單粒精播對(duì)花生產(chǎn)量、光合特性及干物質(zhì)積累的影響
2020-05-21 03:33:29楊吉順齊林李尚霞
江蘇農(nóng)業(yè)科學(xué) 2020年6期
關(guān)鍵詞:產(chǎn)量
楊吉順 齊林 李尚霞
摘要:以高產(chǎn)大花生品種花育25號(hào)為試驗(yàn)材料,研究不同密度下單粒精播對(duì)花生產(chǎn)量、光合特性及干物質(zhì)積累的影響。結(jié)果表明,單粒精播處理產(chǎn)量顯著增加,與雙粒播種相比,單粒播種處理分別增產(chǎn)12.6%、19.8%、13.0%、11.4%、9.9%和10.0%。百果質(zhì)量、單株果數(shù)和飽果數(shù)表現(xiàn)出和產(chǎn)量基本相同的趨勢(shì)。單粒精播花生單葉光合速率,群體光合速率(CAP),莖、葉和莢果干物質(zhì)積累量均高于雙粒播種,而群體呼吸速率(CR)表現(xiàn)為隨著密度的增加而增加的趨勢(shì),但群體呼吸速率占群體光合速率的比例(CR/TCAP)表現(xiàn)為D1、D2和D3處理顯著低于雙粒播種,單粒精播處理間呈現(xiàn)出先減小后增大的趨勢(shì),D2處理最低。可見(jiàn),D2處理有利于減少花生植株呼吸消耗,更有利于花生干物質(zhì)的積累。
關(guān)鍵詞:花生;單粒精播;產(chǎn)量;光合特性;群體光合速率;群體呼吸速率;干物質(zhì)積累
中圖分類號(hào): S565.204? 文獻(xiàn)標(biāo)志碼: A? 文章編號(hào):1002-1302(2020)06-0064-04
花生(Arachis hypogaea L.)是我國(guó)重要的油料作物和經(jīng)濟(jì)作物,花生的持續(xù)增產(chǎn)對(duì)保障我國(guó)食用油脂安全具有重要意義[1]。目前,花生生產(chǎn)上仍然采用傳統(tǒng)雙粒播種為主的栽培模式,較單粒精播可有效減少缺苗斷壟的發(fā)生,而同穴雙株之間過(guò)窄的株距及較大的種植密度容易造成植株間競(jìng)爭(zhēng)加劇,個(gè)體發(fā)育受到限制,生育中期、后期群體環(huán)境惡化,導(dǎo)致葉片過(guò)早衰老,影響花生產(chǎn)量的進(jìn)一步提高[2-3]。另外,傳統(tǒng)雙粒播種用種量大,提高了花生的生產(chǎn)成本,大粒花生品種一般需要種子300~375 kg/hm2,中小粒型品種一般需要220~270 kg/hm2,全國(guó)用于做種的花生約為150萬(wàn)t/年,相當(dāng)于全國(guó)花生總產(chǎn)量的10%左右,種子投入量占農(nóng)資投入量的40%~50%[4],因此,在保持花生產(chǎn)量穩(wěn)定的條件下,減少用種量,可以實(shí)現(xiàn)降低成本、提高經(jīng)濟(jì)效益的目的。花生單粒精播是一項(xiàng)行之有效的節(jié)種、高產(chǎn)和高效的栽培技術(shù)。該技術(shù)改傳統(tǒng)的雙粒播種為單粒精播,減少穴播粒數(shù)地增加穴數(shù),不僅節(jié)約了用種量,而且有利于實(shí)現(xiàn)花生的機(jī)械化操作,王才斌等研究認(rèn)為,高產(chǎn)條件下,改雙粒播種種植為單粒精播種植,可以充分發(fā)揮單株生產(chǎn)力,更有利于群體高產(chǎn)[5]。通過(guò)大田試驗(yàn)和生產(chǎn)示范證明,與傳統(tǒng)雙粒播種相比,單粒精播技術(shù)在節(jié)種20%的前提下,仍可增產(chǎn)10%左右,生產(chǎn)成本卻大幅度下降[6]。馮燁等研究表明,單粒精播能夠有效協(xié)調(diào)根冠比,壯個(gè)體,強(qiáng)群體,充分發(fā)揮花生單株生產(chǎn)潛力,提高花生花后活性氧代謝水平,延緩花生后期的衰老進(jìn)程,增加莢果的干物質(zhì)積累[7-8]。本試驗(yàn)在前人的基礎(chǔ)上,采用高產(chǎn)大花生品種花育25號(hào),研究了不同密度單粒精播對(duì)花生產(chǎn)量、光合特性及干物質(zhì)積累特性的影響,并進(jìn)一步探明了單粒精播種植模式的最佳種植密度,以期為更好地推廣單粒精播種植模式提供理論基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)設(shè)計(jì)
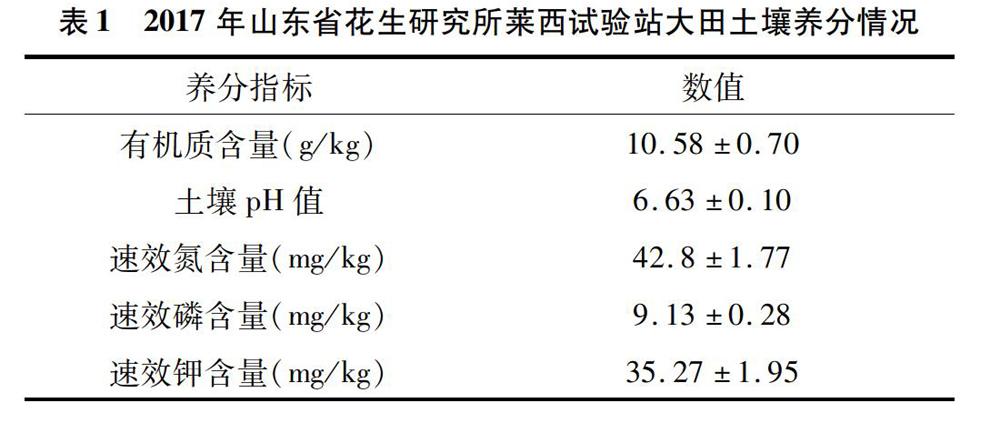
試驗(yàn)于2017年在山東省花生研究所萊西試驗(yàn)站大田內(nèi)進(jìn)行,供試花生品種為花育25號(hào),土壤為沙壤土,耕層(0~20 cm)土壤養(yǎng)分含量見(jiàn)表1。
試驗(yàn)采用大田種植方式,以雙粒播種(CK)為對(duì)照,單粒精播設(shè)置180 000(D1)、195 000(D2)、210 000(D3)、225 000(D4)、240 000(D5)、255 000株/hm2 (D6)6個(gè)密度處理,3次重復(fù),隨機(jī)區(qū)組排列。田間管理同一般大田。
1.2 測(cè)定項(xiàng)目及方法
1.2.1 干物質(zhì)積累的測(cè)定 從出苗后15 d開(kāi)始,每10 d取樣1次,每個(gè)小區(qū)取樣10株(5穴),將整株分為莖、葉和莢果3個(gè)部分,于烘箱內(nèi)105 ℃殺青0.5 h,然后將溫度調(diào)至80 ℃烘干并稱質(zhì)量。
1.2.2 單株葉片凈光合速率 用英國(guó)產(chǎn)的CIRAS-Ⅱ光合測(cè)定系統(tǒng)進(jìn)行測(cè)定。在花生花針期、結(jié)莢期與飽果期選取受光方向和生長(zhǎng)一致的葉片于典型晴天09:00—14:00測(cè)定主莖倒3葉的凈光合速率。
1.2.3 群體光合速率(CAP)和群體呼吸速率(CR) 群體光合速率的測(cè)定采用董樹(shù)亭等的方法[9]并略有改進(jìn)。于花針期、結(jié)莢期和飽果期選擇晴天無(wú)云天氣,光照度穩(wěn)定在1 400~1 600 μmol/(m2·s),用GXH-305型紅外線CO2分析儀在田間直接測(cè)定。同化箱長(zhǎng)1.0 m,寬0.9 m,高1.0 m,箱內(nèi)用風(fēng)扇攪拌氣體,框架外罩透明聚酯薄膜。采用閉路系統(tǒng),重復(fù)3次,每次測(cè)定60 s。用遮光布罩遮光后測(cè)定群體呼吸速率。在群體結(jié)構(gòu)相近的田地上,剪去與同化箱底大小相同面積地表上的植株后,測(cè)定土壤呼吸釋放的CO2,測(cè)定方法同群體光合速率(CAR),以修正群體光合速率和群體呼吸速率(CR)的測(cè)定值。計(jì)算群體呼吸速率占群體總光合速率(TCAR)的比例:CR/TCAP=CR/(CR+CAP)。
1.2.4 測(cè)產(chǎn) 成熟期每個(gè)處理取中間2行,每行連續(xù)取長(zhǎng)勢(shì)良好的10穴,自然風(fēng)干,用于室內(nèi)考種及收獲期農(nóng)藝性狀(單株果數(shù)、主莖高、側(cè)枝長(zhǎng)和分枝數(shù))的考察,收獲長(zhǎng)勢(shì)均勻具有代表性的2 m2自然晾干用于折算產(chǎn)量。
2 結(jié)果與分析
2.1 單粒精播對(duì)花生光合特性的影響
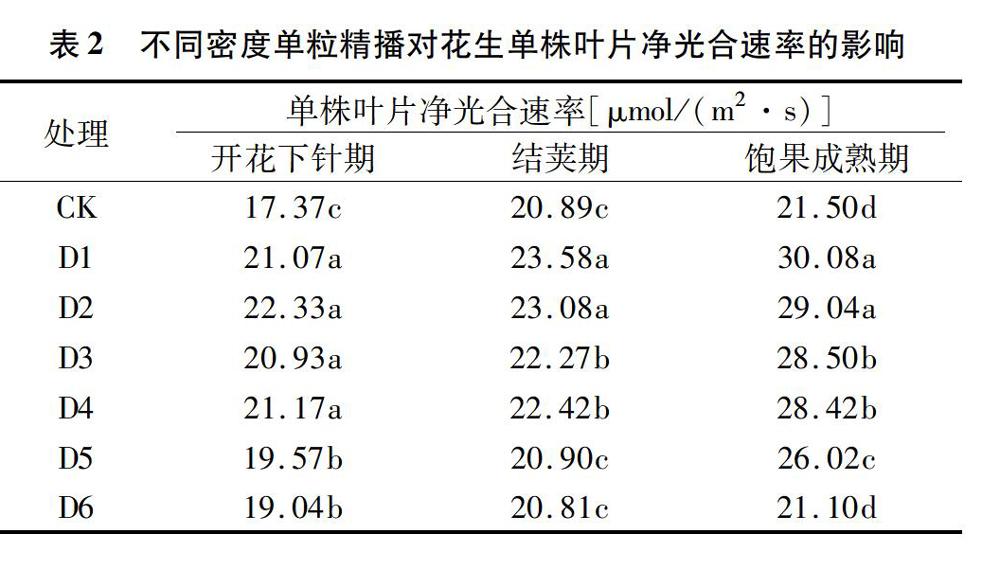
2.1.1 單粒精播對(duì)花生單株葉片凈光合速率的影響 如表2所示,隨著生育進(jìn)程的推進(jìn),各處理單株葉片凈光合速率均在飽果成熟期達(dá)到最高值。不同處理間相比,除飽果成熟期D6略低于CK外,其余各單粒播種處理均高于雙粒播種,而單粒播種處理凈光合速率隨著種植密度的增大呈下降趨勢(shì)。開(kāi)花下針期時(shí),D1~D4處理之間無(wú)顯著差異,但均顯著高于D5、D6與CK;結(jié)莢期與飽果成熟期則表現(xiàn)為D1、D2顯著高于D3、D4,又顯著高于D5、D6與CK,且不同處理間的差異有增大的趨勢(shì)。由此可見(jiàn),單粒精播對(duì)花生單株葉片凈光合速率的影響在生育前期較小,生育后期單粒精播更有利于花生凈光合速率維持在較高水平,從而為花生籽仁的充實(shí)提供物質(zhì)基礎(chǔ)。
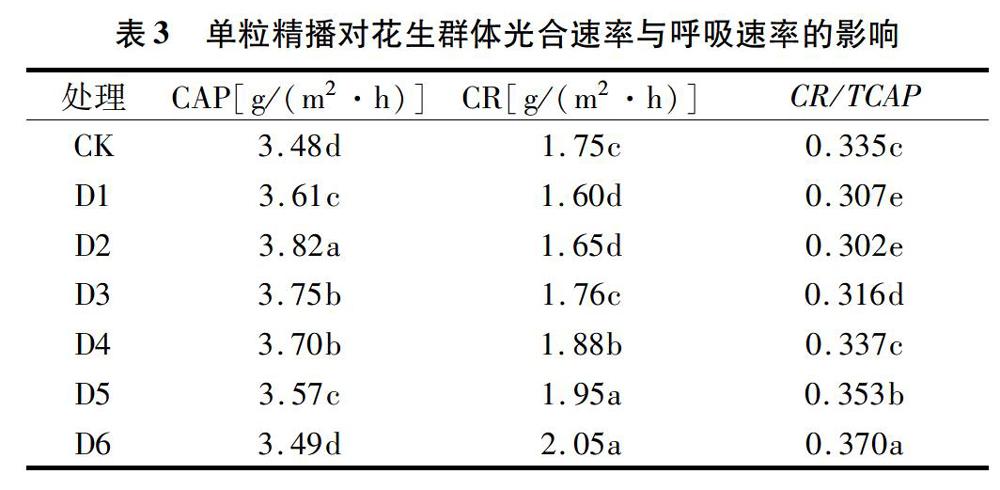
2.1.2 單粒精播對(duì)花生群體光合速率與呼吸速率的影響 如表3所示,群體光合速率(CAP)表現(xiàn)為單粒精播處理均高于雙粒播種,除D6處理以外均達(dá)到顯著水平,而單粒精播處理之間相比較,CAP隨著密度的增加而呈現(xiàn)先增大后減小的趨勢(shì),D2處理為最高。群體呼吸速率(CR)僅D1與D2處理低于雙粒播種,其余處理均高于雙粒播種,單粒精播處理的CR隨著密度的增加呈逐漸增大的趨勢(shì)。而CR/TCAP則表現(xiàn)為單粒精播D1、D2和D3處理顯著低于雙粒播種,單粒精播處理間呈現(xiàn)出先減小后增大的趨勢(shì),D2處理為最低。
2.2 單粒精播對(duì)花生干物質(zhì)積累量的影響
2.2.1 單粒精播對(duì)花生葉片干物質(zhì)積累量的影響 由圖1可知,葉片干物質(zhì)積累量隨著生育進(jìn)程的推進(jìn)呈先增加后減小的趨勢(shì),在播種后75 d達(dá)到最高值,之后呈降低趨勢(shì),單粒精播葉片干物質(zhì)積累量隨著密度的增加而呈降低趨勢(shì),除生育后期D6處理略低于雙粒播種以外,其余均高于雙粒播種。
2.2.2 單粒精播對(duì)花生莖稈干物質(zhì)積累量的影響 如圖2所示,莖稈干物質(zhì)積累量表現(xiàn)出與葉片干物質(zhì)積累量相似的趨勢(shì),除D6處理在生育后期略低于CK之外,單粒精播處理莖稈干物質(zhì)積累量均顯著高于雙粒播種,而單粒精播不同密度之間比較,隨著密度的增加,莖稈干物質(zhì)積累量呈降低的趨勢(shì)。
2.2.3 單粒精播對(duì)花生莢果干物質(zhì)積累量的影響 如圖3所示,花育25號(hào)莢果干物質(zhì)積累量隨著生育進(jìn)程的推進(jìn)而增加,而除D6處理干物質(zhì)積累量在花后55 d低于雙粒播種以外,單粒精播處理的莢果干物質(zhì)積累量均顯著高于雙粒播種。單粒精播處理的莢果干物質(zhì)積累量隨著密度的增加而降低。
2.3 單粒精播對(duì)花生產(chǎn)量及植株農(nóng)藝性狀的影響
如表4所示,各處理產(chǎn)量以雙粒播種最低,以D2產(chǎn)量最高,而密度超過(guò)195 000株/hm2的單粒精播處理的產(chǎn)量隨著密度的增大而呈減小的趨勢(shì)。與雙粒播種相比,單粒精播分別增產(chǎn)12.6%、197%、13.0%、11.3%、9.8%和10.1%。百果質(zhì)量、單株果數(shù)和百仁質(zhì)量表現(xiàn)出和產(chǎn)量基本相同的趨勢(shì)。主莖高表現(xiàn)為D4、D5與D6顯著高于其他處理,三者之間無(wú)顯著差異;側(cè)枝長(zhǎng)表現(xiàn)為D1、D2顯著高于其他處理,單粒精播處理之間的側(cè)枝長(zhǎng)隨著密度的增加而減短;分枝數(shù)與側(cè)枝長(zhǎng)表現(xiàn)出相似的趨勢(shì),單粒精播處理均多于雙粒播種。
3 討論
光合作用是產(chǎn)量形成的基礎(chǔ),作物干物質(zhì)90%以上來(lái)源于光合作用。許多研究認(rèn)為,作物生長(zhǎng)發(fā)育過(guò)程中光合生理特性與其籽粒產(chǎn)量關(guān)系密切[10]。本研究表明,單粒精播可有效提高花生倒3葉凈光合速率,尤其有利于花生生育后期的凈光合速率維持在較高的水平,從而有利于花生光合產(chǎn)物的形成,為花生產(chǎn)量的提高提供物質(zhì)基礎(chǔ)。作物的生產(chǎn)是一個(gè)種群的過(guò)程,而非個(gè)體表現(xiàn)[11],群體光合速率能準(zhǔn)確地描述單位土地面積上的光合能力,而且綜合了基因型效應(yīng)、葉片形態(tài)、冠層結(jié)構(gòu)等[12];作物產(chǎn)量與群體光合速率的關(guān)系較單葉光合速率更為緊密[13]。本研究表明,花生單粒精播處理群體光合速率(CAP)均高于雙粒播種,單粒精播不同密度之間表現(xiàn)為先增大后減小的趨勢(shì),D2處理最高,可見(jiàn)單粒精播有利于花生群體光合速率的提高,而密度過(guò)大會(huì)造成群體內(nèi)通風(fēng)透光性較差,不利于花生群體發(fā)育。而群體呼吸速率(CR)僅D1與D2處理低于雙粒播種,其余處理均高于雙粒播種,單粒精播處理CR隨著密度的增加呈逐漸增大的趨勢(shì);而群體呼吸速率占群體總光合速率的比例(CR/TCAP)則表現(xiàn)為單粒精播D1、D2和D3處理顯著低于雙粒播種,單粒精播處理間呈現(xiàn)出先減小后增大的趨勢(shì),D2處理最低,由此可見(jiàn),單粒精播有利于降低花生群體呼吸消耗,從而促使更多的光合產(chǎn)物用于莢果的干物質(zhì)積累,為花生高產(chǎn)提供前提。
干物質(zhì)是莢果形成的基礎(chǔ),同一品種在同一生態(tài)條件下的生育時(shí)間往往相對(duì)穩(wěn)定,產(chǎn)量主要取決于干物質(zhì)的積累速率[14-15]。本研究表明,單粒精播有利于花生植株莖稈、葉片和莢果的干物質(zhì)積累,為花生產(chǎn)量的提高提供物質(zhì)基礎(chǔ)。
單位面積作物高產(chǎn)主要靠增加群體密度,而高密度條件下群體與個(gè)體矛盾更加突出。密植群體形成高產(chǎn)的關(guān)鍵在于構(gòu)建合理的群體結(jié)構(gòu)[16]。雖然單粒精播花生單位面積植株數(shù)量有所降低,但個(gè)體發(fā)育較好,充分發(fā)揮了單株增產(chǎn)潛力[17],單位面積果數(shù)顯著高于雙粒穴播。孫彥浩等認(rèn)為,花生要進(jìn)一步獲得高產(chǎn),應(yīng)在增加群體果數(shù)的同時(shí),提高雙仁果率和飽果率[18]。本研究表明,單粒精播能顯著增加莢果產(chǎn)量,其中D2處理增產(chǎn)效果最佳,與對(duì)照相比增產(chǎn)達(dá)19.7%,增產(chǎn)的原因主要是增加了單株結(jié)果數(shù)從而增加了群體果數(shù)。
4 結(jié)論
作物的生產(chǎn)是一個(gè)種群的過(guò)程,而非個(gè)體表現(xiàn),要獲得高產(chǎn)穩(wěn)產(chǎn),就必須使個(gè)體、群體和環(huán)境相協(xié)調(diào)達(dá)到最優(yōu)化[19]。趙雙進(jìn)等認(rèn)為,作物群體產(chǎn)量最高時(shí),其單株在田間的分布應(yīng)該處于最佳狀態(tài)[20]。以上結(jié)果表明,單粒精播技術(shù)可促進(jìn)花生植株個(gè)體發(fā)育,充分發(fā)揮花生個(gè)體潛力,使花生群體與個(gè)體得到協(xié)調(diào)發(fā)展,促進(jìn)花生產(chǎn)量的提高。在本試驗(yàn)條件下,花育25號(hào)以單粒精播 195 000株/hm2 密度為最佳種植方式。
參考文獻(xiàn):
[1]萬(wàn)書(shū)波. 山東花生六十年[M]. 北京:中國(guó)農(nóng)業(yè)科學(xué)技術(shù)出版社,2009.
[2]李安東,任衛(wèi)國(guó),王才斌,等. 花生單粒精播高產(chǎn)栽培生育特點(diǎn)及配套技術(shù)研究[J]. 花生學(xué)報(bào),2004,33(2):17-22.
[3]梁曉艷,郭 峰,張佳蕾,等. 單粒精播對(duì)花生冠層微環(huán)境、光合特性及產(chǎn)量的影響[J]. 應(yīng)用生態(tài)學(xué)報(bào),2015,26(12):3700-3706.
[4]吳繼華,王 偉,張秀云. 河南麥套花生可持續(xù)發(fā)展思路與技術(shù)對(duì)策[J]. 農(nóng)業(yè)科技通訊,2008(1):22-25.
[5]王才斌,成 波,遲玉成,等. 高產(chǎn)花生單粒植群體密度研究[J]. 花生科技,1996(3):17-19.
[6]邵長(zhǎng)亮. 花生單粒精播節(jié)種高產(chǎn)理論與技術(shù)研究[D]. 青島:青島農(nóng)業(yè)大學(xué),2005.
[7]馮 燁,郭 峰,李寶龍,等. 單粒精播對(duì)花生根系生長(zhǎng)、根冠比和產(chǎn)量的影響[J]. 作物學(xué)報(bào),2013,39(12):2228-2237.
[8]馮 燁,李寶龍,郭 峰,等. 單粒精播對(duì)花生活性氧代謝、干物質(zhì)積累和產(chǎn)量的影響[J]. 山東農(nóng)業(yè)科學(xué),2013,45(8):42-46.
[9]董樹(shù)亭,高榮岐,胡昌浩,等. 玉米花粒期群體光合性能與高產(chǎn)潛力研究[J]. 作物學(xué)報(bào),1997,23(3):318-325.
[10]Jiang G M,Hao N B,Bai K Z,et al. Chain correlation between variables of gas exchange and yield potential in different winter wheat cultivars[J]. Photosynthetica,2000,38(2):227-232.
[11]趙松嶺,李鳳民,張大勇. 作物生產(chǎn)是一個(gè)種群過(guò)程[J]. 生態(tài)學(xué)報(bào),1997,17(1):100-104.
[12]Wall G W,Kanemasu E T. Carbon dioxide exchange rate in wheat canopies. Part Ⅰ. Influence of canopy geometry on trends in leaf area index,light interception and instantaneous exchange rates[J]. Agricultural and Forest Meteorology,1990,49:81-102.
[13]Lin Z F,Ehleringer J. Photosynthetic characteristics of Amaranthus tricolor,a C4 tropical leafy vegetable[J]. Photosynthesis Research,1983,4:171-178.
[14]杜 紅,閆凌云,路紅衛(wèi),等. 高產(chǎn)花生品種干物質(zhì)生產(chǎn)對(duì)產(chǎn)量的影響[J]. 中國(guó)農(nóng)學(xué)通報(bào),2005,21(8):104-106.
[15]楊偉強(qiáng),宋文武,鞠 倩,等. 不同類型花生品種(系)干物質(zhì)積累特性研究[J]. 山東農(nóng)業(yè)科學(xué),2009(1):47-49.
[16]Maddonni G,Chelle M,Drouet J L,et al. Light interception of contrasting azimuth canopies under square and rectangular plant spatial distributions:simulations and crop measurements[J]. Field Crops Research,2001,70(1):1-13.
[17]趙長(zhǎng)星,邵長(zhǎng)亮,王月福,等. 單粒精播模式下種植密度對(duì)花生群體生態(tài)特征及產(chǎn)量的影響[J]. 農(nóng)學(xué)學(xué)報(bào),2013,3(2):1-5.
[18]孫彥浩,劉恩鴻,隋清衛(wèi),等. 花生畝產(chǎn)千斤高產(chǎn)因素結(jié)構(gòu)與群體動(dòng)態(tài)的研究[J]. 中國(guó)農(nóng)業(yè)科學(xué),1982(1):71-75.
[19]金 劍,劉曉冰,王光華,等. 大豆高產(chǎn)群體的生態(tài)生理特征[J]. 中國(guó)油料作物學(xué)報(bào),2003,25(3):111-116.
[20]趙雙進(jìn),張孟臣,楊春燕,等. 栽培因子對(duì)大豆生長(zhǎng)發(fā)育及群體產(chǎn)量的影響——Ⅰ.播期、密度、行株距(配置方式)對(duì)產(chǎn)量的影響[J]. 中國(guó)油料作物學(xué)報(bào),2002,24(4):31-34.萬(wàn)淑紅,田應(yīng)兵,許昌雨,等. 氮素調(diào)控對(duì)水稻黃華占生長(zhǎng)發(fā)育及產(chǎn)量的影響[J]. 江蘇農(nóng)業(yè)科學(xué),2020,48(6):68-72.
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當(dāng)代水產(chǎn)(2021年10期)2021-12-05 16:31:48
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
今日農(nóng)業(yè)(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中國(guó)果業(yè)信息(2019年10期)2019-11-13 01:21:34
中國(guó)化肥信息(2019年2期)2019-01-18 15:24:35
中國(guó)化肥信息(2019年1期)2019-01-17 21:31:12
中國(guó)化肥信息(2019年4期)2019-01-17 18:47:06
