水稻C24-甾醇甲基轉移酶基因OsSMT2的生物信息學分析
2020-05-05 03:01:46黃小龍翟立升閆慧清
西南農業學報 2020年3期
黃小龍,翟立升,閆慧清
(1. 貴州師范大學 貴州省植物生理與發育調控重點實驗室,貴州 貴陽 550001;2. 貴州師范大學 國家林業局西南喀斯特山地生物多樣性保護重點實驗室,貴州 貴陽 550001;3. 貴州師范大學 生命科學學院, 貴州 貴陽 550001)
【研究意義】植物甾醇在自然界中廣泛存在,是以環戊烴全氫菲為骨架的三萜類化合物,有游離態和結合態2種形式,主要分布在植物的根、莖、葉、果實和種子中[1]。植物甾醇在食品、醫學和動物中的研究表明,其具有抑制腫瘤、降低血液膽固醇、抗菌消炎、調劑免疫和抗衰老的作用,應用前景十分廣闊[2-6]。植物甾醇與動物甾醇的區別是C-24位置多了額外的烷基,這個過程由甾醇甲基轉移酶(SMTs)完成[7],SMTs是植物甾醇代謝途徑中的關鍵限速酶[8],但其在水稻中的功能還有待更深入的研究。【前人研究進展】到目前為止,植物中大量的SMT基因被分離克隆,如菠菜[9]、大豆[10]和擬南芥[11]等。有研究表明,SMT基因在植物響應逆境脅迫的過程中發揮重要作用[12]。在高鹽脅迫下,大豆中總甾醇含量較未脅迫的降低50 %,谷甾醇含量沒有變化,但是飽和脂肪酸的含量大量增加,說明在植物中甾醇含量的穩定對于抵抗高鹽脅迫引起的傷害有重要作用[13]。在植物中乙基甾醇有利于增強細胞膜的內聚力,在面對高溫和低溫脅迫時,植物體內的甲基甾醇大量轉化成乙基甾醇,從而調節植物體的溫度變化以適應高溫和低溫脅迫[14]。植物甾醇除了響應非生物脅迫外,還響應細菌性病原體引起的生物脅迫,在擬南芥中沉默鯊烯合酶、甾醇甲基轉移酶2、C-22甾醇去飽和酶基因,加強擬南芥細胞中營養物質的外排,促進細菌性病原體的生長;在擬南芥超表達C-22甾醇去飽和酶基因的植株中營養物質的外排受到抑制,表明擬南芥通過調控甾醇含量的變化控制細胞中營養物質的外排,響應細菌引起的免疫應答[15]。【本研究切入點】甾醇甲基轉移酶在植物中的作用十分重要,干擾OsSMT1的表達使得水稻具有矮化和抗旱的表型[16],OsSMT2已經被克隆,但目前研究僅知道其參與了植物甾醇的生物合成[7],并不清楚其分子作用機制。【擬解決的關鍵問題】因此,擬通過生物信息學分析的方法研究水稻OsSMT2基因的結構、功能和表達模式,為該基因的進一步研究提供理論參考。
1 材料與方法
1.1 材料
檢索OsSMT2的相關文獻資料,以OsSMT2在RGAP(http://rice.plantbiology.msu.edu/)數據庫中的MSU Locus號(LOC_Os03g04340)為線索,下載該基因的相關序列進行分析。
1.2 方法
1.2.1 水稻OsSMT2基因的序列比對分析 通過RGAP數據庫下載OsSMT2基因的基因組序列、CDS序列和氨基酸序列,利用NCBI(National Center for Biotechnology Information)網站(https://www.ncbi.nlm.nih.gov/)中的Blast工具,將水稻OsSMT2基因的氨基酸序列與GenBank(https://www.ncbi.nlm.nih.gov/genbank/)數據庫進行比對,選取水稻(Oryzasativa)、藥用野生稻(Oryzabrachyantha)、粟(Setariaitalica)、高粱(Sorghumbicolor)、玉米(Zeamays)、節節麥(Aegilopstauschii)、大麥(Hordeumvulgare)、鳳梨(Ananascomosus)、油棕(Elaeisguineensis)、小果野芭蕉(Musaacuminata)等10種不同植物的SMT2蛋白進行序列比對分析,尋找STM2蛋白的保守結構域。選取多個物種的SMT2氨基酸序列,利用MEGA-X軟件進行同源性分析,并通過鄰接法構建進化樹。
1.2.2 水稻OsSMT2蛋白的性質和結構分析 通過Protparam (https://web.expasy.org/protparam/)所提供的蛋白質序列分析工具,對OsSMT2蛋白質的理化性質(分子量、理論pI、正負電荷殘基總數及半衰期等)進行預測與分析。利用ProtScale (https://web.expasy.org/protscale/)預測OsSMT2蛋白質的疏水性/親水性,SignalP (http://www.cbs.dtu.dk/services/SignalP/)預測OsSMT2蛋白質的信號肽,TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)預測OsSMT2蛋白質的跨膜結構域,NetPhos (http://www.cbs.dtu.dk/services/NetPhos/)預測磷酸化位點。通過SMART(Simple Modular Architecture Research Tool)工具 (http://smart.embl-heidelberg.de/)分析OsSMT2蛋白質結構域,利用在線網站GOR4 (https://prabi.ibcp.fr/htm/site/web/home)對OsSMT2蛋白質的二級結構進行預測,利用SWISS-MODEL(https://www.swissmodel.expasy.org/)同源建模網站預測OsSMT2蛋白質的三級結構。
1.2.3 水稻OsSMT2基因順式調控元件的分析 在水稻RAP-DB數據庫(https://rapdb.dna.affrc.go.jp/)中,獲取水稻OsSMT2基因起始密碼子上游2kb區域的核苷酸序列作為OsSMT2基因的啟動子區域,利用PlantCARE植物順式調控元件分析工具(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/),對OsSMT2基因的啟動子進行分析。
1.2.4 水稻OsSMT2基因共表達基因的GO 富集和KEGG富集分析 通過RED數據庫(http://expression.ic4r.org/co-search)中的共表達工具,設置PCC(Pearson correlation coefficient)為0.7,獲得OsSMT2基因的共表達基因。然后通過David網站(https://david.ncifcrf.gov/)和Kobas網站(http://kobas.cbi.pku.edu.cn/index.php)對這些基因進行GO富集和KEGG富集分析。
1.2.5 水稻OsSMT2基因的表達模式分析 通過CREP數據庫(http://crep.ncpgr.cn/)下載水稻OsSMT2基因在Minghui 63中的芯片數據,研究OsSMT2基因在水稻不同組織中的表達模式。
2 結果與分析
2.1 水稻OsSMT2蛋白的性質
水稻OsSMT2基因的基因組序列全長為1875 bp,無內含子,CDS全長1092 bp,編碼363個氨基酸。OsSMT2蛋白質分子量40 618.61,理論pI為8.19,負電荷殘基總數(Asp+Glu)為41,正電荷殘基總數(Arg+Lys)為43,分子式C1822H2828N508O514S17,總原子數5689,脂肪酸指數為82.20,預計半衰期為30 h,不穩定指數(Ⅱ)為28.51,該蛋白為不穩定蛋白。OsSMT2蛋白的親水性水平平均為-0.176,是親水性蛋白(圖1A)。信號肽S值在1~32位氨基酸之間,其中最大值在第26個氨基酸;預測值是0.797,平均分值為0.371(小于閾值0.5),說明該蛋白不含有信號肽(圖1B)。該蛋白在第6和第24個氨基酸之間存在1個跨膜結構域(圖1C)。磷酸化位點有31個,包括Ser16個、Thr8個和Tyr7個(圖1D)。
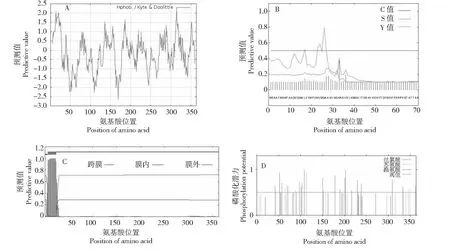
A.OsSMT2蛋白的親水性分析; B.OsSMT2蛋白的信號肽分析; C.OsSMT2蛋白的跨膜結構域預測; D.OsSMT2蛋白的磷酸化位點分析A.Hydrophilic analysis of OsSMT2; B.Signal peptide analysis of OsSMT2; C.Transmembrane domain prediction of OsSMT2; D.Phosphorylation sites prediction of OsSMT2圖1 OsSMT2蛋白的性質分析 Fig.1 Analysis of the properties of OsSMT2

h為α螺旋,e為延伸鏈,c為無規則卷曲h: Alpha helix; e: Extended strand; c: Random coil圖2 OsSMT2蛋白的二級結構預測Fig.2 Secondary structure prediction of OsSMT2
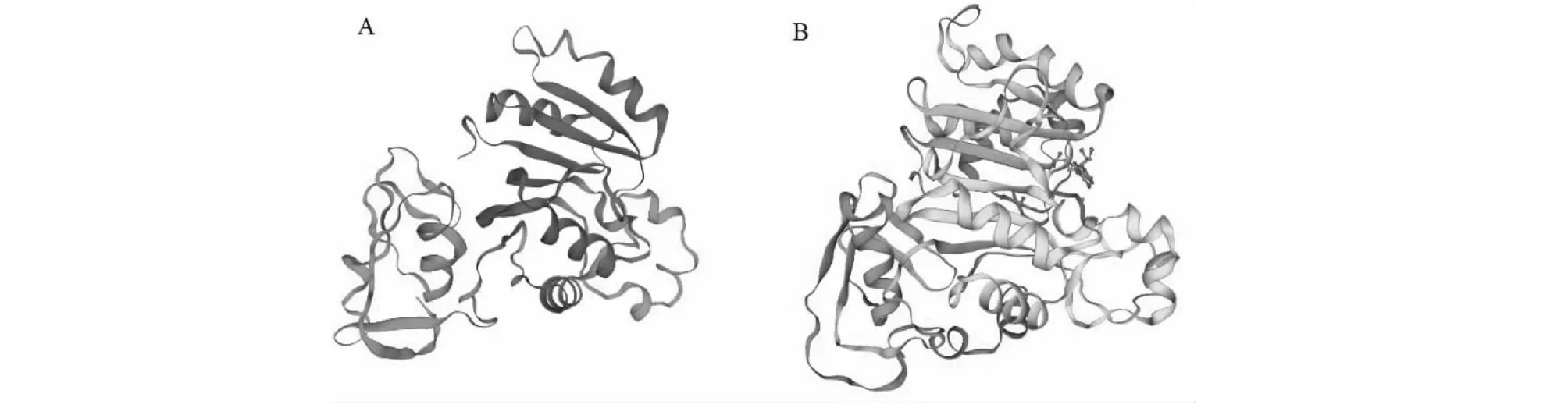
圖3 水稻OsSMT2(A)和砷(III)S-腺苷甲硫氨酸甲基轉移酶(B)蛋白的三級結構Fig.3 Tertiary structure of OsSMT2 (A) and As(III) S-adenosylmethionine methyltransferase (B)
2.2 水稻OsSMT2蛋白的結構
2.2.1 二級結構 預測結果顯示,OsSMT2蛋白有3種二級結構,其中α螺旋有132個氨基酸殘基參與,占36.36 %;延伸鏈有76個氨基酸殘基參與,占20.94 %;無規則卷曲有155個氨基酸殘基參與,占42.70 %(圖2)。
2.2.2 三級結構 從圖3看出,水稻OsSMT2蛋白質的三級結構以無規則卷曲為主。

圖4 不同物種中OsSMT2的同源蛋白比對Fig.4 Homologous alignment of OsSMT2 in different species

圖5 不同物種中SMT2蛋白的保守結構域Fig.5 Conserved domains of SMT2 in different species
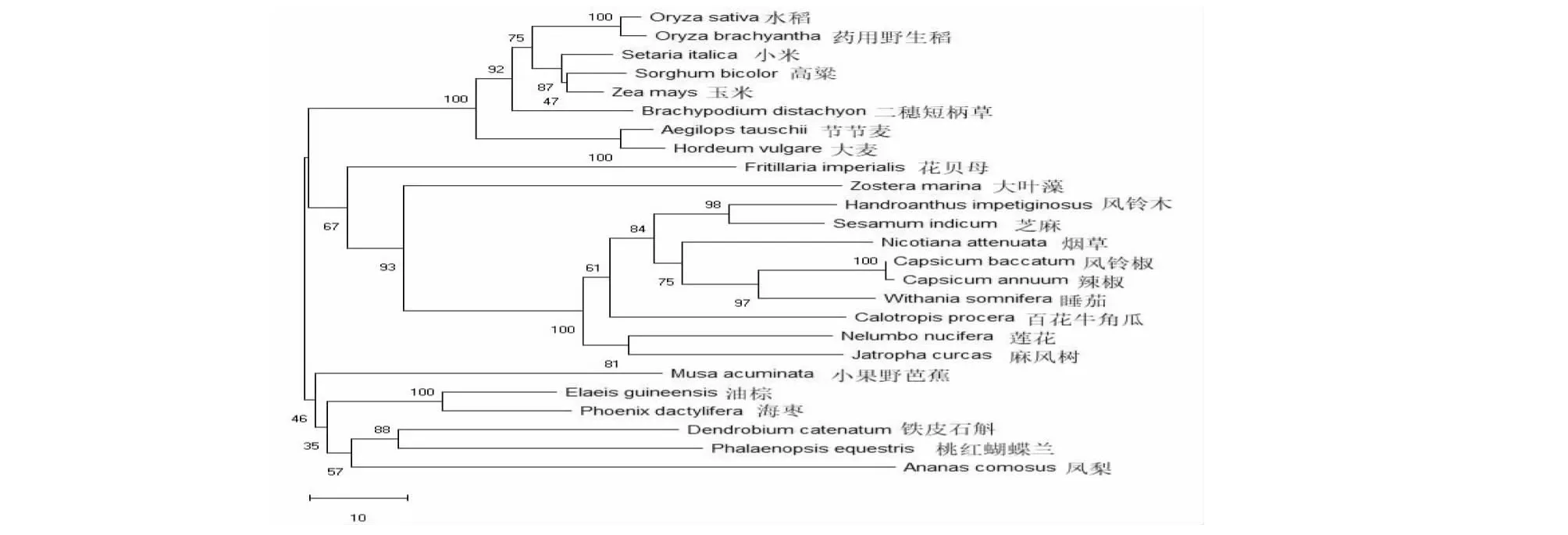
圖6 OsSMT2基因的系統發育樹Fig.6 Phylogenetic tree of OsSMT2
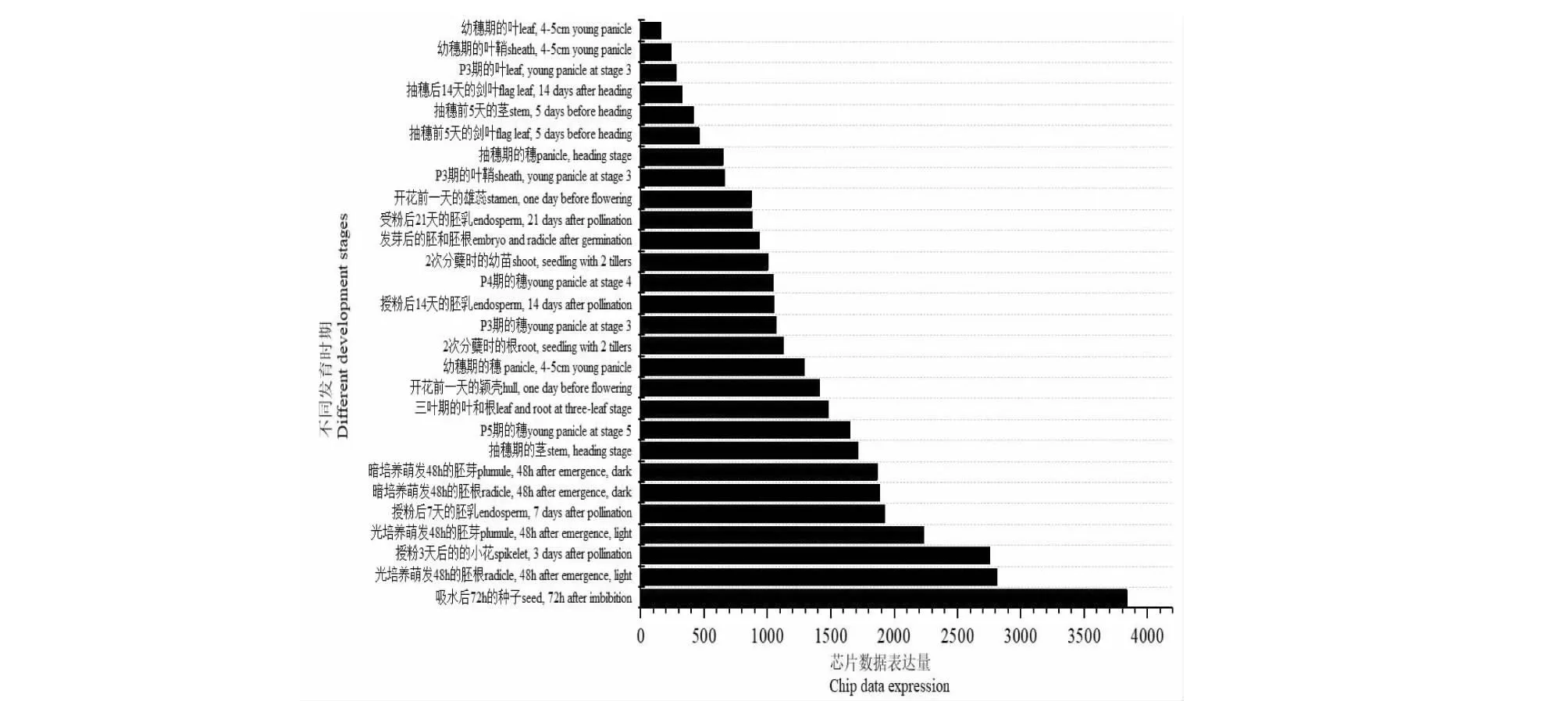
圖7 OsSMT2在水稻Minghui 63 中的芯片表達數據Fig.7 Genechip expression data of OsSMT2 in Minghui 63
2.3 水稻OsSMT2蛋白的同源性
OsSMT2蛋白具有甲基轉移酶結構域,位于OsSMT2蛋白質第107~283位氨基酸,還含有甾醇甲基轉移酶C末端結構域,位于OsSMT2蛋白質的第295~360位氨基酸(圖4)。通過MEME網站預測這些蛋白的保守基序發現,共有10個保守基序,其中Motif 1~9在10個物種中都含有,可見這9個基序為SMT2蛋白的核心基序(圖5)。
2.4 OsSMT2基因的系統發育樹和表達譜
從圖6看出,水稻中OsSMT2與藥用野生稻的ObSMT2親緣關系最近,與小米、高粱、玉米、大麥等主要糧食作物中SMT2的親緣關系較近。水稻OsSMT2基因在吸水后72 h種子中的表達量最高,在胚根、胚芽和小花中表達量相對較高,在葉中表達量最低(圖7)。表明,OsSMT2基因在水稻生長起始階段有重要的功能。
2.5 水稻OsSMT2基因啟動子元件
在水稻OsSMT2基因啟動子區域,除存在RNA聚合酶結合位點TATA-box、CAAT-box等基本調控元件外,還存在脅迫誘導元件、激素響應元件和光誘導元件(表1)。從OsSMT2基因啟動子區域具有的順式作用元件種類和數量推測,OsSMT2基因的表達模式受光和激素的誘導。啟動子中還存在4種脅迫響應元件,表明,OsSMT2基因可能與植物的脅迫響應有關。
2.6 OsSMT2基因共表達基因的GO富集和KEGG富集
共表達基因GO主要富集在微管、微管運動活性、基于微管的運動、驅動蛋白復合物、ATP結合、ATP酶活性、染色質結合、有絲分裂染色體濃縮、胞質分裂、微管細胞骨架組織中(圖8A)。KEGG富集9個代謝通路,包括DNA復制、泛素介導的蛋白水解、類固醇生物合成、吞噬、嘧啶代謝、內吞作用、嘌呤代謝、次生代謝產物的生物合成和代謝途徑等代謝通路中(圖8B)。
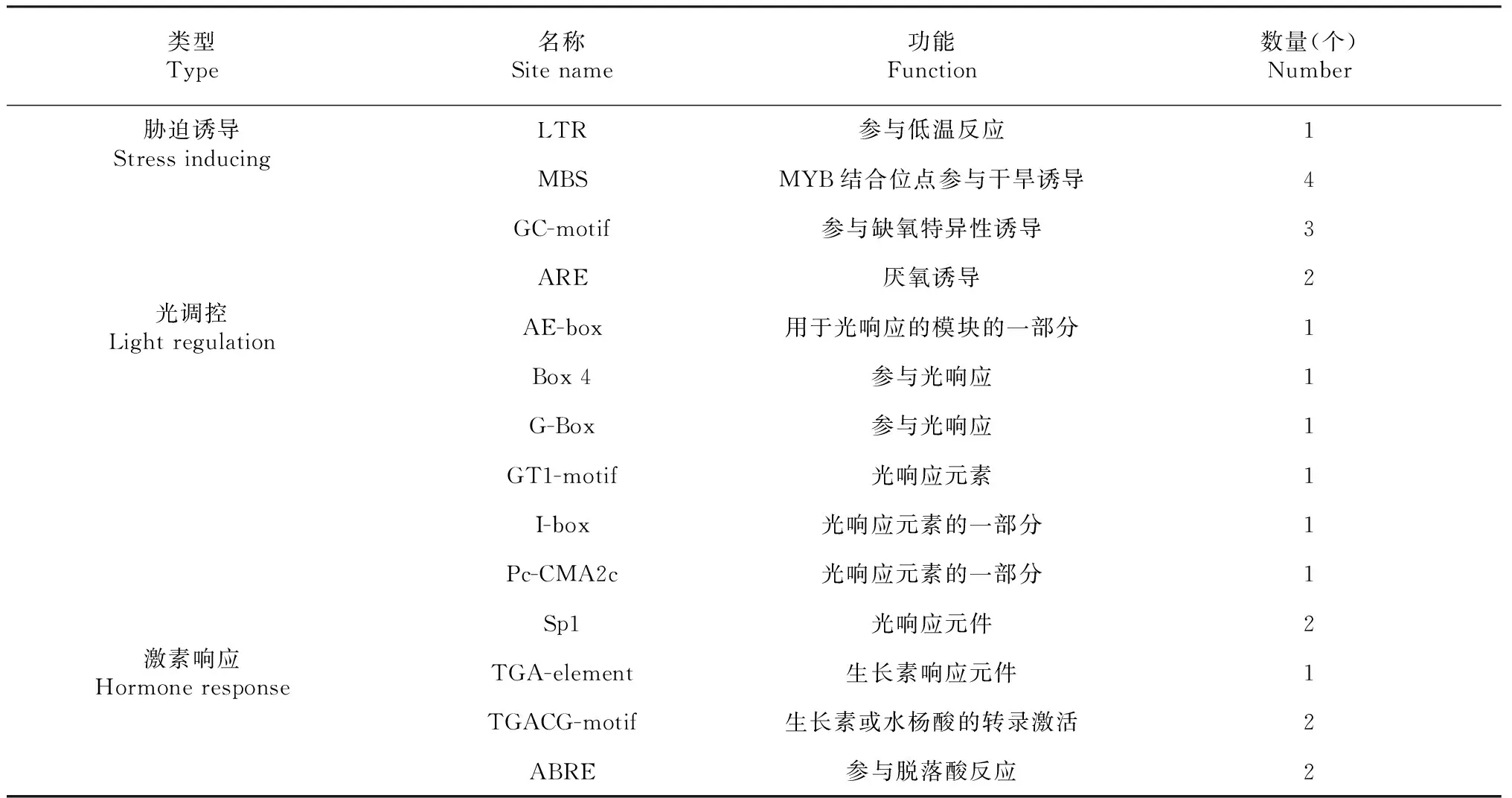
表1 OsSMT2的啟動子元件Table 1 Promoter elements of OsSMT2
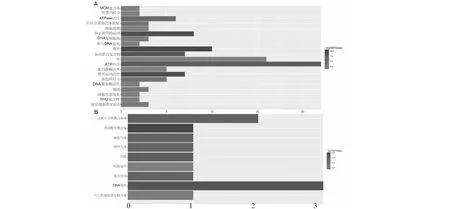
橫坐標表示富集到的基因數,A為 GO富集,B為 KEGG富集The abscissa indicates the number of genes enriched, A, GO enrichment, B, KEGG enrichment圖8 水稻OsSMT2基因共表達基因的富集Fig.8 Enrichment of co-expressed genes of OsSMT2
3 討 論
3.1 OsSMT2基因的功能相對保守
蛋白質的二級結構一般包含α螺旋、β轉角、延伸鏈和無規則卷曲,相鄰氨基酸殘基在氫鍵的作用下形成不同的空間結構,使蛋白質具有不同的活性和功能。因此,蛋白質結構的預測對推測其功能具有重要參考價值。水稻OsSMT2基因的二級結構以無規則卷曲為主,有42.70 %的氨基酸殘基參與。蛋白質的三級結構是在二級結構的基礎上發生折疊,使蛋白質具有更復雜的生物活性,并且能夠執行生物催化功能。對OsSMT2蛋白進行同源建模時發現,OsSMT2蛋白結構同樣以無規則卷曲為主,表明水稻OsSMT2蛋白中132個氨基酸殘基參與的α螺旋可能是起主要功能的區域。
水稻OsSMT2基因的功能結構域在不同物種中保守性較高,Motif 3、Motif 4、Motif 5、Motif 6、Motif 7、Motif 9處在甲基轉移酶結構域和甾醇甲基轉移酶C末端結構域中,是2種結構域的核心基序,SMT2基因在不同植物中的核心保守基序相同。因此,不同植物中SMT2基因的功能十分相似,水稻OsSMT2基因的研究結果也可為其他物種提供參考。水稻OsSMT2基因在主要糧食作物中同玉米、小麥、高粱的親緣關系最近,同大麥的親緣關系最遠,表明水稻OsSMT2基因的功能預測可以為玉米、小麥、高粱等糧食作物SMT2基因的研究提供參考。
3.2 OsSMT2基因參與水稻的抗寒和耐旱
在OsSMT2基因的啟動子區域存在多種響應元件,包括激素響應元件、光響應元件和脅迫響應元件。這些元件很可能會影響OsSMT2基因的轉錄激活,從而影響水稻中甾醇的含量以應對不同的外界脅迫。在大豆中,當環境溫度較低時,植物總甾醇的含量會降低,豆甾醇和β-谷甾醇的比值升高,當溫度升高時,大豆中植物總甾醇的水平會升高,其中油菜甾醇所占的比例與溫度變化成正相關,并且豆甾醇和β-谷甾醇的比例下降[17]。水稻OsSMT2基因的啟動子區域存在LTR元件參與低溫響應,因此低溫條件可能會影響水稻中OsSMT2基因的表達,進而影響水稻中甾醇的含量以抵抗低溫脅迫。同樣,在水稻OsSMT2基因的啟動子區域存在MBS元件,該元件可以參與干旱誘導,推測OsSMT2基因可能響應干旱脅迫,因為當植物遇到干旱脅迫時,植物甾醇的分子結構使得其能夠調節細胞膜的流動性和通透性,這種細胞膜性質的改變就可以抑制植物葉片中水分的流失,使植物在干旱條件下也能夠生存[18]。黃瓜在低氧脅迫下,甾醇合成途徑的產物-油菜素內酯可以促進黃瓜中糖類由葉片向根部的轉運,并提高黃瓜根部糖酵解代謝酶的活性[19]。水稻OsSMT2基因啟動子區域存在GC-motif 和ARE兩種厭氧誘導元件,因此OsSMT2基因可能會響應低氧脅迫。綜上所述,推測OsSMT2基因可能響應水稻低溫、干旱和低氧脅迫,可作為水稻抗寒和耐旱育種方面的候選基因。
3.3 OsSMT2基因影響水稻種子萌發
擬南芥AtSMT2-1基因在煙草中超表達后,與野生型相比,轉基因煙草中24-甲基甾醇的含量降低,抑制了細胞分裂,影響生長發育[20]。水稻OsSMT2基因共表達基因的GO富集和KEGG富集分析顯示,這些基因主要參與DNA的復制和細胞分裂的主要過程,如DNA復制的起始、DNA解旋酶的活性、細胞形態的發生等過程。OsSMT2基因表達模式分析顯示該基因在水稻種子萌發時的表達量最高,因此推測水稻OsSMT2基因可能影響水稻萌發過程中的細胞分裂,進而影響水稻的種子萌發率。
4 結 論
通過對水稻OsSMT2基因的生物信息學分析發現,水稻OsSMT2基因具有甲基轉移酶結構域和甾醇甲基轉移酶C末端結構域,參與水稻中甾醇的生物合成。該基因的啟動子區域含有低溫和干旱脅迫響應元件,因此OsSMT2基因可能參與水稻的非生物脅迫響應。共表達基因的富集分析顯示,該基因可能參與DNA復制和細胞分裂的主要過程。OsSMT2基因的表達模式分析表明,OsSMT2基因在吸水后72 h種子中的表達量最高,OsSMT2可能會影響水稻種子萌發和植物發育。研究結果為水稻的抗逆改良和種子萌發提供了理論參考和候選基因。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
