水稻抗細菌性條斑病主效基因位點的遺傳分析及定位
2020-04-22 06:25:12覃麗萍岑貞陸
西南農業學報 2020年1期
關鍵詞:水稻
覃麗萍,謝 玲*,岑貞陸
(1.廣西農業科學院微生物研究所,廣西 南寧 530007;2.廣西農業科學院植物保護研究所,廣西 南寧 530007)
【研究意義】水稻細菌性條斑病(OryzasativaL.,簡稱水稻細條病)由水稻黃單胞菌棲稻致病變種Xanthomonasoryzaepv.Oryzicola(Fang,et al.)Swings et al.引起,是我國南方及東南亞國家水稻上的一種主要細菌性病害,通常減產5 %~20 %,在適宜的氣候條件下感病品種減產在30 %以上[1],嚴重影響水稻的產量和品質,被我國列為植物檢疫對象。目前水稻細條病防治主要依賴化學藥物,但效果不明顯,且長期使用易造成病菌產生抗(耐)藥性、環境污染及農藥殘留等負面影響[2],不利于保持生態平衡和促進農業的可持續發展。培育和種植抗病品種是防治細條病經濟、有效且環保的方法,而挖掘抗病相關基因資源、掌握數量豐富的抗性基因位點是培育抗病品種的關鍵。主效基因控制的質量抗性能很好地抵御病菌侵染及繁殖而表現出免疫或高抗,是當前利用的主要植物抗病性類型,但目前對水稻細條病抗性基因的研究主要集中在數量性狀基因(QTL)定位,對主效基因的定位研究相對較少。SSR標記具有較高的應用價值,可應用于動植物的遺傳圖譜構建、目標基因標定及指紋圖繪制等研究中[3-6]。因此,廣泛深入發掘抗水稻細條病國際稻品種BJ1的主效基因,豐富水稻抗病基因位點數量,對水稻抗細條病的分子標記輔助選擇具有重要意義。【前人研究進展】多數學者認為,水稻對細條病的抗性屬QTL控制的數量性狀,分別在第1、2、3、4、5、7、8和11號染色體定位到十幾個抗性QTL,并獲得相應的分子標記[7-13]。但也有學者通過對不同水稻抗源材料進行研究發現,水稻對細條病的抗性由顯性或隱性主效基因控制,屬質量性狀。何月秋等[14]研究發現13個雜交稻組合對細條病的抗性受1~2顯性主效基因控制;張紅生等[15]研究證實秈稻品種IR36對細條病的抗性由1對隱性主效基因控制,Dular、IR26和IR1545-339則由1對顯性主效基因控制;徐建龍等[16]研究認為抗源品種Hashikalmi和Dular的抗性受2對獨立的隱性主效基因控制,90IRBBN44受1對隱性主效基因控制;黃大輝等[17]研究表明野生稻DP3的抗性由2對隱性重疊作用的基因控制;賀文愛等[18]研究認為野生稻抗源DY3、DY17和DY20 的抗性由 2 對隱性主效基因控制,而DY16 的抗性由 1 對隱性主效基因控制。國際稻BJ1全生育期抗細條病,曾有不同學者嘗試分析其對細條病的抗性遺傳規律,如周明華等[19]認為BJ1對細條病的抗性由1對顯性基因控制,Nayak等[20]則認為BJ1對細條病的抗性受3對獨立隱性基因控制,二者的研究結果雖存在差異,但均認為BJ1對細條病的抗性由主效基因控制。【本研究切入點】目前,雖然已發現抗源BJ1攜帶抗細條病主效基因,但鮮見對其抗性基因進行定位的研究報道。【擬解決的關鍵問題】以高抗細條病的國際稻品種 BJ1與感病品種油占8號為親本,通過雜交、自交構建F2代分離群體,采用混合分組分析法(BSA)和簡單重復序列(SSR)分子標記技術結合抗病性鑒定,對BJ1攜帶的抗性基因位點進行初步定位,為水稻抗細條病分子標記輔助育種打下基礎。
1 材料與方法
1.1 試驗材料
抗病親本為國際稻品種BJ1,感病親本為秈稻品種油占8號,感病對照為TN1,接種菌株蓮塘13為廣西水稻細條病病菌優勢致病型Ⅲ型,均由本研究課題組保存提供。
1.2 群體構建
BJ1與油占8號進行雜交,收獲F1代種子后播種,F1代植株自交得到F2代。在廣西農業科學院玻璃大棚的栽培池中種植抗、感親本及F1代各10株,種植F2代251株。各生育期除抗病性鑒定時外均保持淺水灌溉。
1.3 抗病性鑒定
在早稻分蘗盛期(5月底)進行接種:將 2 根大頭針以約8 mm的間距插進橡皮膠中,高壓滅菌備用;供試菌株在肉汁凍培養基上培養2 d后,用無菌水沖洗掉菌苔,配成3×108CFU/mL懸浮液(當天使用);將大小與培養皿內空間相當、厚約2 cm的無菌海綿置于培養皿內并倒入菌液,使海綿吸足菌液,然后將水稻新生的完全展開葉正面朝上平置于海綿碟上,用準備好的大頭針在葉片中上部刺一下,注意使橡皮膠上的2 根針分別刺在葉片中脈兩側,稍按壓橡皮膠擠出海綿中的菌液使之滲入葉片傷口;在接種過程中根據菌液損耗情況適時給海綿補充菌液;每株接種2 片葉。接種 20 d后,參考農秀美等[21]的方法(略有修改)進行病情調查和標準劃分,測量接種點病斑的長度,每個單株測量10個病斑,取其平均值作為評判單株對細條病抗性水平的依據。詳細劃分標準見表1,以 10.0 mm 的病斑長度作為劃分抗、感分界線。接種完成后至病情調查期間保持深水灌溉。
1.4 抗性基因初步定位
1.4.1 DNA提取及基因池構建 采用 CTAB 法提取抗、感親本及 F2代群體各單株的基因組DNA,結合抗病性鑒定選取極端抗病(免疫I和高抗HR)和極端感病(高感HS)F2代單株各15株的DNA分別混合成抗病基因池和感病基因池。提取方法:每株取新鮮嫩葉約 0.1 g,剪碎,放入內有一顆直徑約5 mm鋼珠的2.0 mL EP管中,加入600.0 μl CATB提取液;使用組織研磨儀充分研磨;65 ℃水浴30 min,期間將樣品上下顛倒2次;加入等體積(600.0 μl)氯仿—異戊醇混合液(混合比例為24∶1),搖勻后10 000 r/min離心10 min;取上清液400.0 μl,置于1.5 mL EP管中;加入2倍體積(800.0 μl)無水乙醇,上下顛倒1次,4 ℃靜置30 min;10 000 r/min離心10 min,棄上清液,自然晾干沉淀,加入100.0 μl ddH2O,置于4 ℃或-20 ℃保存備用。
表1 水稻抗、感細條病劃分標準及其相應癥狀表現
Table 1 The standard of division for rice bacterial leaf streak and the corresponding symptoms
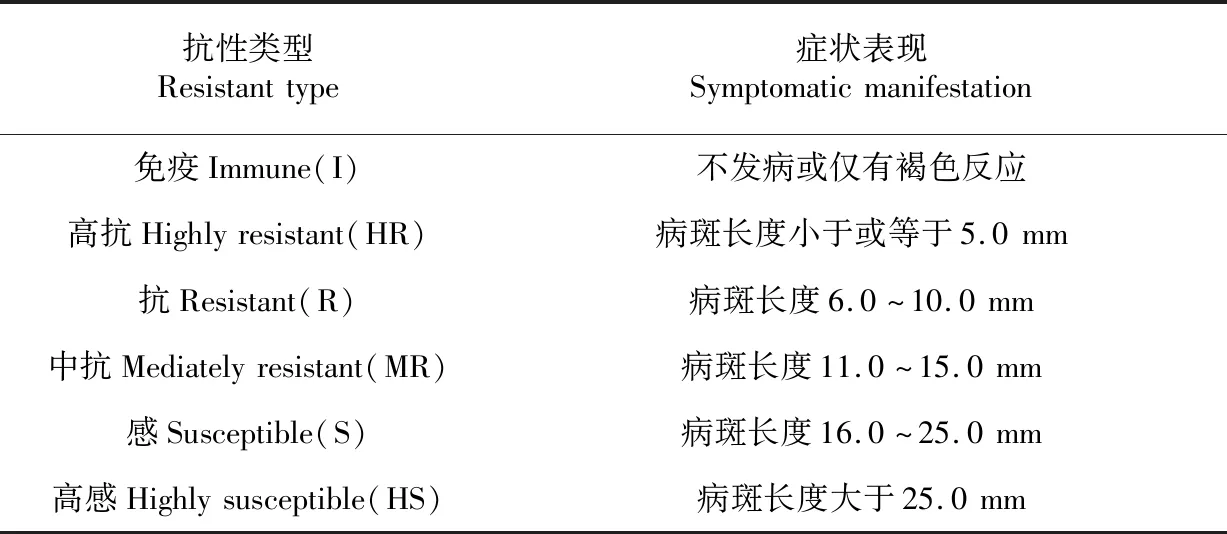
抗性類型Resistanttype癥狀表現Symptomaticmanifestation免疫Immune(I)不發病或僅有褐色反應高抗Highlyresistant(HR)病斑長度小于或等于5.0mm抗Resistant(R)病斑長度6.0~10.0mm中抗Mediatelyresistant(MR)病斑長度11.0~15.0mm感Susceptible(S)病斑長度16.0~25.0mm高感Highlysusceptible(HS)病斑長度大于25.0mm
1.4.2 SSR分子標記篩選 從已公布的水稻SSR引物[由捷瑞生物工程(上海)有限公司合成]中選出在水稻12條不同染色體上均勻分布的279對引物,對抗、感親本進行初篩,選擇在抗、感親本間具有多態性的SSR引物在抗、感基因池中進行多態性檢測。
PCR 反應體系20.0 μl:包括2.0 μl 10×Buffer(含 Mg2+)、2.0 μl dNTPs (2.5 mmol/L)、2.0 μl引物(10 μmol/L)、2.0 μl模板DNA、0.1 μlTaqDNA 聚合酶(5 U/μl)、11.9 μl滅菌純凈水。擴增程序:95 ℃ 預變性 5 min;95 ℃ 變性 30 s,52 ℃復性 30 s,72 ℃ 延伸 30 s,進行35 個循環;72 ℃延伸 5 min,4 ℃ 保存待用。PCR擴增產物經 8.0 %非變性丙烯酰胺凝膠電泳分離、快速銀染后觀察。銀染步驟(各步驟均需輕微振蕩):10.0 %乙醇(v/v)+0.5 % 冰醋酸(v/v)固定10 min; 0.2 %硝酸銀(w/w)滲透10 min,雙蒸水漂洗 2次,每次10 s; 10.0 %硫代硫酸鈉漂洗30 s,去除多余的銀離子; 1.5 %氫氧化鈉(w/w)+1.0 %(v/v)甲醛顯色,待條帶清晰顯現時用清水沖洗。
1.4.3 連鎖標記 利用在抗病基因池和感病基因池間表現出多態性的SSR標記引物對F2群體進行擴增,擴增產物按1.4.2的方法進行電泳分離、銀染觀察,采用“1-2”系統記載各單株的帶型,與抗病親本帶型一致的記為“1”,與感病親本帶型一致的記為“2”,雜合帶型記為“3”,缺失記為“0”。
1.5 統計分析
采用OriginPro 9.1分析F2代群體基因型與表型的相關性。
2 結果與分析
2.1 抗源抗性的遺傳特征
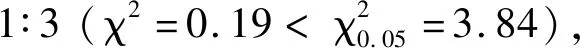
2.2 親本間SSR分子標記的初篩結果
從已公布的水稻SSR引物中選出均勻分布于水稻12條不同染色體上的279對引物,對抗、感親本進行初篩,結果有RM310、RM330A和RM257等65對引物在親本間檢測到多態性,各染色體上的SSR分子標記引物在親本間的多態性表現率存在明顯差異,變幅在0 %~42.9 %,平均多態性表現率為23.3 %(表3)。
表2 親本、F1和F2代對細條病的抗性表現Table 2 Resistance reaction of parents,F1 and F2 to bacterial leaf streak

a:感病材料發病情況;b:F2代不同單株的發病情況a:Symptoms on susceptible plants;b:Symptoms on different F2 plants圖1 水稻接種細條病菌株蓮塘13后20 d的癥狀表現Fig.1 Symptoms of bacterial leaf streak after 20 days inoculated with Liantang 13
2.3 抗性基因的定位結果
在抗病、感病親本間呈多態性的65對引物中,RM 258(上游引物5′-CTCCCTGGCCTTTAAAGCTGTCG-3′,下游引物5′-GACGAACAGCAGCAGAAGAGAAGC-3′)在抗、感基因池間呈多態性。利用RM258擴增F2代群體單株251株(圖2),結果檢測到抗病帶型68株,其中表型鑒定為抗病的有60株、感病的有8株,基因型與表型符合率為88.2 %;檢測到感病帶型(感病親本帶型和雜合帶型)175株,其中表型鑒定為抗病的有4株、感病的有171株,基因型與表型符合率為97.7 %;剔除缺失的8株,有243株F2代群體的基因型與表型符合率為93.0 %(表4),基因型與表型的相關系數為0.5705,二者的相關性達顯著水平(P<0.05),表明SSR分子標記RM258與BJ1攜帶的抗水稻細條病隱性主效基因存在連鎖關系。對照已公布的水稻分子遺傳連鎖圖,可知RM258位于第10號染色體上約48.8 cM處。
3 討 論
國際稻 BJ1不但高抗細條病,而且抗白葉枯病[22]、抗蟲和抗倒伏,綜合抗性好[23],雖然其農藝性狀較差,但仍不失為一個培育抗水稻細條病中間材料的好抗源。本研究以BJ1為供體,油占8號為受體,構建其F2代分離群體對抗源BJ1的抗性遺傳特性進行研究,結果表明,來自國際稻圃的秈稻品種BJ1對細條病的抗性屬隱性遺傳,受1對主效基因控制。周明華等[19]利用BJ1與感病品種南農秈2號、金剛 30雜交、回交構建F2、BC1和BC2群體研究BJ1對細條病的抗性遺傳規律,發現其抗性由1對主效顯性基因控制;Nayak等[20]通過研究BJ1與感病品種TKM-6和IR-8構建的 F2和F3代群體,發現BJ1對細條病的抗性由3對獨立主效隱性基因控制。本研究結果與周明華等[19]、Nayak 等[20]的研究結果雖然存在差異,但均表明BJ1對水稻細條病的抗性由主效基因控制,該抗性屬質量性狀。
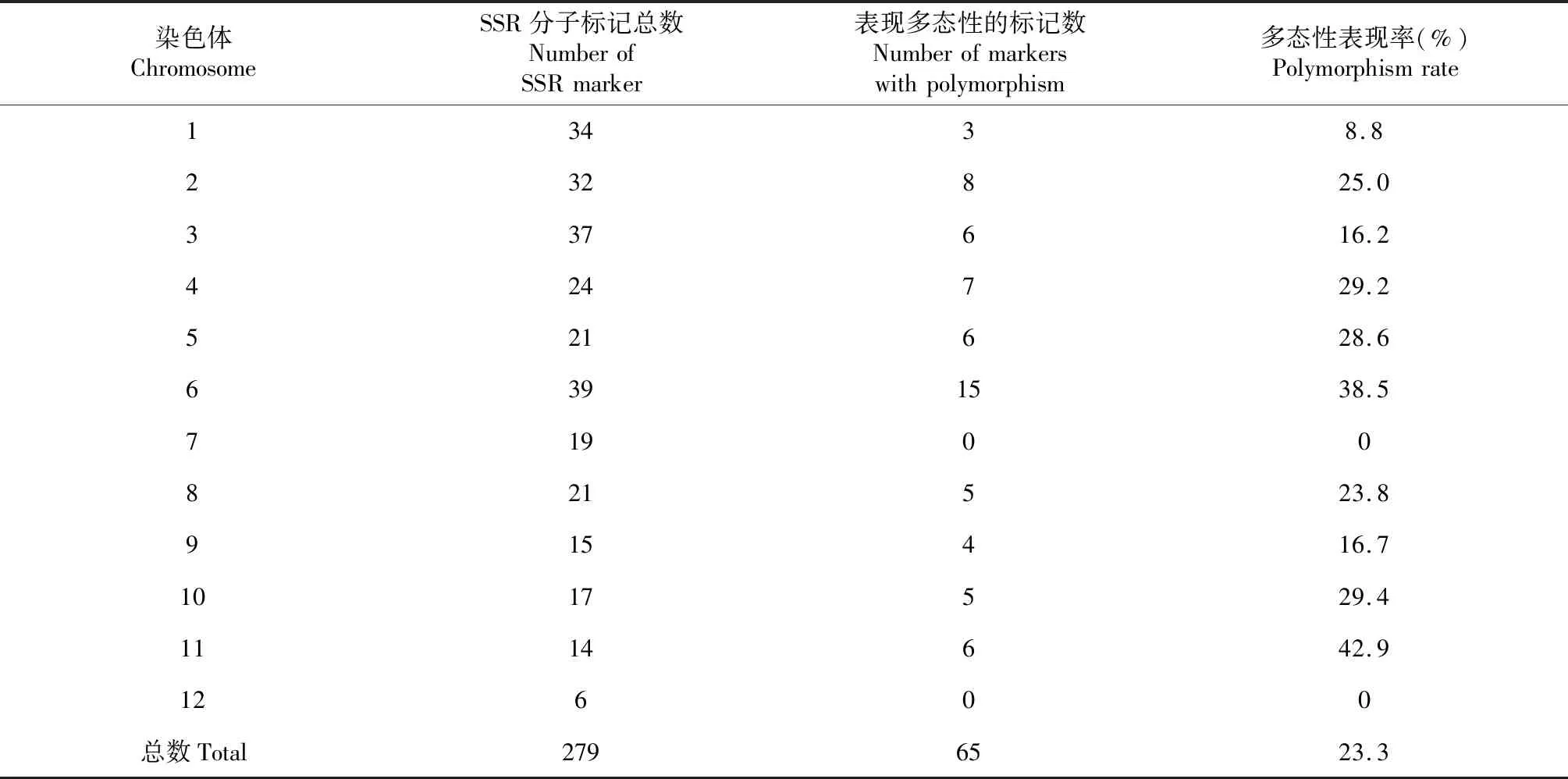
表3 各條染色體上SSR分子標記在親本間的多態性水平Table 3 The polymorphism level of SSR markers on different chromosomes in parents

M:Marker;2:抗病親本BJ1帶型;3:感病親本油占8號帶型;4~16:部分F2代單株的帶型M:Marker;2:PCR pattern of resistant parent BJ1;3:PCR pattern of susceptible parent Youzhan 8;4-16:PCR pattern of partial F2 plants圖2 SSR分子標記引物RM258擴增親本及部分F2代單株的電泳結果Fig.2 The electrophoretogram of parents and partial F2 amplified by SSR primer RM258

表4 SSR 標記引物RM 258 檢測F2代群體基因型與表型的符合率Table 4 Efficiency of genotype and phenotype of SSR marker RM258 in F2 populations
注:*表示顯著相關(P<0.05)。
Note:* represented significant correlation(P<0.05).
已有學者通過經典遺傳分析發現不少水稻種質攜帶抗細條病主效基因,但已定位的僅有普通野生稻DP3和DY19的抗性基因bls1和bls2[24-25]及美國祖傳品種Carolina Gold Select的抗性基因Xo1[26],且這3個基因分別位于第6號染色體RM1940與 RM510 間的21-kb 物理距離內、第2號染色體23474851~24484154 bp間約1.009 Mb距離內和第4號染色體長臂31358156~32448509 bp間1.09 Mb 距離內。本研究發現BJ1攜帶1對抗細條病隱性主效基因,并初步將該基因定位于第10號染色體約48.8 cM處,與 SSR分子標記RM258緊密連鎖。這是首次在第10號染色體上檢測到水稻細條病抗性基因,可為培育抗細條病水稻品種(系)提供新的抗性位點。
曾有學者認為水稻對細條病的抗性屬數量性狀,不存在主效基因[7-8],但世界上抗細條病的水稻種質資源非常豐富[17,27-30],尤其隨著研究廣度和深度的提高,不斷有新的細條病抗性主效基因或QTL被發現和定位。本研究認為,前人對水稻細條病抗性遺傳的研究結論不一致,除了與采用的菌株、接種方法、分級標準及環境因素等不同有關外,主要是因為所用稻種材料的遺傳背景不同及抗病遺傳機制存在差異,因此,水稻對細條病的抗性應該同時存在質量抗性和數量抗性。
主效基因控制的質量抗性效應較大,在基因克隆和功能利用上相對容易成功,但易因病原菌發生變異而使抗性不穩定或喪失;QTL控制的數量抗性效應微小、難以精細定位和克隆,但其抗性較持久[31]。若將BJ1的抗性主效基因導入其他攜帶多個抗性QTL、農藝性狀優良的品種中,有望培育出抗性廣譜、持久及穩定的水稻品種(系)。由于本研究僅對BJ1抗細條病基因定位進行初步探討,離水稻生產實際應用還存在差距,因此今后有待進行精細的定位、分離、克隆和轉化。
4 結 論
通過對國際稻材料BJ1進行常規雜交、選育和SSR分子標記結合抗病性鑒定,檢測到BJ1所攜帶的細條病抗性基因為隱性主效基因,位于10號染色體上約48.8 cM處,且與SSR分子標記RM258緊密連鎖,推測是一個全新的抗水稻細條病基因位點。
致 謝:廣西農業科學院水稻研究所李丹婷研究員、農保選副研究員、夏秀忠副研究員、馬增鳳副研究員、高利軍副研究員為本研究內容的完善提供了技術指導及部分實驗材料,謹向他們表示真摯的感謝!
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
