選擇性剪接與細胞凋亡
2020-05-25 08:24:36黎權文李全忠
世界最新醫學信息文摘 2020年36期
黎權文,李全忠
(桂林醫學院附屬醫院心血管內科,廣西 桂林)
0 引言
凋亡定義為單個細胞破碎成膜結合顆粒,表現為DNA 片段化、胞質和核濃縮、染色質濃縮、凋亡小體形成,然后被特殊的吞噬細胞如巨噬細胞和嗜中性粒細胞吞噬。凋亡有助于機體去除感染、受損、老化或危險的細胞,從而限制周圍組織的破壞,并維持組織的動態平衡和器官的正常發育。凋亡的紊亂與自身免疫、慢性炎癥性疾病、腫瘤直接相關。
真核生物中,選擇性剪接(alternative splicing,AS)發生在基因轉錄后,使單個基因產生一個以上的轉錄本[1]。通過AS,同一基因產生的不同的mRNA 轉錄本,經翻譯形成的蛋白質可能在結構、功能、活性等方面存在明顯差異[2]。這種差異,造就了蛋白質的多樣性及物種的復雜性。對AS 的研究逐漸深入,已經從單個剪接事件發展到描述全局的剪接網,著重強調它們如何進行分子協調。現已確定,AS 有助于細胞分化、組織和器官發育[3]。而全基因組研究估計人類90-95%的基因經歷不同程度的AS[3,4]。同時,前體mRNA 中順式作用的序列元件、反式作用的剪接因子或剪接體其他成分的突變引起人類多種疾病,進一步凸顯了AS 的生物學重要性[5]。本綜述介紹AS 生理學基礎及調控機制,同時還探討了AS 事件與凋亡的關系。
1 AS 的生物學基礎
真核生物基因序列中含有多個外顯子及內含子,然而成熟的mRNA 只含有外顯子序列信息,這種將前體mRNA 剪接加工去除內含子形成成熟mRNA 的現象稱為選擇性剪接(alternative splicing,AS)。剪接位點選擇的改變直接影響遺傳序列,導致蛋白質一級結構序列改變,最終將可能影響蛋白質的變構調節、酶活性或配體結合。剪接因子識別并結合順式作用調控序列,是建立和維持AS 的基本機制。這些順式作用調控序列包括外顯子增強子、外顯子沉默子、內含子增強子和內含子沉默子。剪接因子與順式作用調控序列的識別機制復雜而精密,大體上分為RNA-蛋白質識別、RNA-RNA 識別和基因調控的特異性位點識別。
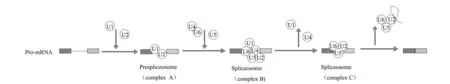
剪接體是一種高度動態的核糖核蛋白復合體,由小的核糖核蛋白顆粒U1、U2、U4/U6 和U5 組成。它精密的組裝與分解直接影響剪接作用,是AS 的執行者[6]。在識別剪接位點后,剪接體集合并組裝成剪接復合體完成AS,其機制主要包括:(1)剪接體U1、U2 與前體mRNA 上的剪接位點結合,并形成前剪接體復合體A[7];(2)snRNP 將U4/U6、U5 轉運至剪接位點,與前剪接體復合體A 形成前剪接體復合體B[8,9];(3)U1、U4 脫離,U2、U5、U6 形成具有催化活性的剪接體復合物C[10,11],最終執行AS(圖1)。
2 AS 的調控
AS 受順式作用調控序列和反式作用因子。反式作用因子包括富含絲氨酸/精氨酸(SR)的蛋白家族、異源核糖核蛋白(hnRNPs)家族和組織特異性RNA 結合蛋白。反式作用因子識別并結合順式作用調控序列,促進或抑制AS,是AS 的基本調控方式[12]。這些順式作用調控序列和反式作用因子的多重作用和作用機理已得到廣泛研究,但是實際上更為復雜。因為剪接與轉錄相關,并且調節轉錄的因子也直接影響AS[13]。其次,基因的轉錄與前體mRNA 的AS 存在時間及空間上的交錯。真核生物基因轉錄編碼形成mRNA 由RNA 聚合酶II(RNA polymerase II,RNAPII)執行。進一步研究表明,RNAPII 在AS 中至關重要,其羧基端結構域對剪接因子聚集到轉錄位置是必不可少的[14]。RNAPII 的延伸速率直接影響轉錄過程中新生前體mRNA 的剪接位點和順式作用調控序列出現的速度。RNAPII 延伸減慢或暫停,有利于促進外顯子包含;反之,則有利于促進外顯子跳躍。最后,在基因轉錄過程中,核小體不可避免的阻礙RNAPII 的延伸,這種方式有利于剪接因子的識別并促進外顯子包含[15]。組蛋白高乙酰化與共轉錄剪接調控相關,其通過增加或減少剪接因子的募集而調控AS;同時組蛋白高乙酰化可導致局部RNAPII 延伸率增加,促進外顯子跳躍。組蛋白標記也可以對AS 造成直接的影響,因為某些組蛋白修飾會通過染色質結合蛋白募集剪接因子[16]。甲基結合域蛋白募集組蛋白修飾酶來改變周圍的染色質,干擾RNAPII的延伸率。DNA 甲基化可能通過改變染色質或RNAPII 的延伸率影響AS。這些說明,表觀遺傳學也直接影響AS。
3 AS 與凋亡
3.1 剪接體核心成分與凋亡
mRNA 前體加工因子(pre-mRNA processing factor,PRPF)是剪接體核心成分。PRPF8 氨基酸序列437-770 結構域結合U5[17],同時羧基端激活U4/U6 的解旋酶[18]。PRPF8 突變引起細胞凋亡導致神經變性[19]。PRPF4 是U4/U6 的核心蛋白,能通過p38 MAPK 信號通路抑制乳腺癌細胞凋亡[20]。PRPF3 與PRPF4相互作用,共同維持snRNP 結構穩定[21]。Comitato A 等研究發現,突變的PRPF3 聚集在核仁內大的聚集物中,影響了SRSF2 的定位,觸發光感受器細胞的凋亡[22]。PRPF31 的突變促進RHO 基因外顯子3 跳過,降低視紫紅質的表達,并導致視紫紅質陽性的視網膜細胞凋亡[23]。PRPF40B 定位于富含剪接因子的核斑點,與剪接因子SF1 和U2AF65 結合,調節不同的剪接事件。PRPF40B 的缺失促進Fas 外顯子6 的包含,導致長促凋亡Fas 亞型增加,膜結合Fas 受體增加,促進細胞凋亡[24]。
3.2 剪接因子與凋亡
剪接因子主要由SR 蛋白家族和hnRNPs 蛋白家族組成。大多數SR 蛋白與剪接體結合促進剪接體識別外顯子,從而促進AS;而hnRNPs 與前體mRNA 的剪接位點結合,阻止剪接體識別剪接位點。SR 蛋白經常與hnRNPs 結合,以濃度依賴的方式拮抗hnRNPs 的作用[25]。剪接因子3b 亞基1(splicing factor 3b subunit 1,SF3B1)是U2 復合體的一個組成成分。Zhang Q 等研究發現,SF3B1 是Tap73 的關鍵剪接因子,抑制SF3B1 表達能提高Tap73/DNp73 比列,并促進宮頸癌細胞凋亡[26]。hnRNA 蛋白K(Heterogeneous nuclear ribonucleo protein K ,hnRNPK)是MRPL33 前體mRNA 的關鍵剪接調節因子,促進MRPL33 外顯子3 包含,導致MRPL33-L 表達增加,抑制結直腸癌細胞凋亡[27]。富含絲氨酸/精氨酸的剪接因子/剪接因子2(Serine/arginine rich splicing factor/Splicing factor 2 ,ASF1/SF2)是序列特異性剪接因子。Iwanaga N 等使用促凋亡刺激物處理U937 細胞發現,caspase-2 前體mRNA 外顯子9 包含增加,導致caspase-2S/caspase-2L 的比例增加[28]。Jiang ZH 等研究表明ASF1/SF2、剪接因子SC35 促進caspase-2 的外顯子9 跳躍,而hnRNP A1 促進外顯子9 包含[29]。這表明ASF1/SF2、剪接因子SC35、hnRNP A1共同調節caspase-2 的AS,調控細胞凋亡。
3.3 凋亡基因的AS 與凋亡
凋亡是一種程序性細胞細胞,凋亡的激活表現為促凋亡基因的增加和抗凋亡基因的減少以及caspase 的激活,是一種規律的級聯反應。然而,許多凋亡基因的AS 形成不同的亞型,它們在功能上常表現出拮抗作用。
Bcl-x 基因含有3 個外顯子,在外顯子2 中具有兩個5,剪接位點。Bcl-x 基因的AS 中,選擇了近端5,剪接位點,則會產生長的Bcl-xL 亞型;而選擇了遠端5,剪接,則會產生短的Bcl-xS 亞型。Bcl-xL 具有233 個氨基酸,含有BH1、BH2、BH3 和BH4 結構域。Bcl-xL 通過BH1、BH2 結構域與線粒體膜結合,誘導與線粒體膜結合的Bax 向細胞質轉移,或者直接與Bax 結合,阻止了tBid 與Bax 結合,防止線粒體外膜持續開發以及細胞色素c 的釋放[30]。因此,Bcl-xL 通過抑制線粒體外膜持續開發來抑制細胞凋亡[31]。而Bcl-xS 與Bcl-xL 相比,缺少了63 個氨基酸區段,該區段包含保守的BH1 和BH2 結構域。Bcl-xS 與Bcl-xL 形成異源二聚體,抑制Bcl-xL 的抗凋亡作用。多種剪接因子和信號通路影響Bcl-xL/Bcl-xS 的剪接比,包括SR 蛋白、hnRNP、RNA 結合蛋白和轉錄因子。PUMA 有PUMA-α,-β,-δ 和-γ 四種不同的 剪 接 變 體[32]。PUMA-α 和-β 都 包 含BH3 域,而PUMA-γ 和PUMA-δ 沒有BH3 域。進一步研究表明,PUMA-α 和-β 通過結合BAX 的BH3 結構域,使線粒體外膜通透性改變,釋放細胞色素c,從而介導線粒體誘導的細胞凋亡[33]。Bnip3 是一種BH3-only蛋白。Bnip3 基因的AS 產生兩種剪接變體,分別為Bnip3FL 和Bnip3Δex3。Bnip3FL 同時具有BH3 和羧基末端跨膜結構域,而Bnip3Δex3 兩者均缺失[34]。Bnip3FL 蛋白結構中含有羧基末端跨膜結構域,是Bnip3 進入線粒體執行促凋亡生物功能的基礎[35]。Bnip3Δex3 中缺少羧基末端跨膜結構域,表明該剪接體無法將自身插入線粒體膜中。但是Bnip3Δex3 與Bnip3FL 結合形成同源二聚體,防止Bnip3FL 與線粒體膜結合[34]。凋亡基因的AS 受多種因素調控,包括細胞內及細胞外環境因素,其調控機理精密而復雜。以前,由于復雜的分析,較高的成本以及在較舊的、較不敏感的技術中發現的假陽性結果,凋亡基因AS 調控的研究受到限制。但是,隨著測序成本的降低和二代測序的出現,凋亡基因AS 的調控機制將得到更深入的認識。
4 總結
AS 是真核生物基因表達調控的關鍵部分,受多種因素調節,參與真核生物的生長、發育、進化。同時,細胞在各種病理或應激狀態下,AS 作用導致基因組表達發生差異性改變以維持細胞的穩態。然而,AS 的調控不僅僅是單一的調控機制,而是復雜精細的調控網。凋亡是在各種病理或應激狀態下以適應機體需要而執行的程序性細胞死亡,其與AS 相關的事件足以證明AS 與凋亡直接相關。因此,有必要進一步闡明剪接調控網在凋亡中的級聯改變。深入了解凋亡的剪接調控網,將有可能開發出新的治療靶標,為新的藥物研發提供新的思路。
