
建蘭種子萌發(fā)的影響因素研究
2020-03-27 12:15:34王思蘄曹小勇胡選萍秦公偉趙垚
農(nóng)業(yè)與技術(shù) 2020年4期
王思蘄 曹小勇 胡選萍 秦公偉 趙垚

摘要:以建蘭不同發(fā)育狀態(tài)種子為材料,通過無菌播種的方法,研究種子成熟度(授粉后60d、90d、180d、270d、360d、480d的種子,下文授粉后天數(shù)用DAP表示),液體與固體培養(yǎng)方式以及植物生長調(diào)節(jié)劑6-BA,NAA等因素對種子萌發(fā)的影響。結(jié)果表明:DAP60種子未萌發(fā),DAP90、DAP180、DAP270種子的萌發(fā)率高于DAP360、DAP480種子;液體培養(yǎng)基中種子萌發(fā)率為39.56%高于固體培養(yǎng)基萌發(fā)率23.47%;6-BA與NAA都可促進建蘭種子萌發(fā),且萌發(fā)率隨著激素濃度增加而升高,種子在1/2MS+6-BA 2mg/L中萌發(fā)率達到96.27%,且萌發(fā)指數(shù)優(yōu)于NAA及其它濃度6-BA。
關(guān)鍵詞:建蘭;種子成熟度;固液培養(yǎng);6-BA;NAA
中圖分類號:S-3? 文獻標(biāo)識碼:ADOI:10.19754/j.nyyjs.20200229007
建蘭(Cymbidium ensifolium),又名四季蘭,為蘭科蘭屬多年地生草本植物,生于海拔600~1800m的山谷、疏林、灌叢或草叢中,廣泛分布于東南亞和南亞各國。我國主要產(chǎn)地有浙江、福建、廣東、海南等。葉片寬1cm左右,長40~60cm,形狀如劍,花多葶長,花清香,花梗和花瓣多數(shù)為淡黃色,唇瓣上有暗點紫塊,在我國有悠久的栽培歷史[1,2]。
建蘭單個蒴果內(nèi)具有大量種子,和其它蘭科植物相似其種子細小,無胚乳,種子發(fā)育緩慢,萌發(fā)率低[3-5]。目前,對建蘭組織培養(yǎng)研究主要在莖尖與側(cè)芽誘導(dǎo)原球莖[6-9]、根狀莖增殖與分化[10-12]、花芽誘導(dǎo)[13]、遺傳多樣性分析[14,15] 等方面。同時,建蘭不同種子成熟度與萌發(fā)率的關(guān)系未見報道。因此,本試驗通過研究建蘭種子成熟度、液體與固體培養(yǎng)方式、植物生長調(diào)節(jié)劑6-BA及NAA對萌發(fā)的影響,確定適合建蘭種子萌發(fā)的培養(yǎng)基,以提高種子萌發(fā)率,為推進建蘭離體培養(yǎng)的效率提供技術(shù)支持。
1材料和方法
1.1材料
建蘭植株培養(yǎng)于陜西理工大學(xué)生物科學(xué)與工程學(xué)院實驗準(zhǔn)備室,2017年9月—2018年6月盛花期進行人工授粉,2019年1月8日首個果實開裂成熟,從授粉到果實成熟需要約480d。選取DAP60、DAP90、DAP180、DAP270、DAP360、DAP480的無菌種子用于不同成熟度種子萌發(fā)的培養(yǎng),DAP360用于液體、固體培養(yǎng)基與不同濃度6-BA、NAA培養(yǎng)。
1.2培養(yǎng)方式與培養(yǎng)條件
采用液體、固體培養(yǎng)方式,以30mL的玻璃螺口頂空瓶為培養(yǎng)容器,每瓶內(nèi)培養(yǎng)基約為10~15mL,每個處理重復(fù)3次,培養(yǎng)溫度25±1℃,種子激素處理暗培養(yǎng),其他處理光照強度1000~2000Lx,光照12h/d。
1.3試驗方法
1.3.1建蘭果實消毒
果實采收后,去除干枯花瓣,用毛筆粘上肥皂刷洗果皮,流水沖洗2h后移入超凈工作臺,用75%酒精浸泡30s,0.1%升汞消毒15min,無菌水清洗3次,在無菌培養(yǎng)皿中切去多余果柄、干枯合蕊柱后,切分果實,得到的種子作為實驗材料。
1.3.2不同成熟度種子培養(yǎng)
以1/2MS液體培養(yǎng)基為萌發(fā)培養(yǎng)基。分別將不同發(fā)育時期的無菌種子均勻播種至培養(yǎng)基內(nèi)150d后,統(tǒng)計各時期種子萌發(fā)(萌發(fā)為種子發(fā)育至下文B階段)數(shù)量。
1.3.3液體、固體培養(yǎng)
將無菌種子均勻播種于1/2MS固體、液體培養(yǎng)基中360d后,統(tǒng)計萌發(fā)種子數(shù)量。固體培養(yǎng)基中添加7g/L瓊脂,液體培養(yǎng)基不添加瓊脂。每瓶種子數(shù)量約為70~200粒。
1.3.4不同濃度6-BA與NAA培養(yǎng)
將無菌種子均勻播種于以下7種液體培養(yǎng)基中:(A1):1/2MS(對照組); (A2):1/2MS+0.1mg/L 6-BA;(A3):1/2MS+1mg/L 6-BA;(A4):1/2MS+2mg/L 6-BA;(A5):1/2MS+0.1mg/L NAA;(A6):1/2MS+1mg/L NAA;(A7):1/2MS+2mg/L NAA。培養(yǎng)360d后,統(tǒng)計種子萌發(fā)數(shù)量。每瓶種子數(shù)量約為70~200粒。
1.4萌發(fā)指標(biāo)計量
萌發(fā)率=B+C+D+EA+B+C+D+E×100%
萌發(fā)指數(shù)=10B+20C+30D+40EA+B+C+D+E×100%
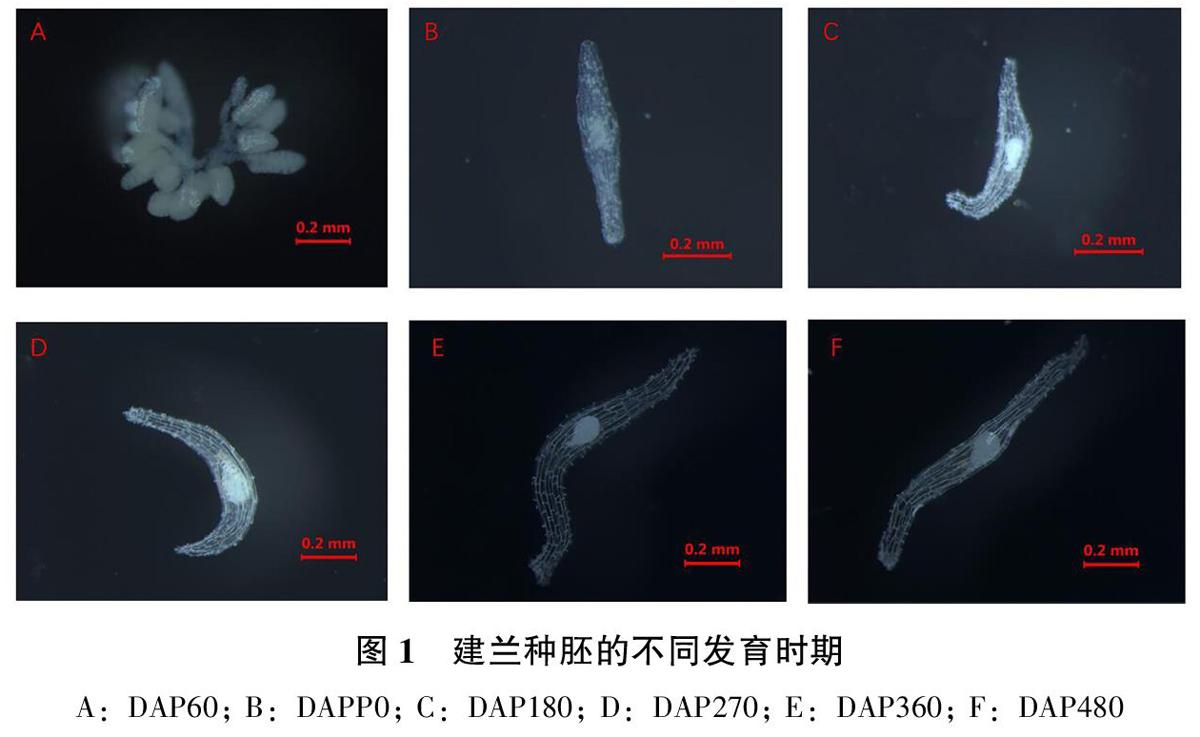
ABCDE分別表示建蘭種子萌發(fā)過程5個階段(未萌動、膨大、突破種皮、伸長、分叉)。
A:種胚未萌動,胚的長度小于0.19cm;
B:種胚膨大但未突破種皮,胚的長度0.19~0.23cm;
C:原球莖突破種皮,原球莖長度0.24~0.35cm;
D:根狀莖大部分已突破種皮或與種皮分離,此階段許多根狀莖變成綠色,長度大于0.35cm;
E:一個根狀莖上的分叉數(shù)量大于1。
通過對種子萌發(fā)各階段數(shù)量統(tǒng)計,得到萌發(fā)指數(shù)。
1.5統(tǒng)計分析
采用Microsoft Excel 2007、SPSS19.0和Prism 5對試驗結(jié)果進行統(tǒng)計學(xué)分析。
2結(jié)果與分析
2.1種子成熟度對萌發(fā)的影響
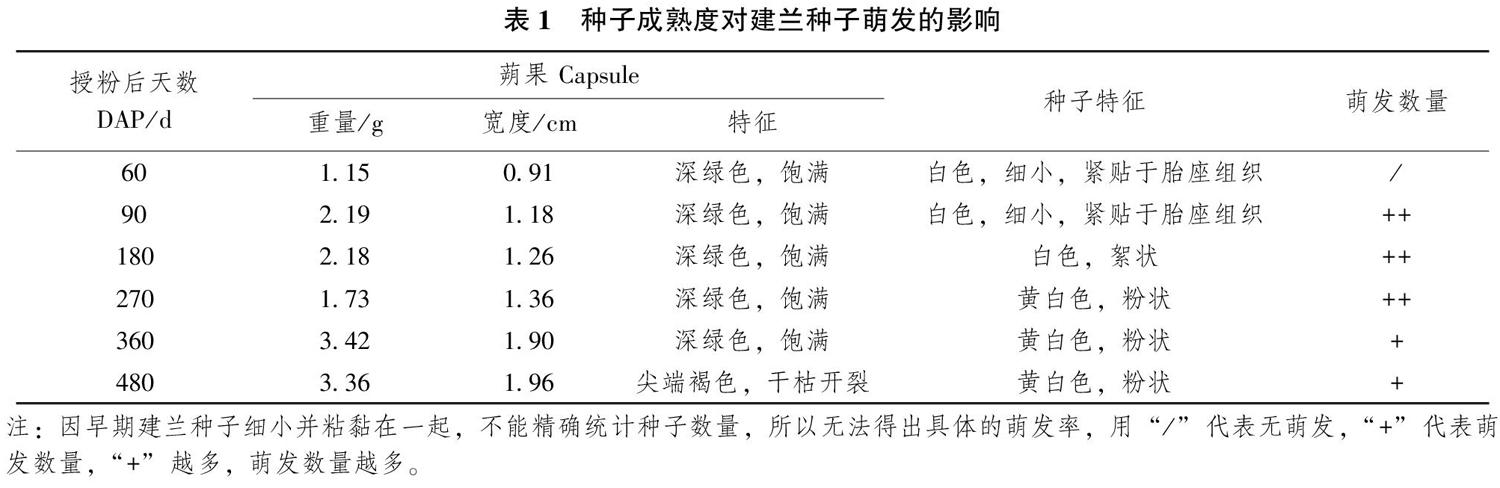
建蘭種子的成熟度顯著影響種子的萌發(fā)數(shù)量,成熟度低以及完全成熟的種子萌發(fā)數(shù)量均較低。DAP60的蒴果瘦小,其寬度與DAP90-480蒴果存在明顯差異,解剖蒴果種子白色,細小,緊貼于胎座組織,在培養(yǎng)基上未萌發(fā);DAP90種子白色,細小,可見發(fā)育較小的胚,在培養(yǎng)基上出現(xiàn)萌發(fā);DAP180種子白色,伸長呈絮狀,顯微鏡下可見明顯胚體,在培養(yǎng)基上出現(xiàn)萌發(fā);DAP270-480種子黃白色,粉狀,均在培養(yǎng)基上出現(xiàn)萌發(fā),蒴果的寬度隨著生長時間的延長逐漸增加。DAP90、DAP180、DAP270種子萌發(fā)數(shù)量高于DAP60、DAP360、DAP480,說明建蘭種子成熟度對種子萌發(fā)有較大影響。
DAP/d蒴果 Capsule重量/g寬度/cm特征種子特征萌發(fā)數(shù)量601.150.91深綠色,飽滿白色,細小,緊貼于胎座組織/902.191.18深綠色,飽滿白色,細小,緊貼于胎座組織++1802.181.26深綠色,飽滿白色,絮狀++2701.731.36深綠色,飽滿黃白色,粉狀++3603.421.90深綠色,飽滿黃白色,粉狀+4803.361.96尖端褐色,干枯開裂黃白色,粉狀+注:因早期建蘭種子細小并粘黏在一起,不能精確統(tǒng)計種子數(shù)量,所以無法得出具體的萌發(fā)率,用“/”代表無萌發(fā),“+”代表萌發(fā)數(shù)量,“+”越多,萌發(fā)數(shù)量越多。
2.2液體、固體培養(yǎng)方式對建蘭種子萌發(fā)的影響
液體、固體培養(yǎng)方式對建蘭種子的萌發(fā)影響較大,液體培養(yǎng)方式種子的萌發(fā)率為39.56%,較固體培養(yǎng)萌發(fā)率23.47%更高。在液體培養(yǎng)基中種子充分接觸水與營養(yǎng)物質(zhì),同時稀釋了種子萌發(fā)抑制物,從而促進種子萌發(fā)[21]。因此在本試驗中,液體培養(yǎng)基較固體培養(yǎng)基更能提高建蘭種子的萌發(fā)率。
2.3不同濃度6-BA與NAA對建蘭種子萌發(fā)的影響
不同濃度6-BA與NAA對建蘭種子萌發(fā)率影響均有顯著性差異。不添加任何外源植物生長調(diào)節(jié)劑時,種子也可萌發(fā),萌發(fā)率為16.43%。添加不同濃度6-BA與NAA后,可明顯提高種子的萌發(fā)率,隨著6-BA與NAA濃度的增加,萌發(fā)率和萌發(fā)生長指數(shù)逐漸增加,在濃度為2mg/L時,種子的萌發(fā)率可分別達到96.27%、95.84%,極顯著高于對照組,較對照組增加了486%、483%;種子的萌發(fā)指數(shù)可分別達到27.76、20.82,遠高于對照組,較對照組增加了658%、469%,這說明2mg/L 6-BA與NAA都能顯著提高種子萌發(fā)率,且種子在2mg/L 6-BA中萌發(fā)后的生長速度更優(yōu)。因此,建蘭種子適宜萌發(fā)的培養(yǎng)基為1/2MS+2mg/L 6-BA+25g/L蔗糖(暗培養(yǎng))。
3討論
成熟度是蘭科植物種子萌發(fā)的影響因素之一,許多蘭科植物種子完全成熟后種子的萌發(fā)率遠低于未成熟種子。本實驗研究表明,建蘭種子不同發(fā)育階段對其萌發(fā)有較大影響:DAP60種子過于細小、幼嫩,尚無萌發(fā);DAP360及以后的種子萌發(fā)率低于DAP90、DAP180、DAP270種子,這說明建蘭種子過于幼嫩或高度成熟均不利于種子萌發(fā),此研究結(jié)果與大花杓蘭(C. macranthum)、小葉兜蘭(P. barbigerum)、C. reginae、黃花杓蘭 (C. flavum)結(jié)果相似。相關(guān)研究發(fā)現(xiàn),內(nèi)源性脫落酸濃度隨著種子成熟度的增加而增加,高濃度的脫落酸是阻礙種子萌發(fā)的主要因素,這可能是導(dǎo)致該現(xiàn)象的原因之一。Yamazaki and Miyoshi認為,在種子成熟過程中木質(zhì)素等物質(zhì)在胚胎周圍的內(nèi)珠被積累,對誘導(dǎo)成熟種子休眠具有重要作用。也有人認為,蘭科植物種子在成熟過程中內(nèi)種皮會變成一層“Carapace”的封閉結(jié)構(gòu),從而導(dǎo)致成熟種子的種皮滲透性差,種子萌發(fā)困難。因此,成熟種子內(nèi)的抑制萌發(fā)物質(zhì),或種皮滲透性差,抑或是以上因素共同作用引起種子休眠,降低了成熟種子萌發(fā)率;過于幼嫩種子可能由于胚發(fā)育不完全,而未發(fā)生萌發(fā)。
雖然建蘭種子在液體培養(yǎng)基中萌發(fā)率更高的結(jié)果與Zeng S、丁長春關(guān)于兜蘭的研究結(jié)果一致,但液體培養(yǎng)方式不一定適合所有蘭科植物種子。這可能是因為液體培養(yǎng)存在缺少氧氣及長時間培養(yǎng)營養(yǎng)不足的問題。因此在液體培養(yǎng)的過程中可以減少液體培養(yǎng)的時間、定期加入營養(yǎng)液并及時將萌發(fā)的原球莖轉(zhuǎn)接至固體培養(yǎng)基中,從而促進種子萌發(fā)及萌發(fā)后原球莖的健康生長。
參考文獻
[1] 陳心啟,吉占和.中國蘭花全書[M].北京:中國林業(yè)出版社,1998:70-100.
[2]中國科學(xué)院中國植物志編輯委員會.中國植物志(第18卷)[M].北京:科學(xué)出版社,1999:213.
[3]田梅生,王伏雄,錢南芬,等.四季蘭種子離體萌發(fā)及器官建成的研究[J].Journal of Integrative Plant Biology,1985(05):455-459,564.
[4]劉思思,陳娟,郭順星.蘭科植物種子萌發(fā)的研究進展[J].種子,2015,34(06):43-50.
[5]郭順星,徐錦堂.蘭科植物種子無菌萌發(fā)的研究[J].種子,1990(05):36-37,58.
[6]王熊,陳季楚,劉桂云,顧梅仙,包慈華.建蘭和秋蘭原球莖的發(fā)生及其無性系的建立[J].植物生理學(xué)報,1981(02):203-207.
[7]王熊,張菊野,連宏坤,龔頌福,金于榮.素心建蘭無性繁殖系的建立及其開花[J].園藝學(xué)報,1988(03):205-208,218.
[8]傅向東,錢秀紅,毛碧增,李德葆.幾種理化因子對建蘭原球莖生長分化的影響[J].浙江農(nóng)業(yè)大學(xué)學(xué)報,1997(05):65-68.
[9]孫安慈,任玲,王伏雄.建蘭根狀莖增殖條件的研究[J].植物學(xué)通報,1989(03):147-150.
[10]毛碧增,林蔚紅,錢秀紅,傅向東,李德葆.影響建蘭原球莖增殖的若干因素[J].浙江農(nóng)業(yè)大學(xué)學(xué)報,1998(01):68-70.
[11]高麗,李洪林,楊波.基本培養(yǎng)基與生長調(diào)節(jié)劑組合對素心建蘭根狀莖增殖和芽分化的影響[J].亞熱帶植物科學(xué),2007(04):13-15.
[12]劉翠華,蒙陽,王朝雯,張小燕,楊柏云.建蘭組織培養(yǎng)及根狀莖增殖的動力學(xué)[J].南昌大學(xué)學(xué)報(理科版),2012,36(03):264-267,272.
[13]賈勇炯,曹有龍,王水,唐琳,徐鶯,陳放.彩心建蘭花枝莖節(jié)離體培養(yǎng)的研究[J].四川大學(xué)學(xué)報(自然科學(xué)版),2000(01):94-97.
[14]白堅,胡旭,周淑婷,王慧中.47個建蘭品種的SRAP遺傳多樣性分析[J].植物遺傳資源學(xué)報,2012,13(03):376-380.
[15]胡薇,黃儒珠,潘曉華,李錦鳳,孫端.建蘭38個品種的RAPD分析[J].園藝學(xué)報,2008(02):289-294.
