeromonashydrophila)、腸炎沙門氏菌(Salmonellaenteritidis)由中國海洋微生物菌種保藏管理中心MCCC 提供,在LB斜面固體培養基4 ℃保存;哈維氏弧菌(V.harveyi)由中國海洋微生物菌種保藏管理中心MCCC提供,在改良Zobell 2216E斜面固體培養基4 ℃保存;大腸桿菌(Escherichiacoli)、銅綠假單胞菌(Pseudomonasaeruginosa)、金黃色葡萄球菌(Staphylococcusaureus)由本實驗室提供,在LB斜面固體培養基4 ℃保存。無乳鏈球菌(Streptococcusagalactiae)由廣東微生物菌種保藏中心提供,在TSB(Tryptic Soy Broth)斜面固體培養基4 ℃保存。
指示病原真菌:層生鐮刀菌(Fusariumproliferatum)、笄霉屬(Choanephora)和水賊鐮刀菌(Fusariumequisetum)分離自感染植物病害的蔬菜,由本實驗室提供,在PD(Potato Dextrose)斜面固體培養基4 ℃保存。灰葡萄孢菌(Botryiscinerea)由中國典型培養物保藏中心提供,在PD斜面固體培養基4 ℃保存。
1.1.2 培養基 LB培養基(1 000 mL):蛋白胨10 g、酵母粉5 g、氯化鈉10 g、pH=7.3,改良Zobell 2216E培養基(1 000 mL):蛋白胨5 g、酵母提取物1 g、天然過濾海水∶純水=2∶1、pH=7.6,用于病原細菌培養;改良LB 培養基(1 000 mL):胰蛋白胨10 g、酵母粉5 g、氯化鈉5 g、pH=7.3,用于菌株DH82種子液培養及發酵;PD培養基(1 000 mL):馬鈴薯200 g、蔗糖20 g、pH=6.7,用于病原真菌的活化及培養。
1.2 方法
1.2.1 抗菌蛋白的初步分離純化 ①菌株DH82的發酵上清液制備。將菌株DH82活化菌液接種至500 mL培養基(1 L 錐形瓶),接種量5 mL,37 ℃,180 r/min,24 h后,發酵液離心(4 ℃,10 000 r/min,10 min)棄菌體,過0.22 μm微孔濾膜,得到發酵上清液。② 硫酸銨的分級沉淀。取500 mL發酵上清液,向其中緩慢加入固體硫酸銨至飽和度分別為30%、40%、50%、60%、70%、80%、90%,4 ℃,過夜沉淀,4 ℃,10 000 r/min,離心20 min,收集沉淀。用純水復溶沉淀,于3 000 Da透析袋中除鹽,透析18 h,以遲緩愛德華氏菌為指示菌,用改良瓊脂擴散法檢測各組分抑菌活性[16-17]。③ Sephadex G-75柱層析。Sephadex G-75用超純水浸泡24 h,充分溶脹后,裝柱(1.6 cm×50.0 cm);用超純水平衡24 h,上樣,上樣量為5 mL,超純水洗脫,流速2 mL/min,核酸蛋白檢測儀(HD-5電腦紫外檢測儀購上海瀘西分析儀器廠有限公司)280 nm檢測,分管收集,每管收集8 mL,以遲緩愛德華氏菌為指示菌,檢測抑菌活性,收集具有抑菌活性的洗脫部分。
1.2.2 抗菌蛋白抑菌譜測定 ①改良瓊脂擴散法測定。取1.5 mL新鮮的指示病原細菌液(在18 mm×180 mm試管于37 ℃培養12 h)接種于150 mL的LB/改良Zobell 2216E(46 ℃)固體培養基中,迅速搖勻倒入培養皿中(9 cm 規格),凝固后用無菌打孔器(直徑8.58 mm)在抑菌板上打孔,并用滅過菌的牙簽挑出培養基塊。每孔加入50 μL待檢測液,在超凈臺中吹干后置于37 ℃,培養12 h后觀察是否有抑菌圈,測量抑菌圈的直徑。②濾紙片法測定[18]。濾紙裁剪成直徑為8.58 mm的圓形濾紙片,121 ℃高壓滅菌,干燥備用。用無菌水沖洗產孢子的病原真菌,把孢子濃度配置成1×104CFU/mL,然后按10%接種量接到冷卻至46 ℃的PD固體培養基上,迅速混勻倒平板。用無菌鑷子取無菌的濾紙片放置在抑菌板上,用移液槍在每個濾紙片上滴加待檢測液,設置3個重復。對照組只加無菌水,28 ℃倒置培養至對照組的菌落長滿平板,測量各抑菌活性物質的抑菌圈直徑。③分離純化蛋白抑菌譜檢測。純化后的抗菌蛋白液采用瓊脂擴散法和濾紙片法對上述指示病原菌進行抑菌活性檢測,檢測方法分別參照上文。
1.2.3 最小抑菌濃度(MIC)測定 ①菌懸液的制備。病原細菌的菌懸液制備:將病原細菌接種在LB固體平板37 ℃,12~24 h;用接種環刮取少量菌體,放入生理鹽水中,漩渦震蕩混勻,用麥氏比濁管(購自廣東環凱微生物科技有限公司)計算細菌的濃度,分別稀釋至濃度為2×104CFU/mL。病原真菌的菌懸液制備:將病原真菌接種在PD固體平板28 ℃,4~6 d;用沾無菌水的接種環刮取少量孢子,放入無菌水,漩渦震蕩混勻,血球計數板計數,稀釋至濃度為2×104CFU/mL。②蛋白質含量測定。蛋白質含量采用改良型BCA(Bicinchoninic acid)法測定。改良型BCA法蛋白質濃度測定試劑盒購自生工生物工程(上海)股份有限公司。配置相應濃度的標準蛋白質溶液,取5 μL相應濃度的標準蛋白質溶液或樣品加5 μL已配置好溶液F(烷基化試劑+Tris緩沖液),混勻,于37 ℃,30 min后加200 μL BCA工作液,混勻,于37 ℃,30 min,冷卻至室溫后,在562 nm處測定吸光度,做標準曲線,在標準曲線上計算出各蛋白質含量。③最小抑菌濃度(MIC)測定。抗菌蛋白對病原菌作用效果通過進行最小抑菌濃度的測定獲得[19]。在96孔板每孔加入培養指示菌的空白培養基100 μL,在A/B/C三排的第一孔加配好待檢測液100 μL,然后對待檢測液進行二倍稀釋。即第一孔加入待測液后,用移液槍充分吹打三次,使待測液與培養基充分混勻,吸取100 μL加入第二孔,充分吹打混勻,照此重復直至最后一孔,吸取100 μL棄去;再在每一孔加入稀釋好的菌懸液100 μL;在同一塊板上的G排做好陰性對照(僅加200 μL空白肉湯)和在H排上做一排陽性對照(加100 μL菌液100 μL空白培養基),96孔板放入恒溫培養箱,病原細菌:37 ℃,18~24 h;病原真菌:28 ℃,48~72 h,觀察抑菌效果。
2 結果與分析
2.1 抗菌粗蛋白沉淀的最佳硫酸銨飽和度
不同硫酸銨飽和度提取的粗蛋白對遲緩愛德華氏菌的抑菌效果。如圖1所示,硫酸銨飽和度為70%時,所提取的粗蛋白抑菌活性最高。隨著硫酸銨飽和度不斷增加,所提取的蛋白抑菌活性沒有繼續增加,說明硫酸銨飽和度為70%時,抗菌粗蛋白幾乎已經沉淀完全。因此,提取抗菌粗蛋白的最佳硫酸銨飽和度為70%。
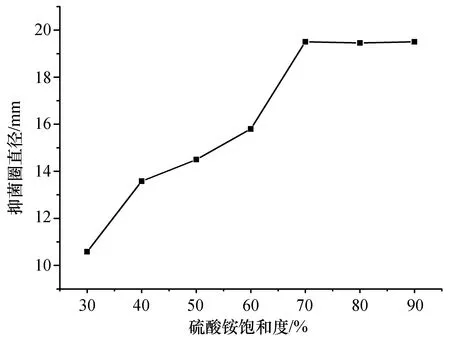
圖1 不同硫酸銨飽和度提取的粗蛋白抑菌活性
2.2 抗菌粗蛋白Sephadex G-75柱層析
將菌株DH82的發酵上清液經硫酸銨飽和度70%鹽析,提取的抗菌粗蛋白液經Sephadex G-75柱層析分離,從圖2看出,抗菌粗蛋白被分離出2個峰。峰Ⅰ所對應的收集管T8、T9,其管液檢測均有抑菌活性,該峰蛋白命名為PrI;峰Ⅱ所對應的收集管為T12、T13、T14,其管液檢測均有抑菌活性,該峰蛋白命名為PrII。表明峰蛋白PrI和峰蛋白PrII都是有抑菌活性的目標蛋白。
2.3 抗菌蛋白抑菌譜
2.3.1 峰蛋白PrI和PrII對水產病原細菌的拮抗作用 將峰蛋白PrI和PrII對上述常見指示水產病原細菌進行拮抗實驗,得到其抑菌作用效果。如表1所示,峰蛋白PrI和PrII分別對遲緩愛德華氏菌、嗜水氣單胞菌、坎氏弧菌、溶藻弧菌、無乳鏈球菌和金黃色葡萄球菌有較好抑菌活性,其中峰蛋白PrII對哈維氏弧菌也具有較好抑菌活性;峰蛋白PrI和PrII分別對副溶血弧菌、銅綠假單胞菌、腸炎沙門氏菌、大腸桿菌作用未檢出,其中峰蛋白PrI對哈維氏弧菌作用也未檢出;表明峰蛋白PrI和PrII對某些常見水產病原細菌具有較好抑菌活性。
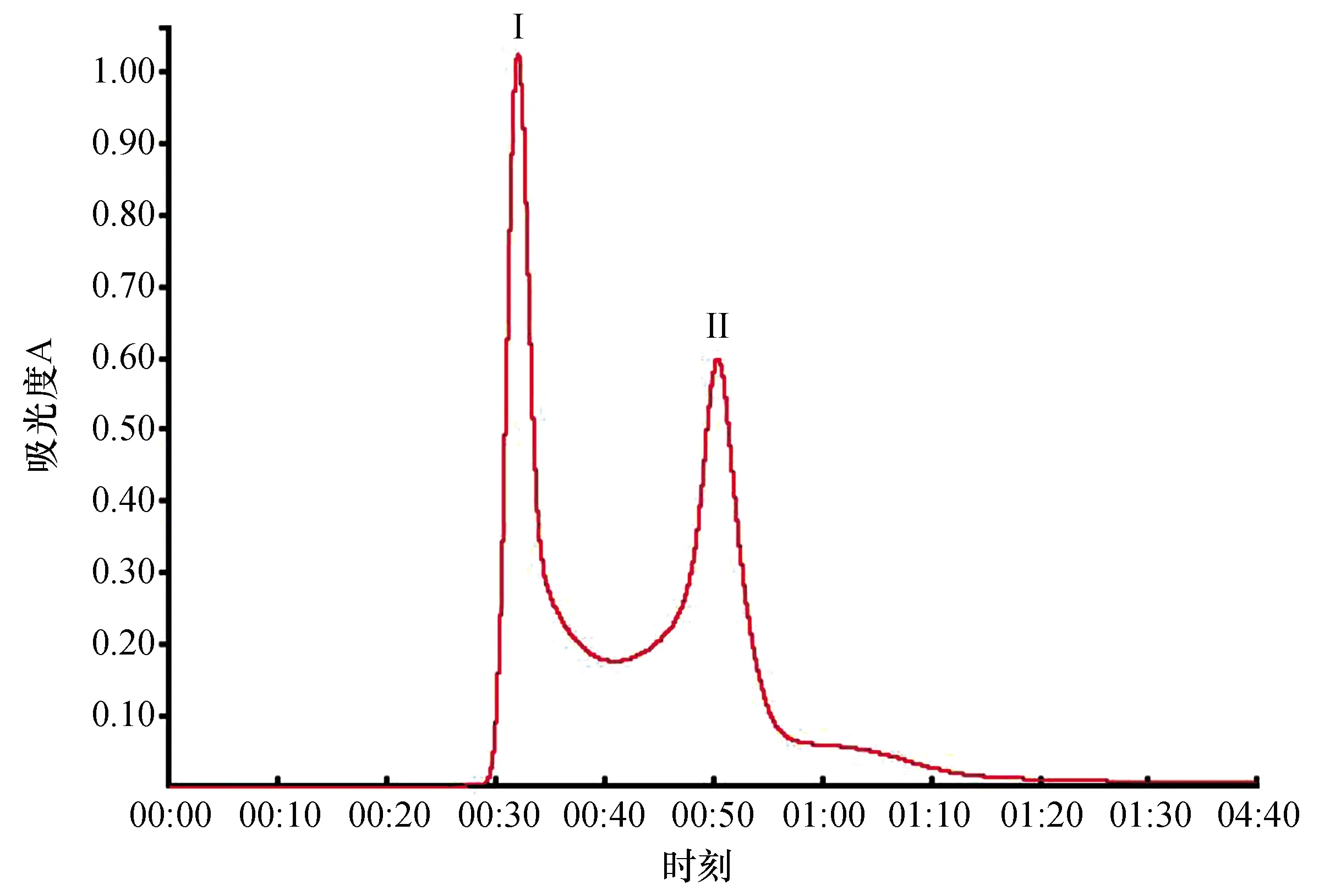
圖2 抗菌粗蛋白Sephadex G-75層析圖
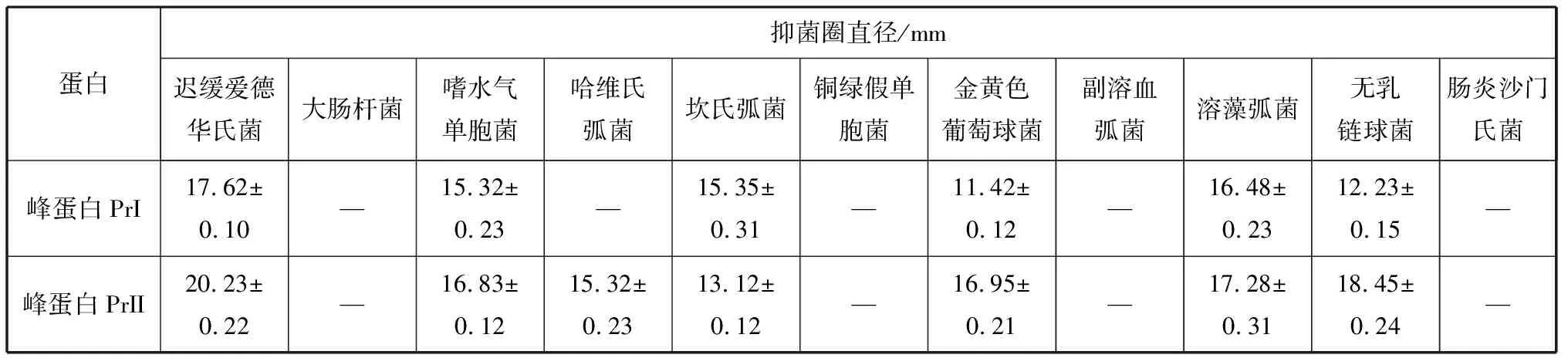
表1 峰蛋白PrI和PrII對11種水產病原細菌的拮抗作用
2.3.2 峰蛋白PrI和PrII對植物病原真菌的拮抗作用 將峰蛋白PrI和PrII對上述4種指示植物病原真菌進行拮抗實驗,其抑菌效果由表2所示,峰蛋白PrI和PrII分別對層生鐮刀菌、笄霉屬和灰葡萄孢菌抑菌作用未檢出,但峰蛋白PrII對水賊鐮刀菌有較好抑菌活性,峰蛋白PrI對水賊鐮刀菌抑菌作用未檢出。
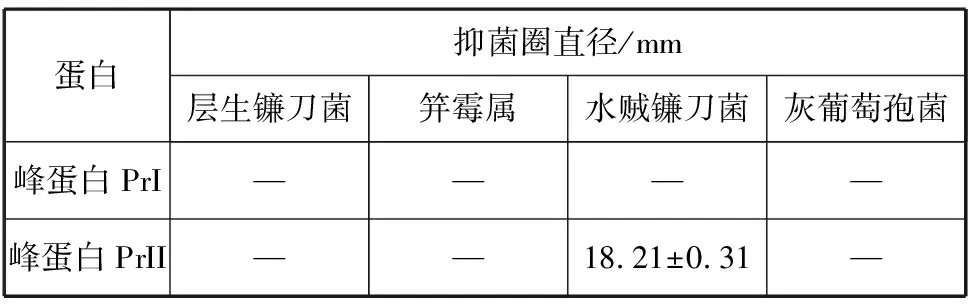
表2 峰蛋白PrI 和PrII對4種植物病原真菌的拮抗作用
2.4 抗菌蛋白對病原菌的最低抑菌濃度(MIC)
2.4.1 標準曲線的繪制 使用改良BCA法進行蛋白濃度測定,以標準牛血清蛋白溶液的蛋白濃度(μg/mL)作為橫坐標,以其吸光度為縱坐標,繪出標準曲線,如圖3所示,得出線性回歸方程y=0.000 2x-0.001 5,相關系數是0.999 3。
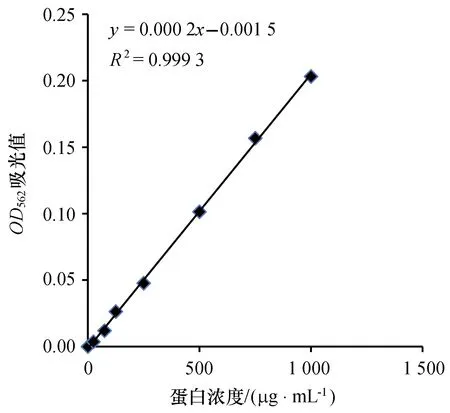
圖3 牛血清蛋白標準曲線
2.4.2 峰蛋白PrI和PrII對水產病原細菌的最低抑菌濃度(MIC) 峰蛋白PrI和PrII分別對7種常見水產病原細菌的最低抑菌濃度值如表3所示。可以看出,峰蛋白PrI對遲緩愛德華氏菌和溶藻弧菌的MIC最低,對金黃色葡萄球菌的MIC最高。峰蛋白PrII對遲緩愛德華氏菌的MIC最低,對嗜水氣單胞菌、溶藻弧菌、無乳鏈球菌和金黃色葡萄球菌的MIC都較低,對哈維氏弧菌的MIC最高。

表3 峰蛋白PrI和PrII拮抗7種水產病原細菌的MIC測定
2.4.3 峰蛋白PrII對植物病原真菌的最低抑菌濃度(MIC) 峰蛋白PrII對水賊鐮刀菌的MIC為7.16 μg/mL,表明峰蛋白PrII在較低MIC就能抑制水賊鐮刀菌。
2.5 討論
貝萊斯芽孢桿菌是具有廣譜抑菌活性的拮抗細菌,其主要通過產生抗菌物質包括抗菌蛋白和脂肽類抗生素來發揮抑菌活性[20-21]。貝萊斯芽孢桿菌DH82沉淀抗菌粗蛋白的硫酸銨最佳飽和度是70%,與李姝江等(2013)鹽析抗菌粗蛋白的所需硫酸銨最佳飽和度50%有一定差異[20],與黃銘慧(2017)鹽析的抗菌粗蛋白所需硫酸銨最佳飽和度70%一致[22]。
貝萊斯芽孢桿菌DH82發酵上清液經Sephadex G-75柱層析分離得到的兩個峰蛋白PrI和PrII對常見水產病原細菌抑菌譜都相對較廣,且對遲緩愛德華氏菌和溶藻弧菌的最小抑菌濃度(MIC)都較低,可為針對這兩種病原細菌進一步開發殺菌劑奠定基礎。峰蛋白PrII對引起植物病害的水賊鐮刀菌有較好的抑菌活性,對灰葡萄孢菌、笄霉屬、層生鐮刀菌均無抑菌活性,這與一些文獻報道不一致,如Kaki等(2013)從金盞花根際分離的貝萊斯芽孢桿菌分泌的脂肽對尖孢鐮刀菌(F.oxysporum)和葡萄孢菌有抑制作用[21];另外,有研究者報道了貝萊斯芽孢桿菌對鐮刀菌屬有拮抗作用,并用于禾谷鐮刀菌(F.graminearum)引起的小麥赤霉病和對尖孢鐮刀菌引起的植物枯萎病的防控[23-25],但在對水賊鐮刀菌引起的枯萎病防治還未有報道,這些差異有待以后進一步研究探討。峰蛋白PrII對嗜水氣單胞菌、無乳鏈球菌、金黃色葡萄球菌和水賊鐮刀菌的MIC都較低(7.16μg/mL),這在常見水產病原細菌和個別植物真菌病害防治上有較好的利用價值。另外,隨著生物質譜技術和分離技術快速發展[26-27],可用這些技術來進一步分離和純化該抗菌蛋白,為克隆表達相關抑菌功能基因奠定基礎,并進而利用基因工程技術規模化制備生產該抗菌蛋白,為生產綠色水產品和農產品服務。
3 結論
本研究結果表明,貝萊斯芽孢桿菌DH82的抗菌粗蛋白中分離出兩個具有抑菌活性的峰蛋白PrI和PrII,峰蛋白對常見水產病原細菌有較廣譜的抑菌活性,對遲緩愛德華氏菌和溶藻弧菌的最小抑菌濃度(MIC)都較低,其中峰蛋白PrII對植物病害真菌水賊鐮刀菌也有較好的抑菌活性,且峰蛋白PrII對嗜水氣單胞菌、無乳鏈球菌、金黃色葡萄球菌和水賊鐮刀菌的MIC都較低。因此,貝萊斯芽孢桿菌DH82作為生防菌在常見水產細菌性病害的防治及由水賊鐮刀菌感染引起的植物枯萎病防治上有較好的開發利用價值。
