基于ITS和RLKs序列的苦蕎種質(zhì)資源遺傳多樣性分析
2020-06-01 07:47:32張久盤常克勤楊崇慶穆蘭海杜燕萍
南方農(nóng)業(yè)·下旬 2020年1期
關鍵詞:苦蕎
張久盤 常克勤 楊崇慶 穆蘭海 杜燕萍

摘 要 為明確苦蕎資源的遺傳多樣性水平及遺傳關系,對45份苦蕎地方品種的ITS和RLKs序列進行基因測序和序列比較分析。結(jié)果顯示,苦蕎ITS和RLKs擴增序列較為保守,基因多態(tài)位點數(shù)分別占10.6%和7.7%。云貴川地區(qū)苦蕎材料的遺傳多樣性最豐富,陜西、山西及寧夏地區(qū)次之,甘肅及內(nèi)蒙古地區(qū)最低。四川和貴州地區(qū)的苦蕎品種均能夠單獨聚為一類,基于ITS序列構(gòu)建的NJ樹中,陜西地區(qū)以北的苦蕎品種未形成明顯的分支,而云南地區(qū)的苦蕎穿插在不同的分支中;基于RLKs基因構(gòu)建的NJ樹中,同一地區(qū)的苦蕎材料較為明顯地形成地域聚類,但自展值均較低。
關鍵詞:苦蕎;遺傳多樣性;ITS;RLKs
蕎麥(Fagopyrum tataricum)又稱烏麥、三角麥、花蕎、蕎子,屬于蓼科(Polygnaceae)蕎麥屬(Fagopyrum),起源于我國西南地區(qū)[1]。目前,發(fā)現(xiàn)蕎麥共有28個種、亞種和變種,而蕎麥栽培種僅有甜蕎和苦蕎,其生物學特征及栽培適宜區(qū)域均有所不同[2]。蕎麥營養(yǎng)成分均衡,富含賴氨酸[3-5]及黃酮類化合物,具有降低“三高”、軟化血管、保護視力和預防腦溢血的作用[6-7],是藥食兼用的保健和功能產(chǎn)品。由于蕎麥具有抗逆性強、適應性廣、耐瘠薄耐粗放等優(yōu)點,成為生產(chǎn)條件較差的地區(qū)大量種植的糧食作物。
過去國內(nèi)外學者多采用形態(tài)學標記、細胞學標記、種子蛋白標記和同工酶標記等方法對蕎麥親緣關系及遺傳多樣性進行分析,但分子標記技術已成為近年來最主要的種質(zhì)資源遺傳多樣性分析手段,國內(nèi)外廣泛使用RAPD、AFLP、SSR等分子標記來評價蕎麥遺傳多樣性。胡亞妮等[8]基于ITS和ndhF-rpl32序列構(gòu)建的進化樹將71份蕎麥材料分為大粒組和小粒組,而ndhF-rpl32序列構(gòu)建的進化樹還能區(qū)分栽培甜蕎和野生甜蕎,具有更好的聚類效果。李敏等[9]利用PAL基因?qū)?7份苦蕎地方品種分為7個類群,分類與地理類群無關,僅來源于西藏的5份材料單獨聚為一枝;PAL基因在苦蕎中遺傳較穩(wěn)定,多數(shù)材料之間變化差異較小,在西藏部分材料中可能存在突變的熱點區(qū)。梁成剛等[10]研究表明蕎麥屬MAPK基因序列高度保守,通過聚類分析發(fā)現(xiàn)野生甜蕎與左貢野蕎被聚為一類,野生苦蕎、毛野蕎、大野蕎、金蕎麥、細柄野蕎和硬枝萬年蕎被聚為一大類。
本研究以45份苦蕎為試驗材料,通過PCR直接測序的方法獲得ITS和RLKs基因序列,進行序列差異和聚類分析,闡明苦蕎地方品種的遺傳多樣性水平,旨在揭示我國苦蕎種質(zhì)資源間的遺傳關系,為苦蕎種質(zhì)資源的收集、保護、利用提供依據(jù)。
1 材料與方法
1.1 試驗材料
試驗材料為苦蕎地方品種,共45份,分別收集自四川、貴州、云南及寧夏等8個省份(自治區(qū)),供試材料的代碼、名稱及來源見表1。
1.2 試驗方法
1.2.1 DNA提取
采集供試材料3~4片葉齡時的嫩葉0.5 g,利用Tiangen公司植物基因組DNA提取試劑盒(DP305)提取樣本DNA,用1%瓊脂糖凝膠電泳和分光光度計檢測DNA的純度和濃度。
1.2.2 PCR擴增、測序
用于擴增目的片段ITS和RLKs的引物序列如下。
ITS: F5'-TCCTCCGCTTATTGATATGC-3',R5'-TCCGTAGGTGAACCTGCGG-3'。RLKs:F5'-GT-GTTGCTCACCAGTTGGATT-3',R5'-TTC-TATCCAGTGGGGTGACTG-3'。引物序列由北京擎科生物技術有限公司合成。
PCR反應體積20 uL,包括2×Taq PCR預混試劑Ⅱ2.8 uL、10 mM引物各0.6 uL、DNA模板1 uL、ddH2O 15 uL。PCR反應程序:95 ℃預變性3 min;94℃ 變性30 s,54 ℃/56 ℃(ITS/RLKs)退火30 s,72 ℃延伸90 s,共30個循環(huán);72 ℃延伸10 min。PCR擴增產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳檢測后送至北京擎科生物有限公司進行測序。
1.2.3 數(shù)據(jù)分析
通過Chromas 1.45軟件獲得原始序列數(shù)據(jù),用ClustalX 1.83程序?qū)y得的DNA序列進行比對,并加以人工校對,用DnaSP 5.1軟件[11]統(tǒng)計多態(tài)位點。用Modeltest 3.06軟件[12]選擇2個基因片段核苷酸的最佳替換模型,根據(jù)最佳替換模型運用Mega 5.0軟件[13]進行遺傳多樣性指數(shù)、差異位點的統(tǒng)計分析及遺傳距離的計算。基于Jukes-Cantor距離法,進行1 000次自展重復檢測支持率,構(gòu)建Neighbor-Joining(鄰接法)系統(tǒng)發(fā)育樹。
2 結(jié)果與分析
2.1 ITS和RLKs序列特征及遺傳多樣性參數(shù)分析
經(jīng)PCR擴增、測序分別獲得43條ITS和45條RLKs同源序列。經(jīng)過DNA序列比對,切除兩端引物序列后,獲得ITS和RLKs同源序列片段長度分別為539 bp和815 bp。利用MEGA 5.0和DnaSP 5.1軟件對ITS、RLKs序列數(shù)據(jù)進行分析,在ITS序列539個位點中,變異位點為57個,基因多態(tài)位點百分比為10.6%;在815 bp的RLKs序列矩陣中,變異位點63個,基因多態(tài)位點百分比為7.7%。苦蕎各地方品種兩序列的變異信息及遺傳多樣性參數(shù)見表2。
不同地區(qū)的苦蕎品種ITS序列的核苷酸多樣度π的變化范圍為0.001 9~0.010 5,平均雜合度θ的變化范圍為0.001 9~0.015 1;RLKs基因的核苷酸多樣度π的變化范圍為0.002 2~0.009 0,平均雜合度θ的變化范圍為0.002 4~0.012 2。從ITS和RLKs序列的π值和θ值看出,貴州、四川和云南地區(qū)苦蕎材料的遺傳多樣性最豐富,陜西、山西及寧夏地區(qū)次之,甘肅及內(nèi)蒙古地區(qū)的苦蕎材料的遺傳多樣性最低。
2.2 基于ITS和RLKs序列的苦蕎組內(nèi)及組間遺傳距離分析
利用MEGA 5.0軟件,基于Jukes-Cantor距離模型分析ITS和RLKs序列數(shù)據(jù),將來源相同省(自治區(qū))的苦蕎品種進行分組,得到不同地方來源苦蕎的組內(nèi)平均遺傳距離(表3)及組間平均遺傳距離(表4)。
由表3和表4可知,基于ITS序列的組內(nèi)遺傳距離為0.002~0.011,貴州、四川及陜西苦蕎材料組內(nèi)遺傳距離相對較大;內(nèi)蒙古與云南、四川地區(qū)的苦蕎材料組間遺傳距離較大,陜西與云南地區(qū)、內(nèi)蒙古與貴州地區(qū)的苦蕎材料組間遺傳距離次之。基于RLKs序列組內(nèi)遺傳距離為0.002~0.010,貴州、四川及云南苦蕎材料組內(nèi)遺傳距離相對較大;內(nèi)蒙古與云南地區(qū)、四川地區(qū)的苦蕎材料組間遺傳距離較大,陜西與云南地區(qū)的苦蕎材料組間遺傳距離次之。
從結(jié)果來看,無論是相同地方的苦蕎品種間還是不同地方的苦蕎品種間,遺傳距離基本處于同一水平。北方產(chǎn)區(qū)的苦蕎資源變異較小,而南方產(chǎn)區(qū)苦蕎變異較為豐富,且北方產(chǎn)區(qū)苦蕎與南方產(chǎn)區(qū)苦蕎組間遺傳距離相對較大。
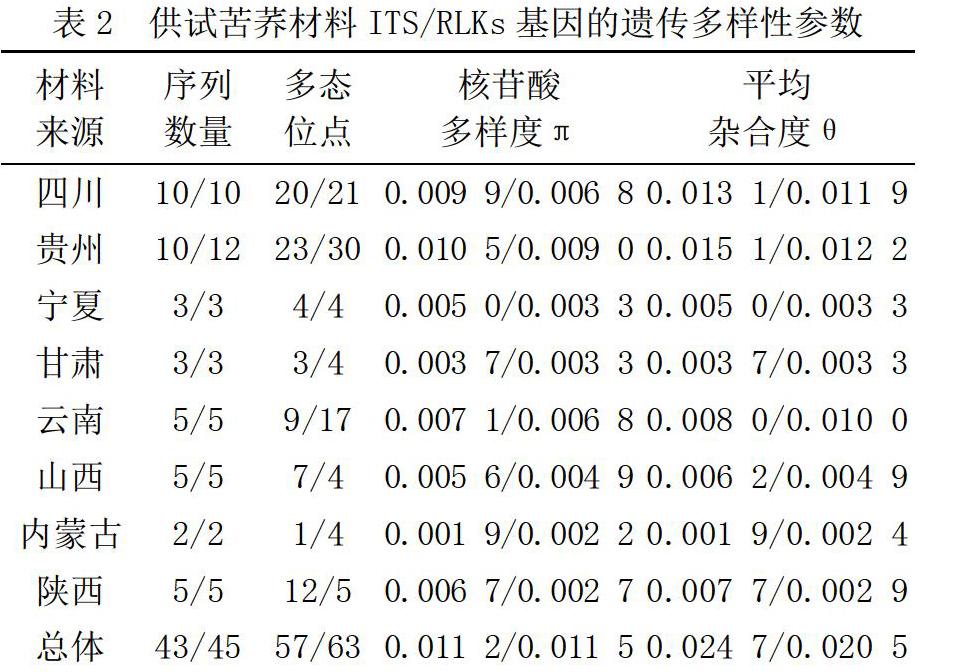
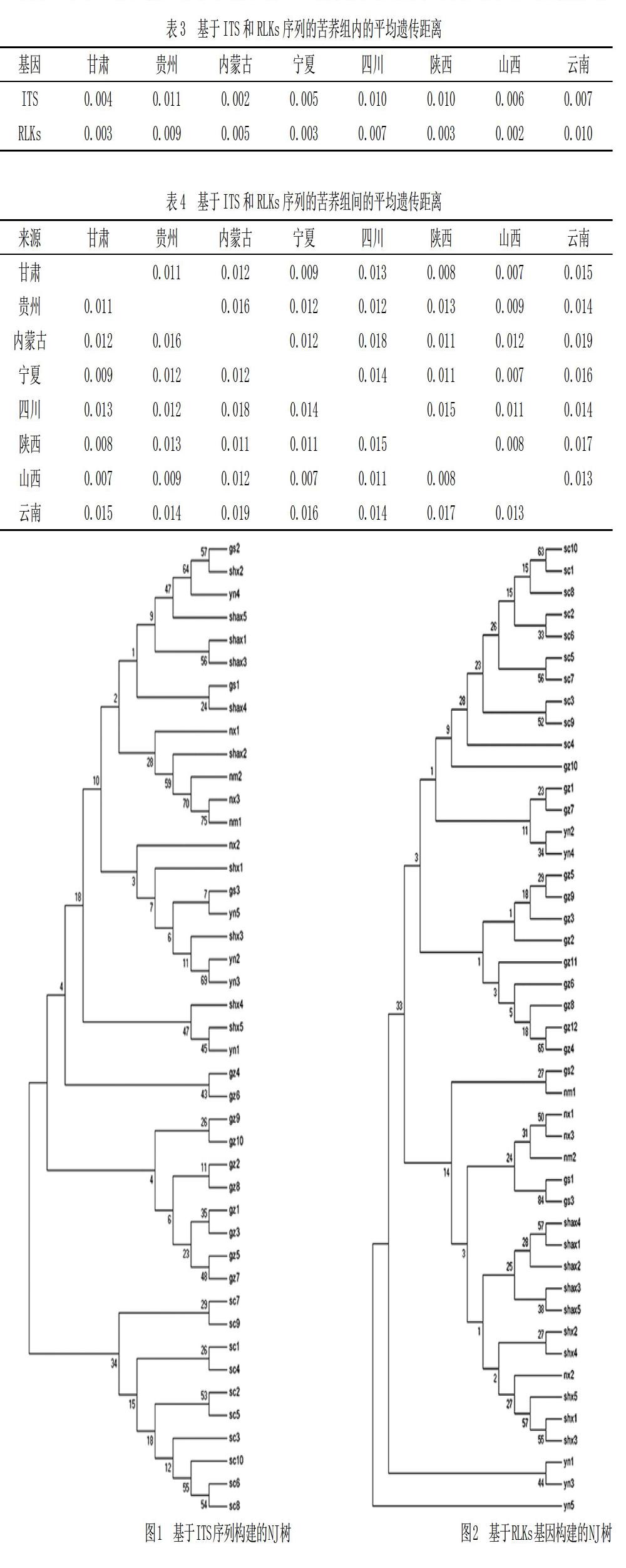
2.3 基于ITS和RLKs序列的系統(tǒng)發(fā)育樹分析
利用ITS和RLKs序列數(shù)據(jù)構(gòu)建的系統(tǒng)發(fā)育樹如圖1、圖2所示。
從圖1和圖2可以看出,基于ITS和RLKs序列構(gòu)建的NJ系統(tǒng)發(fā)育樹中,四川和貴州地區(qū)的苦蕎品種均能夠單獨聚為一類,具有明顯的地域聚類特點。基于ITS序列構(gòu)建的發(fā)育樹中,陜西地區(qū)以北的苦蕎品種未形成明顯的分支,組成較為分散,而云南地區(qū)的苦蕎穿插在不同的分支中。基于RLKs基因構(gòu)建的NJ樹中,同一地區(qū)的苦蕎材料較為明顯的形成地域聚類,但自展值均較低。
3 討論與結(jié)論
3.1 基于ITS和RLKs序列的苦蕎遺傳多樣性
遺傳多樣性是物種多樣性的重要及核心組成部分,代表著生物攜帶的遺傳信息總和,對物種的遺傳、進化和變異具有決定性作用[14]。核苷酸多樣度(π)是個體間待檢測位點的平均多樣性,代表一個基因的遺傳變異程度[15]。本研究中,ITS和RLKs基因序列的核苷酸多樣度(π)分別為0.011 2和0.011 5,均高于苦蕎PAL基因的核苷酸多樣度(π=0.003 4),但兩基因序列的平均雜合度(θ)(ITS/0.024 7,RLKs/0.020 5)均低于苦蕎PAL基因中的平均雜合度(θ=0.014 3)[15]。以上研究結(jié)果表明核苷酸多樣性的變異程度,隨作物基因類型的不同而不同。
本研究中,無論是基于ITS和RLKs序列的數(shù)據(jù)分析結(jié)果均表明,貴州、四川和云南地區(qū)苦蕎材料的遺傳多樣性最豐富,陜西、山西及寧夏地區(qū)次之,甘肅及內(nèi)蒙古地區(qū)的苦蕎材料的遺傳多樣性最低,這與屈洋等[16]研究得出的北方苦蕎產(chǎn)區(qū)的多樣性水平低于西南苦蕎產(chǎn)區(qū)的結(jié)論一致,在一定程度上支持了西南地區(qū)的云南、四川、貴州地區(qū)一帶是苦蕎麥的現(xiàn)代分布中心和起源地之一[17],貴州地區(qū)更接近中國蕎麥次生起源地之一的川中南部地區(qū)[18]等學說。
3.2 基于ITS和RLKs序列的聚類分析
本研究基于ITS和RLKs序列分別對所收集的苦蕎材料進行聚類分析,據(jù)系統(tǒng)進化樹來看,四川和貴州地區(qū)的苦蕎品種均能夠單獨聚為一類,但自展值均較低。形成這一結(jié)果的原因可能是該地區(qū)苦蕎材料在遺傳組成上與其他地區(qū)差異較大,產(chǎn)生了遺傳分化,也有可能與自然環(huán)境、產(chǎn)地的氣候有關,具體原因需要借助進一步的研究來挖掘。
基于ITS序列構(gòu)建的發(fā)育樹中,陜西地區(qū)以北的苦蕎品種未形成明顯的分支,組成較為分散,表明該地區(qū)的苦蕎材料在遺傳組成上差異較小,未形成明顯的遺傳分化。而云南地區(qū)的苦蕎穿插在不同的分支中,說明云南地區(qū)的苦蕎材料既產(chǎn)生了遺傳分化,也具有遺傳相似性。基于RLKs基因構(gòu)建的NJ樹中,同一地區(qū)的苦蕎材料較為明顯地形成地域聚類,但系統(tǒng)進化樹中自展值均較低。
參考文獻:
[1] 陳慶富.蕎麥屬植物科學[M].北京:科學出版社,2012.
[2] 任長忠,趙鋼.中國蕎麥學[M].北京:中國農(nóng)業(yè)出版社,2015.
[3] Ahmed A,Khalid N,Ahmad A,et al.Phytochemicals and biofunctional properties of buckwheat: a review[J].Journal of Agricultural Science,2014,152(3):349-369.
[4] 林汝法.苦蕎舉要[M].北京:中國農(nóng)業(yè)科技出版社,2013.
[5] Jacquemart A L,Ledent J F O,Quinet M,et al.Isbuckwheat (Fagopyrum esculentum Moench) still a valuable crop today?[J].European Journal of Plant Science & Bio-technology,2012,6(2):1-10.
[6] 徐瓏珀.蕎麥營養(yǎng)與化學成份研究進展[J].四川化工,2014,17(4):4-8.
[7] 張玲,高飛虎,高倫江,等.蕎麥營養(yǎng)功能及其利用研究進展[J].南方農(nóng)業(yè),2011,5(6):74-77.
[8] 胡亞妮,張宗文,吳斌,等.基于ITS和ndhF-rpl32序列的蕎麥種間親緣關系分析[J].生物多樣性,2016,24(3):296-303.
[9] 李敏,張宗文,李艷琴,等.基于PAL基因序列的地方苦蕎品種遺傳多樣性分析[J].植物遺傳資源學報,2017,18(3):530-537.
[10] 梁成剛,陳晴晴,石桃雄,等.蕎麥屬植物MAPK基因片段序列比較與進化關系研究[J].浙江農(nóng)業(yè)學報,2016,28(10):1631-1636.
[11] Librado P,Rozas J.DnaSP v5: A software for comprehensive analysis of DNA polymorphism data[J].Bioinformatics,2009,25(11):1451-1452.
[12] Posada D,Crandall K A.Modeltest: Testing the model of DNA substitution [J].Bioinformatics,1998,14(9):817-818.
[13] Tamura K,Peterson D,Peterson N,et al.MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods[J].Molecular Biology & Evolution,2011,28(10):2731-2739.
[14] 蒲艷艷,宮永超,李娜娜,等.中國大豆種質(zhì)資源遺傳多樣性研究進展[J].大豆科學,2018,37(2):315-321.
[15] 陳吉寶,景蕊蓮,員海燕,等.小麥TaDREB1基因的單核苷酸多態(tài)性分析[J].中國農(nóng)業(yè)科學,2005,38(12):2387-2394.
[16] 屈洋,周瑜,王釗,等.苦蕎產(chǎn)區(qū)種質(zhì)資源遺傳多樣性和遺傳結(jié)構(gòu)分析[J].中國農(nóng)業(yè)科學,2016(11):2049-2062.
[17] 楊學樂,何錄秋,邱博,等.蕎麥生物學研究進展[J].作物研究,2018,32(2):169-174.
[18] Tsuji K,Ohnishi O.Phylogenetic relationships among wild and cultivated Tartary buckwheat(Fagopyrum tataricum Gaert.) populations revealed by AFLP analyses[J].Genes & Genetic Systems,2001,76(1):47-52.
(責任編輯:趙中正)
猜你喜歡
農(nóng)產(chǎn)品加工(2023年14期)2023-08-17 10:32:28
農(nóng)業(yè)科技與信息(2021年8期)2021-12-06 04:24:43
詩潮(2021年11期)2021-11-24 19:55:50
四川農(nóng)業(yè)與農(nóng)機(2021年1期)2021-01-05 19:15:18
青年歌聲(2018年5期)2018-10-29 03:18:40
現(xiàn)代食品(2018年21期)2018-02-14 20:51:24
食品安全導刊(2018年30期)2018-01-17 05:47:48
廣東飼料(2016年3期)2016-12-01 03:43:12
家庭百事通·健康一點通(2016年4期)2016-04-22 09:16:57
林業(yè)與生態(tài)(2016年2期)2016-02-27 14:24:01
