銅或鎘脅迫對海洋青鳉生理生化指標及產卵量的影響
2020-03-04 03:15:56孫金輝徐永健謝尚端
水生生物學報 2020年1期
孫金輝 潘 霞 徐永健 顏 鑫 謝尚端
(1. 天津農學院水產學院天津市水產生態及養殖重點實驗室,天津 300384; 2. 寧波大學海洋學院, 寧波 315211)
重金屬是常見的海洋污染物, 主要由人類生產生活產生并被攜帶至海中[1,2]。由于重金屬的毒性、持久性以及生物累積的特性, 會對人類、生物和自然生態系統構成嚴重威脅[3,4]。如銅能影響生物體內的酶系統[5], 會導致生理受阻, 發育停滯[6],急性銅中毒會刺激胃腸道黏膜, 慢性中毒則會引起肝硬化[7]; 鎘為“五毒之首”, 有致畸、致癌特性, 鎘污染會使動物酶系統受到抑制[8,9], 長期受鎘影響會損傷腎、肺、肝[10]等, 會使卵母細胞成活率、受精率降低, 最終導致不孕不育[11]。
海洋青鳉(Oryzias melastigma)是一種新興的海水模式生物, 對水質和環境變化極敏感, 是毒理學上至關重要的監測物種[12,13]。相比斑馬魚(Barchydanio rerio)和日本青鳉(Oryzias latipes), 其在培養條件、生理、行為及分子信息等方面具有明顯優勢[14,15], 是海洋環境毒理學上有應用潛力的重要生物監測種類[16—18], 已被國際生命科學學會(ILSI)認定為毒理學研究的重要模式生物[19], 也是國際標準組織(ISO)推薦的毒性實驗標準用魚之一[20]。研究海洋青鳉能夠擴充海洋生態毒理模式的生物領域, 豐富不同污染條件下海洋青鳉的反應情況, 為日后海洋污染生態學和模式生物的研究發展打好基礎[21]。
利用日本青鳉監測淡水水質的相關方法已較為成熟, 通過識別魚的行為判斷水質是否發生變化[22,23]。應用該方法所建立的純生物傳感技術, 現已能精確地判斷水質的污染狀態, 并可根據行為差異進行污染程度劃分[23]。而利用海洋青鳉進行海水水質監測的相關方法和技術較日本青鳉比還有很大的差距, 目前涉及海洋青鳉的研究, 主要以幼魚較多[24], 成魚較少; 高濃度重金屬脅迫較多, 低濃度脅迫較少。
本文以海洋青鳉成魚作為實驗生物, 分析不同濃度銅、鎘脅迫下魚體的一些生理生化指標變化,以乳酸(LA)含量、乳酸脫氫酶(LDH)活性來反應脅迫對魚體體能消耗的影響[25], 以睪酮(T)、促卵泡激素(FSH)和促黃體生成素(LH)含量和產卵量來反應脅迫對魚體生殖能力的影響。把環境脅迫-能量消耗-生殖能力下降等現象聯系起來, 反映一些化學監測、物理監測甚至常規生物監測都難以發現的低濃度毒物對生物有機體的慢性和深度影響。
1 材料與方法
1.1 實驗動物
海洋青鳉魚卵由廈門大學王克堅老師實驗室惠贈, 已于本實驗室成功擴繁五代以上。擴繁條件: 水溫: 28—30℃, 鹽度: 28—30, 光照: 8000 lx, 光照周期: L14h∶D10h, 餌料: 新孵化鹵蟲幼體。試驗用的青鳉成魚為4—5月齡, 是正常繁殖群體。雄魚體全長為(4.0±0.25) cm、體重(0.52±0.13) g, 雌魚體全長為(3.5±0.19) cm、體重(0.45±0.11) g。
1.2 實驗設計
以海水水質標準為參考, 依據文獻[26]報道設置銅和鎘的濃度梯度。銅(CuSO4·5H2O(AR)配制)設3個梯度: C1: 1/10的96hLC50(0.017 mg/L), C2:1/2的96hLC50(0.087 mg/L), C3: 96hLC50(0.174 mg/L); 鎘[CdCl2·2.5H2O (AR)]3個梯度: G1: 1/10的96hLC50(0.650 mg/L), G2: 1/2的96hLC50(3.249 mg/L), G3: 96hLC50(6.497 mg/L)。每個處理3個重復,并設置一個空白組作對照(C0/G0)。每組放養青鳉成魚20尾(雌雄各半), 養殖水體30 L, 養殖海水用自來水+海水晶(廣州益爾生物工程有限公司)配制, 條件同擴繁條件。每日清理養殖缸中的排泄物并更換約20%同濃度現配海水。分別于4d和15d, 在吸污換水時收集沉底的卵, 計數后統計各處理的產卵情況, 計算單尾雌魚的平均產卵量(粒)。
試驗共持續15d(28℃時青鳉一個繁殖周期)。分別于4d和15d取樣, 測定LA、LDH、T、FSH和LH。每次從每組中隨機撈取雌雄各3尾魚, 用蒸餾水沖洗2—3遍, 加入5 mL 0.9%生理鹽水并勻漿,3000×g低溫(4℃)離心10min后取上清, 然后根據檢測試劑盒(購于上海源葉生物技術有限公司)說明書的操作步驟測定LA、LDH、T、LH和FSH等指標。
1.3 數據分析方法
實驗結果利用Microsoft Excel和SAS 8.01 軟件進行統計分析, 采用單因素方差分析(ANOVA), 并選取Duncan法對實驗組進行比較,P<0.05 表示有顯著性差異。
2 結果
2.1 銅或鎘脅迫對海洋青鳉成魚中LA含量的影響
如圖1所示, 在試驗過程中, 對照組前后LA含量無顯著性差異, 雌雄個體間也無顯著性差異(P>0.05)。
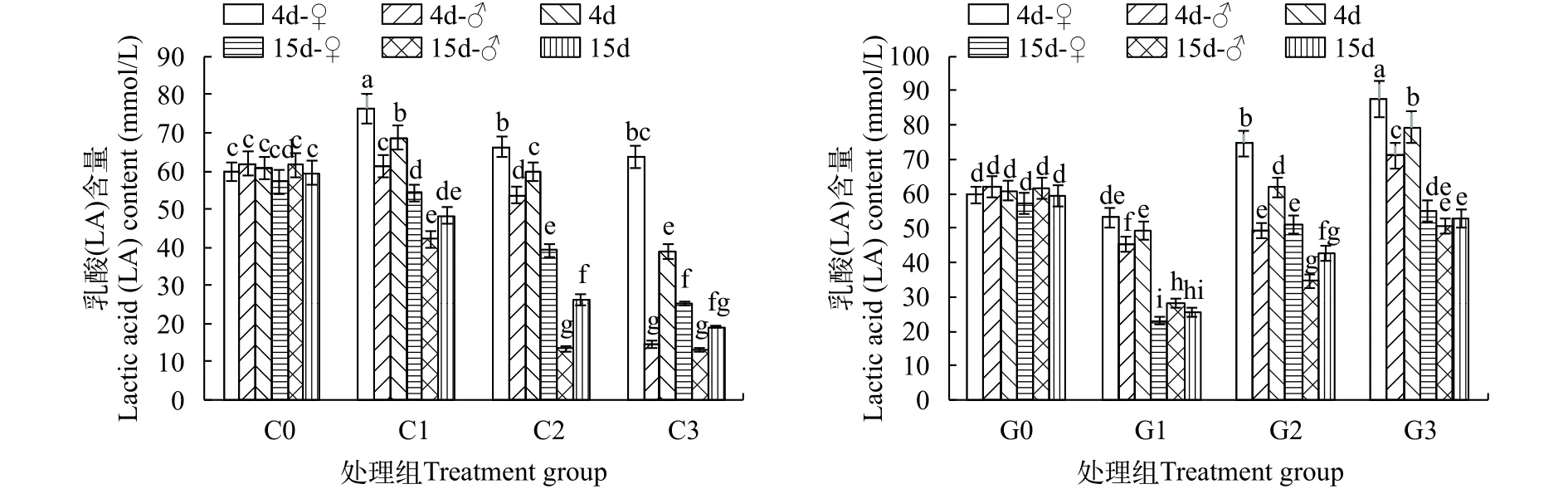
圖1 不同濃度銅或鎘脅迫下海洋青鳉的乳酸(LA)含量變化Fig. 1 The lactic acid contents of Oryzias melastigma under different concentrations of cadmium or copper
銅脅迫4d, 不同濃度處理組的LA含量出現了較大變化, 總體上隨銅濃度升高而LA含量下降, 與C0相比, C1顯著上升、C2持平、C3顯著下降;C1上升主要是雌魚的LA大幅升高所致, 該濃度對雄魚影響不大; C2銅濃度對雌魚和雄魚有相反的影響, 兩者平均后與對照無差異; C3銅濃度對兩性都有影響, 雌魚影響類似于C2, 而雄魚的LA含量非常低, 僅為14.4 mmol/L, 遠低于對照及其他處理。至15d, 魚體中的LA含量仍然是隨著銅濃度的升高而顯著下降。所有處理都低于對照組, 其中, C1的雌魚近似對照, 雄魚有較低值; C2兩者都低于C1和對照, 雄魚大大低于雌魚; C3類似于C2, 其中雄魚與C2相近, 雌魚低于C2, 因此整體不如C2 (P<0.05)。
鎘脅迫4d, 不同濃度處理青鳉魚體內的LA變化與銅脅迫時有所不同。整體上, LA含量隨鎘濃度的升高而升高, 隨脅迫時間的延長而下降。G1處理僅比G0略低, G2高于G0, G3最高, 兩兩組間差異顯著(P<0.05)。對性別的影響, 雄性的LA含量都低于雌性, 差異顯著(P<0.05), 此外G1與G2中雄魚的LA相似, 并低于G3。至15d, 各處理的LA含量進一步下降, 脅迫組的LA含量都比對照組低(P<0.05)。對性別的影響, 除G1外其余處理中雄魚要低于雌魚(圖1)。
2.2 銅或鎘脅迫對海洋青鳉成魚中LDH活性的影響
在實驗過程中, 對照組青鳉體內的LDH活性無顯著性差異(P>0.05)。銅脅迫4d, C2低于對照, 而C1與C3組高于對照, 后兩者間差異不顯著; 其中,C1雌魚的LDH活性較雄魚強, C2相似, 而C3則相反, 雄魚的LDH活性強于雌魚。至15d, 各處理的LDH活性進一步下降, C1處理主要是雄魚的下降造成, C2是雌魚下降明顯, 而C3兩性同時都下降。
鎘脅迫4d (圖2), 與對照比較, G1和G3的LDH活性略有下降, G2下降最多。其中, G1和G2處理, 鎘對雌雄魚的影響相近, 而G3中雄魚LDH活性下降更顯著。至15d, 各處理的LDH活性都有較大的升高。G1升高最為明顯, 雌魚比雄魚升高更多;G2恢復到與對照相似水平, G3比對照稍高一點(P<0.05)。

圖2 不同濃度銅或鎘脅迫下海洋青鳉體內乳酸脫氫酶(LDH)活性的變化Fig. 2 Lactate dehydrogenase activities of Oryzias melastigma under different concentrations of cadmium or copper
2.3 銅或鎘脅迫對海洋青鳉成魚中T含量的影響
在試驗過程中, 對照組前后魚體中的T含量無顯著性差異(P>0.05)。銅脅迫4d, 各處理的T含量都有所上升, 升幅相近。其中C1中雌魚較雄魚上升多, 而C2和C3卻相反。至15d, 各處理的T含量都下降, 其中C3下降最為明顯。兩性間的差異類似于第4天, 其中比較特別的是C2處理, 雄魚保持著高于對照水平的T含量, 但雌魚下降很大; C3兩性都下降,也是雌魚下降更大。
在鎘脅迫時, T含量隨鎘濃度的升高而升高, 其中G1與對照相近, G2和G3高于對照(P<0.05)。脅迫4d時, 3個處理雌魚的T含量都升高了, 且升幅大于雄魚。至15d, G1處理與對照相近, G2和G3處理都高于對照, 雄魚的含量高于雌魚(圖3)。
2.4 銅或鎘脅迫對海洋青鳉成魚中FSH含量的影響
在試驗期間, 對照處理中魚體的FSH含量前后也無顯著性差異(P>0.05)。銅脅迫4d, 僅C2略低于對照, C1和C3與對照無差異。對性別的影響, 也僅C3, 雌魚略有上升, 而雄魚略有下降。至15d, C1比第4天略有下降, 但C2和C3的FSH都有大的降幅,C3的降幅最大, 與C2間有差異(P<0.05)。各組中兩性間的差異不大。鎘脅迫4d, 3個處理中FSH都略有下降, 其中G2降幅最大。在所有處理中, 雄魚的降幅都大于雌魚。至15d, 各處理中FSH含量都上升了。其中, G1和G3已接近于對照, 而G2略低于對照。雌雄間基本無差別(圖4)。
2.5 銅或鎘脅迫對海洋青鳉成魚中LH含量的影響
在試驗期間, 對照組中魚體的LH含量前后無顯著性差異(P>0.05, 圖5)。銅脅迫4d, 各組與對照相似, 性別間也無差異。至15d, 各組的LH含量都降低了, 且隨銅濃度升高而下降越明顯(P<0.05), 但性別間仍無差異。鎘脅迫4d, G1略有下降, G2降幅較大, 而G3與對照相似。所有處理雄魚的降幅都大于雌魚。至15d, 各處理中LH含量都有所恢復。G1和G3 都接近于對照, 而G2略低于對照。

圖3 不同濃度銅或鎘脅迫下海洋青鳉魚體內睪酮(T)含量的變化Fig. 3 Testosterone contents of Oryzias melastigma under different concentrations of cadmium or copper
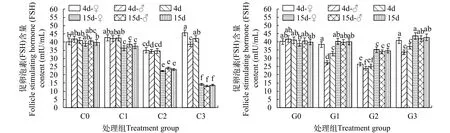
圖4 不同濃度銅或鎘脅迫下海洋青鳉體內促卵泡素(FSH)含量的變化Fig. 4 Follicle stimulating hormone contents of Oryzias melastigma under different concentrations of cadmium or copper
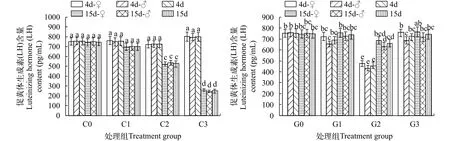
圖5 不同濃度銅或鎘脅迫下海洋青鳉體內促黃體生成素(LH)含量的變化Fig. 5 Luteinizing hormone contents of Oryzias melastigma under different concentrations of cadmium or copper
2.6 銅或鎘脅迫對海洋青鳉成魚產卵量的影響
在試驗過程中, 所有處理都未出現海洋青鳉死亡現象。短時間(4d)的銅脅迫對產卵量基本沒有影響, 但隨著脅迫時間的延長(15d)和脅迫濃度的升高, 海洋青鳉產卵量下降非常明顯(圖6)。而短時(4d)的鎘脅迫, 僅高濃度(G2)處理的產卵量有所下降(P<0.05); 隨脅迫時間的延長(15d)和脅迫濃度的升高, 各處理中海洋青鳉的產卵量逐漸相近(P>0.05), 并與第4天相比不存在顯著差異(P>0.05,圖6)。
2.7 各指標間相互關系分析
在海洋青鳉魚銅或鎘脅迫下各指標間的相關性分析見圖7。銅或鎘脅迫對青鳉的LA含量影響是不同的, 隨銅濃度的升高而降低, 隨鎘濃度的升高而升高(圖7a、7b)。隨銅或鎘脅迫時間的延長,雌雄魚體內LA含量均降低。在雌魚受銅或鎘脅迫時, 體內T含量與LA含量呈正相關關系, 銅脅迫的相關性系數大于鎘脅迫(圖7c、7d), T與FSH/LH間也有良好的正相關關系。此外, 短期(4d)內銅脅迫對雌魚產卵量沒有影響, 但長期(15d)脅迫明顯影響產卵量, 并隨濃度升高而生殖能力下降(圖7e)。而鎘脅迫對產卵量有急性影響(僅中間濃度(G2)處理), 長期無影響, 且與濃度間也無密切關系(圖7f)。同時回歸分析也表明, LA與產卵量間、重金屬離子與產卵量也有密切關系, 而回歸方程的參數體現了不同離子脅迫的影響差異 (圖7g、7h)。
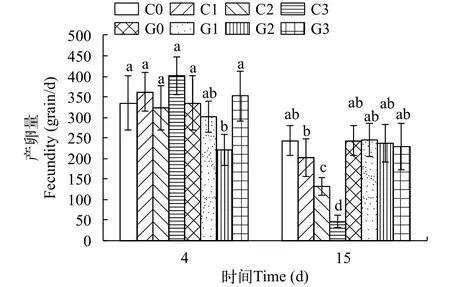
圖6 不同濃度銅或鎘脅迫對海洋青鳉產卵的影響Fig. 6 Fecundities of Oryzias melastigma under different concentrations of cadmium or copper

圖7 銅或鎘脅迫過程中魚體各指標間的關系分析Fig. 7 Correlative analysis of related indicators of fish in copper or cadmium poisoning test
3 討論
機體疲勞主要體現為能量或肌肉力量的下降[27],表現為一系列生化指標的變化, 如乳酸(LA)上升、血尿素氮(BUN)上升和乳酸脫氫酶(LDH)活性上升等[28,29]。肌肉中LA增加會導致氫離子濃度升高, 從而造成機體疲勞, LDH作用是催化LA重新形成丙酮酸, 消除LA積累[30]。因此, LA和LDH兩個指標可以用來反應機體疲勞及能量消耗程度[31]。在本試驗中, 海洋青鳉體內的LA水平(圖1), 短期銅脅迫時有所升高, 而后逐漸減少到低于對照組。一是表明魚體逐漸適應了銅的低毒脅迫, 生理狀態逐漸恢復;二是低濃度的銅離子有生物活性作用, 微量的重金屬對水體中的生物是有益的[32]。而鎘脅迫與銅脅迫剛好相反, 隨鎘濃度升高而顯著上升, 盡管隨著魚體的適應LA略有下調, 但仍保持在高位狀態。從LDH的變化也能看出(圖2), 銅脅迫前期略有升高后逐漸下降, 而鎘脅迫下LDH活性逐漸增強。LDH活性可反應機體能量代謝的水平, 肌體受損也會導致LDH活性增高[33,34]。從上述的LA和LDH變化看, 海洋青鳉對銅脅迫的適應快, LDH也無較大的波動, 表明低濃度銅基本沒有對海洋青鳉造成損傷, 僅在前期抗脅迫中機體能量有所消耗。而鎘脅迫短期內沒有出現太大影響, 但到試驗后期海洋青鳉才表現出疲勞損傷特性(LDH活性高)。睪酮(T)也有一定的消除疲勞功效[35]。在本試驗中T與LDH的變化較為吻合, 表明T可能與LDH共同作用,減少體內LA的積累。但T的變化存在著兩性間的差異性(圖3), 低濃度重金屬的脅迫對雌魚T的影響更大些, 濃度下降也更明顯。因此, 從結果看雄魚的抗疲勞能力提高了, 而雌魚T下降, 疲勞程度卻加重了。根據相關分析分析(圖7), LA與雌魚T間有很強的相關性(圖7c、7d), 可以推斷雌魚受到銅或鎘脅迫后, LA含量發生改變, 進而引起T與LDH的相應變化。而銅、鎘所造成的脅迫效應不同, 應該與魚體對不同毒物的適應能力有關。
作為性激素, T還對精子發生、卵泡發育及維持性器官正常有重要意義[36], 在雌性和雄性的生殖生理過程中都很重要[37]。一般認為, T通過調控FSH和LH的合成與分泌, 發揮生理作用[36,38,39], 本試驗也顯示T與FSH/LH間有良好的正相關關系。而FSH/LH與雌性的產卵密切相關[40—42]。本試驗在銅或鎘脅迫下的海洋青鳉體內FSH變化顯著(圖4),但整體趨勢與T相似。因此, 可以推斷雌魚T含量與其產卵量間也存在密切的正相關關系。前述中, 雌魚對重金屬較敏感, 體內的T變化比較大, 因此對產卵量的影響也比較大。LA與T間有良好的關系, 但根據上述的分析, 應該是LA升高, 而后LDH和T升高以消除LA積累, LDH和T的上升應該要滯后于LA的變化(從本實驗看, 這種滯后的時間應該是不同的毒物是不一樣的, 這需進一步進行驗證)。可以推斷LA升高造成產卵量減少, 當然是由于個體適應或抵抗毒物脅迫消耗能量所致, 如銅脅迫試驗結果; 至于鎘脅迫, 由于青鳉對鎘適應比較慢, 本試驗養殖時間不足, 因此產卵量下降現象還未出現。綜上, 可以初步建立以下的動態關系: 雌性青鳉的LA水平(疲勞程度)-T/FSH(激素變化)-產卵量(繁殖能力)。可以利用這個關系鏈開展環境脅迫程度的分析。盡管銅或鎘低濃度脅迫均對海洋青鳉的生理生化產生影響, 且有關的生理生化指標間存在較好的相關性。但是, 有機體內的各項指標都是動態變化的, 而水生生物對低濃度重金屬也有適應性,只有超出了機體的調節能力, 才會影響到生物的生理穩態[43]。不過, 通過本實驗發現, 生物對不同重金屬種類的響應不同, 且存在性別差異, 還有采樣的時間點等都會對結果產生影響, 影響結果的準確性。因此, 還需要開展更多的試驗研究, 完善相關關系鏈分析, 為準確開展生物監測提供依據。
4 結論
綜上所述, 本研究發現低濃度的銅或鎘脅迫都會影響海洋青鳉的生理生化指標, 且這種影響呈現出明顯的性別差異性。低濃度的銅或鎘脅迫導致雌性青鳉體內的LA升高, 限制了性激素(T、FSH)分泌, 從而導致產卵量下降, 且這種關系與銅或鎘濃度間呈正相關關系, 并隨脅迫時間的延長而效應下降。但銅或鎘脅迫對這種關系的影響也有較大的差異, 如對應的相關回歸方程的斜率、系數及變化周期等。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
音樂探索(2022年2期)2022-05-30 21:01:37
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
航空世界(2020年10期)2020-01-19 14:36:20
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國塑料(2016年3期)2016-06-15 20:30:00
