煙草中脫落酸的相關研究進展
2020-03-30 04:01:02馬斯琦劉萬峰呂林李義強高軍平杜興華蒲文宣
中國煙草科學 2020年1期
關鍵詞:煙草
馬斯琦 劉萬峰 呂林 李義強 高軍平 杜興華 蒲文宣
摘? 要:煙草的生長發育極易受到不良環境的影響,導致煙葉產量和品質下降,其對不利環境因素的抗性受植物激素的調控。植物激素脫落酸是植物體內五大類激素之一,參與植物的生長發育,可提高植物對非生物逆境脅迫的抗性。以脫落酸為核心的基因工程技術是目前作物抗逆育種的重點策略之一,本文重點總結煙草中脫落酸的相關研究進展,包括一般功能、抵御非生物逆境的功能、代謝和信號轉導途徑等,以期為煙草抗逆育種提供理論支持和技術方法借鑒。
關鍵詞:煙草;脫落酸;非生物逆境
Review on Abscisic Acid (ABA) in Tobacco
MA Siqi1, LIU Wanfeng2, LYU Lin1, LI Yiqiang1, GAO Junping2, DU Xinghua3, PU Wenxuan2*
(1. Tobacco Research Institute, Chinese Academy of Agricultural Sciences, Qingdao 266101, China; 2. Chinese Tobacco Hunan Industrial Corporation, Changsha 410007, China; 3. Shaanxi Provincial Tobacco Monopoly Administration, Xi'an 710061, China)
Abstract: Tobacco plant is quite vulnerable to abiotic stresses during its growth and development, which often results in the yield and quality decline of tobacco leaves. Thus it is urgent to enhance tobacco resistance to abiotic stress. The phytohormone abscisic acid (ABA) in tobacco can regulate various physiological processes and promote tobacco adaptation to abiotic stresses. ABA engineering might be one of the key strategies to obtain tobacco with tolerance to abiotic stress. This paper focuses on the research progress of ABA in tobacco, including general function, function in abiotic stress, metabolism and signaling pathway, etc., which might provide reference for the practical applications and research on tobacco stress-resistance breeding.
Keywords: tobacco; abscisic acid; abiotic stress
目前干旱、低溫等非生物逆境脅迫嚴重影響作物的生長、制約作物的產量。煙草是重要的經濟作物和模式植物,喜溫喜光且對水分依賴大,生長發育極易受到環境的影響,因此如何增強煙草自身的抗逆性從而提高煙草的穩產性,已成為煙草行業穩定發展、國家農業可持續發展的重要研究課題。
植物為了應對不利于生長的環境因素,體內進化出一系列的響應機制和主動防御模式,但是這些機制和模式仍不足以保證它們在不利的環境中生存。因此,科學家們嘗試多種策略以提高植物在非生物逆境脅迫下的抗性,其中包括以脫落酸(Abscisic acid,ABA)為核心的基因工程。脫落酸因能促使子葉脫落而得名,廣泛存在于高等植物中,與生長素、乙烯、赤霉素、細胞分裂素并列為植物五大類激素。脫落酸是一種小分子物質,屬于倍半萜類化合物,主要在細胞質中合成,具有加速器官脫落、促進芽和種子的休眠、調節種子發育、
引起氣孔關閉、增加植株抗逆性等功能[1-2]。脫落酸因參與各種生物與非生物逆境脅迫應答享譽為“逆境激素stress hormone”[3-4]。本文主要總結煙草中脫落酸調控作物發育和抗逆的研究進展,包括其一般功能、抵御非生物逆境的功能及其合成、代謝、信號轉導途徑等。
1? 脫落酸的一般功能
1.1 ?調節種子的休眠與萌發
種子的休眠與萌發是植物生命周期中非常重要的階段,也是一個非常重要的生態經濟性狀。種子的休眠是一種生存策略,不僅為種子的傳播擴散爭取時間,還可以促使種子在最適宜的環境條件下萌發。研究發現,脫落酸可以促進種子休眠、抑制種子萌發:在胚發育晚期,胚產生的脫落酸促進種子休眠和抑制穗萌,而在種子萌發過程中,脫落酸的含量可負調控種子吸水吸脹程度[5]。在煙草種子相關研究中發現,成熟的干種子中脫落酸含量較高,而在萌發過程中脫落酸含量逐漸降低。煙草種子吸水萌發大致可分為3個階段,分別是干種子迅速吸水膨脹期、吸水平穩期和吸水旺盛期。在煙草種子吸水前兩個階段脫落酸對種子吸水沒有明顯的抑制作用,但第3個階段明顯受脫落酸抑制。因此,在后熟的煙草種子中降低脫落酸的濃度,可促進煙草種子的休眠釋放,提高煙草種子的萌發效率[6-7]。
1.2 ?調節葉片衰老進程
葉片衰老是植物生命周期后期必經的生理過程,受到植物內源物質和外部環境的多種因素調控。衰老的過程是植物主動將營養成分轉移到新生葉片和種子,蓄積能量促使植物存活和再發育的過程,早衰會縮短農作物生長周期,導致產量降低。已有研究表明,脫落酸對植物葉片衰老具有十分顯著的影響,例如脫落酸可以引起植物葉片變黃(衰老的標志之一)、葉面噴施脫落酸可以顯著加速葉片的衰老等。科學家們還發現在一些持綠性較好的品種葉片中脫落酸含量較少,降低體內脫落酸的濃度可以減緩葉片衰老。除此之外,脫落酸還可以誘導植物體內衰老相關基因的表達等[8]。擬南芥中脫落酸合成或者信號轉導被抑制的突變體,植株的衰老會發生不同程度的改變和延遲,也從遺傳學的角度上印證了脫落酸參與到植物的衰老調控[9]。
煙草葉片的衰老與煙葉生產密切相關,直接影響著煙草產品的產量、品質和效益。煙草是葉用經濟作物,煙葉落黃程度與后續的調制加工息息相關,葉片落黃程度受發育階段、養分供應、水分調控、環境溫度等多種因素影響,對煙葉產品外觀質量、物理特性、化學成分、煙氣特征等質量因素有決定性的影響。在煙草葉片衰老過程中,同樣發現脫落酸的合成和信號傳遞都增強,脫落酸信號傳遞正調控基因的表達量升高、負調控基因的表達量降低[10]。
1.3? 調控氣孔開關
氣孔是植物葉片表面由保衛細胞形成的開孔,可控制植物的氣體交換、葉表面水分蒸發等。光照可以誘發氣孔打開,脫落酸的積累和高CO2可以促使氣孔部分關閉或關閉[11]。氣孔關閉時,氣體交換減少、光合產物減少,而蒸騰作用減少可減少葉片失水。在干旱條件下,脫落酸調控保衛細胞離子轉運,促進氣孔關閉,防止氣孔打開,減少水分流失[12]。研究發現,脫落酸可以通過激活質膜上的S型離子通道,抑制質膜上H+-ATPase的活性,誘導氣孔中活性氧(ROS)積累等多種機制來調節氣孔關閉[13]。在干旱情況下氣孔的及時關閉可以降低細胞間二氧化碳濃度和蒸騰速率,減少水分喪失,提高煙草的抗旱性[14]。
2? 脫落酸提高植株抵御非生物逆境脅迫的功能
脫落酸具有提高植株抵御非生物逆境脅迫的功能,非生物逆境包含干旱、澇害、高溫、低溫、鹽漬、重金屬離子脅迫等,本文主要總結其在煙草生產過程中影響較大的干旱、澇害、低溫、高溫、鹽漬這幾種非生物逆境脅迫條件下的功能。
2.1? 脫落酸與干旱
干旱是主要的非生物逆境脅迫之一,嚴重影響植物的生長和生物量,世界上有超過一半的耕地存在不同程度的干旱。干旱脅迫通常會導致植物發生滲透脅迫,而滲透脅迫會影響植物體內水分的流失和保持。脫落酸在脅迫過程中具有類似控制器的功能,可以通過正調控內源機制抵抗逆境,還可以通過調控一些基因的表達來激活抗性機制。已有研究表明,干旱可以影響脫落酸核心信號通路基因的表達量,包括受體PYLs、蛋白磷酸酶PP2Cs、磷酸激酶SnRKs等成員[15]。
干旱不僅限制煙葉產量,對煙葉品質也極為不利,例如干旱可以造成煙葉的水分含量發生變化從而影響其加工質量、原料損耗,同時還會增加煙葉的總氮含量、生物堿、蛋白質含量令煙葉的色澤、香氣、吃味等品質下降,降低煙葉中水溶性糖、鉀含量令煙葉柔軟性、彈性等品質變差。雖然在煙草中還尚未有關于ABA和煙草耐旱機制的深入報道,但是煙草根部含有豐富的ABA合成前體物質和酶,在土壤有效水變化時,根部可精確感受并作出相應反應,合成大量的ABA并通過木質部導管和韌皮部篩管向植物的其他部位運輸,降低氣孔導度,有效的減少煙草葉片的蒸騰作用,減少水分喪失,提高水分利用率,從而令植物產生抗旱性[16]。
2.2? 脫落酸與澇害
水分是植物生長的不可缺少的關鍵因素,但是
水分過高也不利于植物生長,甚至導致澇害的發生。近年來,全國乃至全球極端氣候頻繁出現,雨量分布極不均衡,再加上存在排水不暢等因素影響,經常發生大面積、不同程度的澇害,影響著農業產業的發展。澇害可抑制植物的生長,影響植物的光合作用,促使葉片老化、彎曲,促使老根枯死、不定根誘發等。煙草生產過程中水分過多會引起植株生長受阻甚至死亡,葉片總氮含量上升、總磷、總鉀、還原糖、煙堿含量下降導致煙葉品質下降[17]。已有研究表明在澇害脅迫下,植物體內的脫落酸含量會發生變化,促使葉片中葉綠素降解、氣孔關閉,降低光合作用和蒸騰速率,保持水勢平衡,但是關于脫落酸在煙草抵抗澇害中的應用還有待深入研究[18]。
2.3? 脫落酸與低溫
低溫是植物在生命周期中經常會遭受的環境脅迫之一,低溫通常可以造成植物細胞功能紊亂,包括膜損傷、ROS的積累、蛋白質的降解、有毒物質的積累等。植物在面對低溫時,體內會發生多種改變:(1)基因表達的改變,(2)細胞膜構象發生改變,(3)脯氨酸、可溶性糖、抗氧化物等低溫保護物質的積累,(4)內源脫落酸水平的升高[8]。
低溫是煙草生產中遇到的主要災害,特別是幼苗移栽到大田這段時期經常遭受“倒春寒”等低溫天氣的影響,由于煙草是喜溫植物,對低溫十分敏感,零下2~3 ℃的極端低溫可導致煙草死亡,在苗期12~16 ℃的持續低溫就會導致煙草提早開花進入生殖生長,抑制煙草植株的高度和葉片數目從而影響煙草的品質和產量。已有研究發現,與脫落酸信號轉導途徑相關的許多基因都參與到植物的低溫應答中。擬南芥中許多與脫落酸信號轉導相關基因的突變體均出現了抗低溫的表型,例如AREB/ABF、CBF/DREB類轉錄因子等也參與到植物對低溫的響應[19-20]。另外,在草本植物中外施脫落酸可以提高植物對低溫的抗性,木本植物中脫落酸既可以幫助植物提高低溫抗性,還可以通過調控幼芽萌發幫助植物更好地越冬[8]。煙草相關研究中發現JERF1可以調控脫落酸合成基因的表達,使得煙草植株體內脫落酸的含量升高,提高植株在低溫下的存活能力[21]。除去內源提高脫落酸的濃度,外源噴施脫落酸也會提高煙草植株對逆境的耐受能力[22]。
2.4 ?脫落酸與高溫
隨著全球氣溫的日益變暖,高溫脅迫已經成為限制植物分布和產量的重要因素。高溫脅迫可以影響植物的一系列生理和生化反應,包括氣孔關閉、水分散失、光合作用的抑制、活性氧積累、增加膜脂質過氧化程度、蛋白質降解和酶活性喪失,最終導致嚴重的細胞損傷或細胞死亡[23]。
雖然煙草是喜溫植物,但是長期生長在溫度相對較高的環境下,煙草的生長發育也會受到不同程度的抑制,同時還會出現煙葉卷曲褶皺、葉片變厚、葉面積變低、加速衰老,甚至出現“高溫逼熟”的現象,對煙草生產極為不利。已有研究表明高溫可誘導植物體內內源脫落酸的積累,可提高植株在高溫脅迫下體內抗過氧化酶的活性,同時降低丙二醛的含量。另外,脫落酸的積累可誘導植物體內相關逆境應答基因的表達和熱激蛋白的產生,提高植株對高溫的抵抗能力[23]。煙草中,外施一定濃度的脫落酸可提高煙草的耐熱性,降低高溫對煙草造成的影響,但其具體的分子機制還有待深入研究[24]。
2.5 ?脫落酸與鹽(漬)
土壤鹽漬化在世界范圍內的農業問題,鹽漬化環境對植物的傷害主要體現在高離子濃度產生的滲透脅迫和離子毒害兩個方面,它們可以改變植物的膜結構,抑制植物的呼吸作用和光合作用,嚴重影響植物的生長發育,影響作物的產量和質量[25]。目前研究結果表明,鹽脅迫下植物脫落酸含量顯著增加,并刺激氣孔的關閉、基因的表達以及滲透物質的積累。鹽脅迫時,噴施外源脫落酸可以降低鹽脅迫對煙苗膜脂的過氧化程度,維持較低丙二醛含量、減少活性氧物質含量,維持膜的穩定性,提高煙草對鹽脅迫的耐受能力[26]。
3? 脫落酸代謝途徑及信號轉導路徑
脫落酸在植物生長發育、抵御逆境環境方面都具有十分重要的作用,關于脫落酸在植物體內的代謝與信號轉導途徑的研究也日益完善,為植物抗逆遺傳機制的解析和植物精準育種提供了大量的思路和理論依據。
3.1? 脫落酸的代謝
脫落酸主要通過從頭合成途徑合成,在質體和細胞質中完成。此途徑的合成前體是玉米黃質素,又因玉米黃質素是類胡蘿卜素的一種,因此這種合成途徑也稱為類胡蘿卜素分解途徑。玉米黃質環氧酶ZEP在胞質中催化玉米黃質素變成反式紫黃質素,重排后變成環氧類胡蘿卜素,在9-順式-環氧類胡蘿卜素雙加氧酶(9-cis-epoxycarotenoid dioxygenase, NCED)的催化下氧化分解,分解產物被運輸到細胞質中,進一步催化產生脫落酸。在此合成過程中,9-順式-環氧類胡蘿卜素雙加氧酶是最關鍵的酶,起著重要的調節作用。脫落酸的降解代謝主要是通過脫落酸羥基化和脫落酸結合物的生成來實現,其中以8-羥基化最為常見。細胞色素P450家族成員脫落酸-8-羥基化酶(CYP707A)催化脫落酸形成8-羥基-脫落酸,隨即生成產物紅花菜豆酸(Phaseic acid, PA)和二羥紅花菜豆酸(Dihydro-phaseic acid, DPA)導致脫落酸失活[27]。
9-順式-環氧類胡蘿卜素雙加氧酶(NCED)基因最早從玉米突變體vp14中鑒定和發現,vp14葉片中脫落酸含量與對照相比顯著下降,造成其葉片失水速率加快[28]。水稻中研究發現有5個NCED家族成員參與到脫落酸合成。OsNCED3基因的表達量在干旱、鹽、過氧化氫等脅迫條件下上調,并且其突變體對這些非生物逆境脅迫更敏感[7],這些結果表明OsNCED3是一個逆境脅迫的正調控因子。另外,OsNCED4和OsNCED5的表達量也受到干旱脅迫的誘導表達,調節植株對干旱脅迫的適應能力[21]。關于脫落酸分解的相關重要基因在水稻中也已有相關報道,研究發現水稻中有3個脫落酸-8-羥基化酶催化脫落酸轉化為紅花菜豆酸[29],分別命名為OsABAox1-3,其中OsABAox1的表達量受到冷脅迫的誘導,OsABAox2和OsABAox3的表達量受到脫落酸處理的誘導,這些結果表明脫落酸降解相關的基因也參與到了逆境脅迫應答中[30]。
在煙草基因組中有18個9-順式-環氧類胡蘿卜素雙加氧酶(NCED)家族成員得到鑒定,可分為4個亞家族,通過生物信息學分析,第一亞家族成員NtNCED1-8與ABA生物合成有關。干旱脅迫可以誘導NtNCED3基因的表達和內源ABA的積累,且NtNCED3基因表達和內源ABA積累的規律一致。也有研究發現,在煙草中過表達柑橘、黃龍膽、柱花草等植物NCED基因,均可提高轉基因植物內源ABA水平、植株抗逆性等[31]。
3.2? 脫落酸的信號轉導
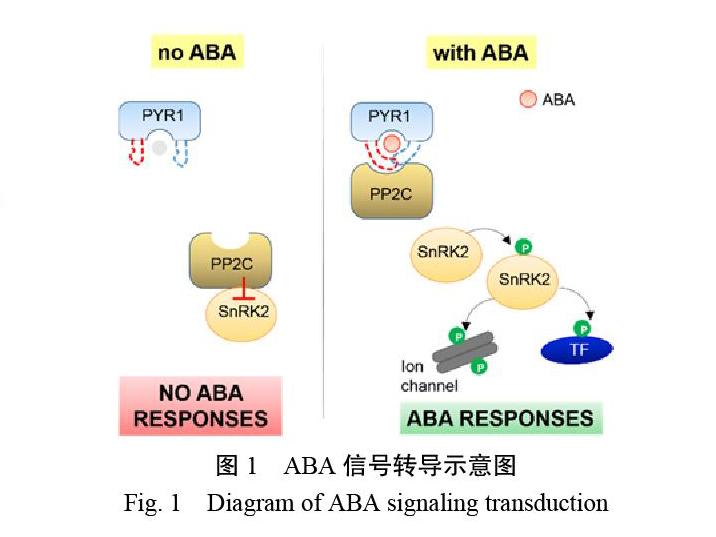
脫落酸信號轉導是響應非生物逆境應答的重要路徑,特別是對干旱脅迫應答來說屬于核心路徑[32]。許多脫落酸信號轉導的相關元件在30年前就已經得到鑒定,但是一直尚未定論,脫落酸信號轉導途徑最大的突破是利用蛋白互作和化學遺傳學的方法,發現了脫落酸信號受體,被命名為PYL。研究發現,脫落酸與信號受體結合后可以抑制蛋白磷酸酶PP2C的磷酸化活性[33-34]。隨后,科學家們利用結構生物學的手段再次證實了PYL是ABA的信號受體,并且通過對PYL-ABA-PP2C蛋白復合體結構的解析,揭示了PP2C的功能是如何被ABA和受體的復合物巧妙抑制的(圖1)[35-36]。由此,ABA信號轉導路徑逐漸建立并完善:植物在受到逆境脅迫時,大量內源ABA積累,ABA受體PYL識別ABA并與之結合,形成的復合物與PP2C磷酸酶結合并抑制其磷酸酶活性,使得絲氨酸-酪氨酸激酶SnRK2s的磷酸化活性被激活,導致下游的轉錄因子受到磷酸化而激活其轉錄活性,最終引發體內的脅迫響應機制[37-39](圖1)。
ABA受體對ABA的識別在ABA信號轉導中起著核心作用。擬南芥中ABA受體基因突變后植株對ABA不敏感,種子發芽和生根均表現出ABA不敏感的表型,而超表達后可顯著增強植株對ABA的敏感性,提高植株的抗旱性。蛋白激酶和磷酸酶催化的可逆蛋白磷酸化在細胞信號轉導中起著重
要的作用,有助于信號從細胞外傳遞到細胞內部環境[33]。蛋白磷酸酶PP2C是ABA信號的負調控因子,擬南芥A類PP2C家族成員ABI1和ABI2的突變體在不同的發育階段和植物組織中都具有對ABA敏感、耐旱等表型。蛋白激酶2(SnRK2)在細胞對干旱和脫水的反應中起著核心作用,依賴于ABA的蛋白激酶的催化活性首次在蠶豆中得到鑒定,命名為AAPK,可以正調控氣孔的關閉,提高植株的抗旱能力[40]。擬南芥中AAPK直系同源基因OST1(Open Stomata 1)/SnRK2.6也被證明在ABA介導的氣孔調節中起正調控作用[41]。這些蛋白激酶可以磷酸化其底物蛋白(如轉錄因子、離子通道和代謝酶),從而將ABA信號級聯放大[42-43]。
在煙草中ABA信號轉導路徑的部分相關基因也已被克隆鑒定,NtPYL4是煙草ABA的受體,可以與PP2C成員互作并抑制其活性,參與煙草ABA信號轉導的核心路徑[44]。NtPYL1和NtPYL9也被認為是ABA受體,在子葉和須根中表達較多,表達量受到ABA的抑制和過氧化氫的誘導[45]。根據PP2C家族特性,NtPP2C16被克隆鑒定,其表達顯著受到ABA和逆境脅迫的誘導,響應干旱、高鹽和低溫脅迫[46]。在云煙87煙草轉錄組數據中發現有18個PP2C家族成員的表達量隨著降雨量發生變化,其中NtPP2C37和NtPP2C50與擬南芥PP2C的A類家族成員同源性較高,推測可能參與到ABA信號核心路徑[47]。同樣通過生物信息學分析方法發現,煙草中有21個SnRK家族成員,可以分為3個亞家族,深入分析發現,第一亞家族的基因主要參與ABA信號轉導途徑[48]。NtSnRK2.1在煙草根部表達最高、其次是葉片,莖部的表達量最低,其表達量受到高鹽、高滲、低溫和ABA的誘導[49]。雖然煙草中關于脫落酸核心信號轉導途徑的研究已有部分進展,但相對于其他植物例如擬南芥、水稻等還亟需精確和深入。
4? 總結和展望
目前國內主要煙草栽培品種在煙葉生產上的種植年限已達20~30年之久,已經出現了不同程度的品種退化、變異和病害,可替代煙草栽培品種匱乏,嚴重制約著“兩煙”生產的穩定發展。另外,目前煙葉生產中存在由于煙葉早采搶采導致成熟度不夠、內含物質不協調等問題,因此亟需突破常規育種周期長、效率低、遺傳多樣性小的瓶頸,培育出有突破性的新品種。“十三五”以來國家啟動的“中國煙草基因組計劃”重大專項旨在源頭上攻克煙葉生產關鍵技術的瓶頸制約,推動中國煙草的研究和應用步入基因時代,實現從傳統育種向精準育種的戰略跨越。亟需持續深入對煙草中脫落酸相關基因的功能及其調控的分子機制進行挖掘和解析,以便更好地利用這些基因資源、借助分子精準育種手段培育出煙草抗逆優質新品種。
4.1 ?脫落酸在煙草抗逆育種方面的策略
脫落酸在植物生長發育和抵御非生物逆境脅迫方面都發揮著重要的功能,以脫落酸為核心的精準育種已成為作物精準育種的重要策略之一。“分子育種”、“基因編輯育種”等精準育種方法可以通過基因和性狀的對應關系,按照需求將不同的優勢聚合,快速“設計培育”出烤煙新品種。干旱等非生物逆境條件可誘導脫落酸的積累,開啟脅迫信號的傳遞。脫落酸信號導致基因表達發生實質性變化,有研究表明近10%的擬南芥基因受脫落酸調控,參與植物生長發育和逆境應答[50],深入挖掘并解析這些基因發揮功能的分子機制有利于我們深入展開精準分子育種。目前,諸多煙草脫落酸相關的抗逆基因已被克隆,不少基因的抗逆效果在幼苗期表現顯著,但其田間效應仍需探討。如何通過遺傳學手段對這些基因加以利用,提高煙草植株在大田中的抗性將成為研究的重點。
4.2 ?脫落酸在煙葉成熟與提質方面的策略
脫落酸對煙葉的衰老調控也是煙草基因研究的重要培育點。煙草與大多數作物延緩衰老提高產量的需求不同,在煙葉成熟采收及烘烤方面,需要促進葉片田間成熟和在烤房內的后熟,提高烤后煙葉的質量。通過ABA對煙葉衰老的調控,消耗煙葉成熟過程中煙堿和過多的淀粉,促進煙葉內在化學成分協調性,提高中部煙葉和上部煙葉可用性,ABA不僅在煙草新品種培育方面具有提高抗逆性的潛力,在煙葉成熟調控方面,也具有提高煙葉質量的潛力。
參考文獻
- RAGHAVENDRA A S, GONUGUNTA V K, CHRISTMANN A, et al. ABA perception and signaling[J]. Trends in Plant Science, 2010, 15(7): 395-401.
- SAH S K, REDDY K R, LI J X. Abscisic acid and abiotic stress
tolerance in crop plants[J]. Frontiers in Plant Science, 2016, 7:571.
- KANCHAN V, NEHA U, NITIN K, et al. Abscisic acid signaling and abiotic stress tolerance in plants: a review on current knowledge and future prospects[J]. Frontiers in Plant Science, 2017, 8(120): 161.
- MEHROTRA R, BHALOTHIA P, BANSAL P, et al. Abscisic acid and abiotic stress tolerance-different tiers of regulation[J]. Journal of Plant Physiology, 2014, 171(7): 486-496.
- VANSTRAELEN M, BENKOVA E. Hormonal interactions in the regulation of plant development[J]. Annu Rev Cell Dev Biol, 2012, 28(28): 463-487.
- 李振華. 煙草種子萌發的激素效應研究進展[J]. 云南農業大學學報(自然科學),2013,28(3):416-423.
- 生利霞,王倩,孟祥毅,等. 植物耐澇分子機理研究進展[J]. 分子植物育種,2017,15(7):2823-2828.
SHENG L X, WANG Q, MENG X Y, et al. Research progress on molecular mechanism of waterlogging tolerance in plants[J]. Molecular Plant Breeding, 2017, 15(7): 2823-2828.
- 劉慶宇,趙越,熊江波,等. 澇害對烤煙光合特性的影響[J]. 江西農業大學學報,2014,36(1):50-55.
LIU Q Y, ZHAO Y,XIONG J B, et al. A study on the effect of waterlogging on photosynthetic characteristics in flue-cured tobacco[J]. Acta Agriculturae Universitatis Jiangxiensis, 2014,36(1): 50-55.
- CHOI H, HONG J, HA J, et al. ABFs, a family of ABA-responsive element binding factors[J]. J Biol Chem, 2000, 275(3): 1723-1730.
- LEE S J, KANG J Y, PARK H J, et al. DREB2C interacts with ABF2, a bZIP protein regulating abscisic acid-responsive gene expression, and its overexpression affects abscisic acid sensitivity[J]. Plant Physiol, 2010, 153(2): 716-727.
- WU L, CHEN X, REN H, et al. ERF protein JERF1 that transcriptionally modulates the expression of abscisic acid biosynthesis-related gene enhances the tolerance under salinity and cold in tobacco[J]. Planta, 2007, 226(4): 815-825.
- ZANDALINA S I, MITTLER R, BALFAGON D, et al. Plant adaptations to the combination of drought and high temperatures[J]. Physiol Plant, 2018, 162(1): 2-12.
- 萬正林,羅慶熙. SA、ABA、BR誘導植物抗高溫可能機理的研究[J]. 西南農業學報,2007(6):1399-1403.
WANG Z L, LUO Q X. The mechanism of SA, ABA and BR in inducing heat resistance on plants[J]. Southwest China Journal of Agricultural Sciences. 2007(6): 1399-1403.
- 劉中明. 過表達SlJA2通過水楊酸途徑降低轉基因煙草的高溫抗性[D]. 泰安:山東農業大學,2017.
LIU Z M. Over-expression of SIJA2 decreases heat tolerance of transgenetic tobacco plants via salicylic acid patyway[D]. Tai'an: Shandong Agricultural University, 2017.
- 岑怡紅,沙波. 煙草的抗逆性研究進展[J]. 北京農業,2012(9):216-217.
CEN Y H, SHA B. Advances in the research of stress tolerance of tobacco[J]. Beijing Agriculture, 2012(9): 216-217.
- 李武琴,曾乙心,陳光勇,等. 外源激素對鹽脅迫下煙草幼苗生理抗性的影響[J]. 山東農業科學,2018,50(9):36-39.
LI W Q, ZENG Y X, CHEN G Y, et al. Effects of exogenous hormones on physiological resistance of tobacco seedlings under salt stress[J]. Shandong Agricultural Sciences, 2018, 50(9): 36-39.
- NAMBARA E, MARION-POLL A. Abscisic acid biosynthesis and catabolism[J]. Annu Rev Plant Biol, 2005, 56:165-185.
- TAN B C, SCHWARTZ S H, ZEEVAART J A D, et al. Genetic control of abscisic acid biosynthesis in maize[J]. PNAS, 1997, 94(22): 12235-12240.
- HUANG Y, GUO Y, LIU Y, et al. 9-cis-Epoxycarotenoid dioxygenase 3 regulates plant growth and enhances multi-abiotic stress tolerance in rice[J]. Front Plant Sci, 2018, 9: 162.
- SAIKA H, OKAMOTO M, MIYOSHI K., et al. Ethylene promotes submergence-induced expression of OsABA8ox1, a gene that encodes ABA 8'-hydroxylase in rice[J]. Plant Cell Physiol, 2007, 48(2): 287-298.
- 白戈,楊大海,姚恒,等. 煙草NtNCED基因的鑒定分析[J]. 分子植物育種,2017,15(10):3907-3912.
BAI G, YANG D H, YAO H, et al. Identification and analysis of NtNCED gene in tobacco[J]. Molecular Plant Breeding, 2017, 15(10): 3907-3912.
- ZHU J K. Salt and drought stress signal transduction in plants[J]. Annu Rev Plant Biol, 2002, 53: 247-273.
- MA Y, SZOSTKIEWICZ I, KORTE A, et al. Regulators of PP2C phosphatase activity function as abscisic acid sensors[J]. Science, 2009, 324(5930): 1064-1068.
- PARK S Y, FUNG P, NISHIMURA N, et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins[J]. Science, 2009, 324(5930): 1068-1071.
- MELCHER K, NG L M, ZHOU X E, et al. A gate-latch-lock mechanism for hormone signalling by abscisic acid receptors[J]. Nature, 2009, 462(7273): 602-608.
- YIN P, FAN H, HAO Q, et al. Structural insights into the mechanism of abscisic acid signaling by PYL proteins[J]. Nat Struct Mol Biol, 2009, 16(12): 1230-1236.
- SINGH A, JHA S K, BAGRI J, et al. ABA inducible rice protein
phosphatase 2C confers ABA insensitivity and abiotic stress tolerance in Arabidopsis[J]. PLoS ONE, 2015, 10(4): 1.
- SARUHASHI M, GHOSH T K, ARAI K, et al. Plant Raf-like kinase integrates abscisic acid and hyperosmotic stress signaling upstream of SNF1-related protein kinase2[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(46): 6388-6396.
- UMEZAWA T, NAKASHIMA K, MIYAKAWA T, et al. Molecular basis of the core regulatory network in ABA responses: sensing, signaling and transport[J]. Plant and Cell Physiology, 2010, 51(11): 1821-1839.
- LI J, WANG X Q, WATSON M B, et al. Regulation of Abscisic acid-induced stomatal closure and anion channels by guard cell AAPK Kinase[J]. Science, 2000, 287: 300–303.
- MUSTILLI A C, MERLOT S, VAVASSEUR A, et al. Arabidopsis OST1 protein kinase mediates the regulation of stomatal aperture by abscisic acid and acts upstream of reactive oxygen species production. plant cell[J]. Plant Cell, 2002, 14(12): 3089-3099.
- YOSHIDA R, HOBO T, ICHIMURA K, et al. ABA-activated SnRK2 protein kinase is required for dehydration stress signaling in Arabidopsis[J]. Plant and Cell Physiology, 2002, 43(12): 1473-1483.
- FUJITA Y, YOSHIDA T, YAMAGUCHI S K. Pivotal role of the AREB/ABF-SnRK2 pathway in ABRE-mediated transcription in response to osmotic stress in plants[J]. Physiologia Plantarum, 2013, 147(1): 15-27.
- LACKMAN, P, GONZALEZ-GUZMAN M, TILLEMAN S, et al. Jasmonate signaling involves the abscisic acid receptor PYL4 to regulate metabolic reprogramming in Arabidopsis and tobacco[J]. Proc Natl Acad Sci U S A, 2011, 108(14): 5891-5896.
- 黃佳,夏玉珍,趙影影,等. 煙草脫落酸受體基因NtPYR1和NtPYL9的克隆及表達模式分析[J]. 煙草科技,2015,48(7):1-8.
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
