LukS-PV通過下調(diào)HDAC3抑制肝癌細胞增殖
2020-07-13 03:51:16汪自然丁篷生馬筱玲
安徽醫(yī)科大學學報 2020年6期
汪自然,馬 凡,丁篷生,馬筱玲
肝癌(hepatocellular carcinoma,HCC)是臨床上尤為常見的消化道惡性腫瘤之一,在全球癌癥相關死亡的主要原因中排名第二位[1]。雖然近年來HCC的診療水平已有了較大程度提升,但臨床療效依舊不理想,晚期HCC患者的5年生存率仍舊處于較低水平。因此,尋找新的靶向治療藥物對于降低HCC的死亡率意義重大。LukS-PV是金黃色葡萄球菌分泌的一種毒素—殺白細胞素(Panton-Valetine leukocidin , PVL)中的S組分。課題組前期研究[2]表明,LukS-PV靶向C5aR誘導白血病細胞株THP-1凋亡。最近研究[3]表明,LukS-PV能夠抑制乳腺癌細胞株MAD-MB-231增殖,促進其凋亡,并誘導其周期阻滯。目前,LukS-PV對HCC細胞的影響及作用機制仍不清楚。組蛋白去乙酰化酶3(histone deacetylase 3, HDAC3)是一種在真核細胞中廣泛表達的Ⅰ類組蛋白去乙酰化酶[4]。HDAC3能夠通過調(diào)控相關基因轉(zhuǎn)錄催化組蛋白的去乙酰化,并參與肝臟的各種生物代謝過程[5]。該研究首先研究LukS-PV對HCC細胞增殖的作用,進一步檢測泛乙酰化水平改變以及使用定量蛋白組學測序,初步探討LukS-PV對HCC細胞增殖的影響及機制。
1 材料與方法
1.1 主要材料人HCC細胞株HepG2和Huh7購于上海中科院細胞庫;DMEM高糖培養(yǎng)基及胎牛血清購于以色列BI公司;青-鏈霉素、胰酶、BCA蛋白濃度測定試劑盒購于上海碧云天公司;CCK-8試劑購于合肥睿捷生物科技公司;Opti-MEM、轉(zhuǎn)染試劑Lipofectamine2000購于美國Invitrogen公司;HDAC3干擾序列由上海吉瑪公司合成;逆轉(zhuǎn)錄酶和SYBR試劑購于日本TaKaRa公司;HDAC3、GAPDH引物由安徽通用生物公司合成;一抗HDAC3、GAPDH抗體購于武漢三鷹生物公司;一抗泛乙酰化抗體購于杭州景杰生物科技有限公司;二抗(羊抗鼠IgG和羊抗兔IgG)購于武漢Abclonal公司;SDS-PAGE配膠試劑盒、快速封閉液、化學發(fā)光液ECL購于上海雅酶生物公司。
1.2 方法
1.2.1細胞培養(yǎng) HCC細胞株HepG2、Huh7在添加了10%胎牛血清、100 U/ml青-鏈霉素的DMEM培養(yǎng)基中培養(yǎng)。
1.2.2LukS-PV的制備 LukS-PV按照常文嬌等[6]提供的方法表達、純化,并進行蛋白定量。
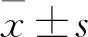
1.2.3CCK-8法檢測細胞增殖能力 取對數(shù)生長期的HepG2、Huh7細胞計數(shù),按4 000個/孔鋪于96孔中,過夜貼壁后加入不同濃度(0.25、0.5、0.75、1 μmol/L)的LukS-PV處理,分別在處理后12、24、36 h每孔加入10 μl的CCK-8試劑,繼續(xù)孵育2 h后使用酶標儀測每孔在450 nm處的吸光度(optical density,OD)。每組分別設置5個復孔,同時設置空白孔。按照細胞存活率=OD(實驗組-空白組)/OD(對照組-空白組)×100%公式計算評估各組細胞增殖能力。
1.2.4定量蛋白組學 定量蛋白組學由杭州景杰生物科技有限公司完成。取對數(shù)生長期的HepG2細胞分為2組進行培養(yǎng),一組加入0.5 μmol/L的LukS-PV,另外一組加入等量的PBS作為對照。培養(yǎng)24 h后離心收集細胞,去除培養(yǎng)液,加入裂解液超聲裂解,4 ℃、12 000 r/min離心15 min留取上清液并使用BCA試劑盒測定蛋白濃度。酶解后的肽段采用TMT標記并使用高ph反向HPLC分級,處理后的肽段再采用液相色譜-串聯(lián)質(zhì)譜分析。二級質(zhì)譜數(shù)據(jù)最后使用 Maxquant (v1.5.2.8)進行檢索并進行生物信息學分析。
1.2.5TCGA數(shù)據(jù)庫分析HDAC3在HCC中的作用 使用UALCAN網(wǎng)站分析TCGA數(shù)據(jù)庫中HDAC3基因在HCC組織及正常肝組織中的表達差異以及高表達和低表達HDAC3患者的預后差異。
1.2.6實時熒光定量PCR(RT-PCR) 提取細胞RNA,并對RNA濃度進行定量,RNA純度OD260/OD280需在1.8~2.0之間方可進行后續(xù)實驗。按逆轉(zhuǎn)錄酶2 μl,RNA 500 ng,總體系10 μl進行逆轉(zhuǎn)錄,反應條件為37 ℃、15 min,85 ℃、 5 s。PCR定量使用羅氏480儀器,PCR體系為TB 10 μl, ddH2O 7.2 μl,上下游引物各0.4 μl,cDNA 2 μl。反應條件為95 ℃預變性30 s;PCR擴增95 ℃、 5 s,60 ℃、 30 s,共計40個循環(huán);融解曲線分析95 ℃、 5 s,60 ℃、 1 min。以GAPDH為內(nèi)參基因,采用2-ΔΔCt法計算HDAC3 mRNA相對表達量。
1.2.7Western blot分析蛋白表達量 預冷的PBS清洗細胞3次,加入蛋白裂解液冰上放置15 min,使用細胞刮將細胞刮至一側(cè)后繼續(xù)置于冰上裂解20 min,期間多次吹打細胞與裂解液使其充分混勻。4 ℃、12 000 r/min離心20 min后留取上清液使用BCA試劑盒進行蛋白定量。獲得的蛋白樣本使用10%的SDS-PAGE分離后,轉(zhuǎn)至NC膜上,轉(zhuǎn)膜條件為200 mA,90 min。使用雅酶快速封閉液室溫封閉30 min后,分別放入一抗HDAC3、GAPDH和泛乙酰化抗體(1 ∶1 000)中4 ℃搖晃孵育過夜。次日,使用TBST洗去未結(jié)合的一抗,加入羊抗鼠或者羊抗兔IgG的二抗(1 ∶5 000)中,室溫孵育2 h,TBST洗去未結(jié)合的二抗。最后使用ECL顯影液于天能曝光儀中顯影。
1.2.8細胞轉(zhuǎn)染 分別取對數(shù)生長期的HepG2和Huh7細胞鋪于6孔板中,待細胞匯合度達到50%~70%時轉(zhuǎn)染。分別取5 μl的SiRNA和5 μl的Lipofectamine2000輕輕加入125 μl的Opti-MEM中,室溫靜置5 min后將二者混合繼續(xù)室溫孵育20 min。期間將各孔培養(yǎng)液更換為無血清DMEM培養(yǎng)基,并將上述混合物加入各組細胞中。培養(yǎng)箱中孵育6 h后將培養(yǎng)液更換為含10%血清的完全培養(yǎng)基后繼續(xù)培養(yǎng)48 h。提取細胞檢測HDAC3 mRNA和蛋白表達量確定敲低效率較高后再進行后續(xù)實驗。
2 結(jié)果
2.1 LukS-PV對HepG2和Huh7細胞增殖的影響采用不同濃度的LukS-PV作用HepG2和Huh7細胞,并使用CCK-8方法于12、24、36 h檢測細胞增殖能力的改變。結(jié)果顯示隨著LukS-PV濃度的增加,細胞抑制率逐漸增高,且細胞抑制率隨著作用時間延續(xù)增加更為明顯,表明LukS-PV可以降低HCC細胞的增殖能力且呈時間依賴性和濃度依賴性。見圖1。
2.2 定量蛋白組學表明LukS-PV作用后HDAC3下調(diào)蛋白翻譯后修飾在腫瘤的發(fā)生發(fā)展中起到了重要的作用。近年來,乙酰化作為一種常見的表觀遺傳學修飾在腫瘤中的作用引起了廣泛的關注。為了探究LukS-PV抑制HCC細胞增殖的機制,首先使用泛乙酰化抗體檢測LukS-PV作用后總體乙酰化水平的改變。結(jié)果顯示,0.5 μmol/L LukS-PV作用24 h后HepG2細胞中蛋白的總體乙酰化水平與PBS組相比增高(圖2B)。進一步使用定量蛋白組學分析LukS-PV作用后蛋白表達譜的改變。通過檢索定量蛋白組學結(jié)果顯示HDAC3表達下調(diào)(圖2A)。為了初步探究HDAC3在HCC中的作用,課題組分析了TCGA數(shù)據(jù)庫中HCC數(shù)據(jù),其中HCC組織371例,正常肝組織50例。圖2C結(jié)果表明,HCC組織中HDAC3 mRNA表達水平(中位數(shù)26.146)明顯高于正常組織中HDAC3 mRNA表達水平(中位數(shù)17.996),差異有統(tǒng)計學意義(P<0.05)。圖2D生存分析結(jié)果進一步表明高表達HDAC3的患者生存期明顯低于低表達患者(P<0.05)。因此,HDAC3可能是HCC中的一個重要的致癌基因。

圖1 不同濃度LukS-PV對HCC細胞增殖的影響

圖2 HDAC3表達下調(diào)及在HCC中的作用
A:蛋白組學熱圖;B:LukS-PV作用后總體乙酰化水平改變;C:HDAC3在HCC組織和正常組織表達差異;D:高表達HDAC3和低表達HDAC3患者預后差異;marker:分子量標準
2.3 RT-PCR和Western blot驗證HDAC3下調(diào)基于定量蛋白組學結(jié)果顯示HDAC3下調(diào),且HDAC3在HCC組織中高表達,與患者預后密切相關,可能是LukS-PV作用的重要靶點。進一步使用RT-PCR和Western blot核實不同濃度LukS-PV作用后HDAC3 mRNA水平和蛋白水平的改變。結(jié)果表明,LukS-PV在HepG2和Huh7細胞株中可以下調(diào)HDAC3的mRNA(F=14.33,P<0.05;F=12.47,P<0.05)和蛋白水平(F=33.66,P<0.05;F=57.97,P<0.05),且呈濃度依賴性。見圖3。
2.4 HDAC3基因敲低驗證為了探討HDAC3在HCC中的作用,實驗使用特異性SiRNA轉(zhuǎn)染HepG2和Huh7細胞干擾HDAC3的表達,并用RT-PCR和Western blot檢測敲低效率。RT-PCR結(jié)果顯示,與NC組相比,干擾組HDAC3 mRNA表達量下降,差異有統(tǒng)計學意義(t=13.23,P<0.05;t=23.21,P<0.05),見圖4A、B。進一步使用Western blot發(fā)現(xiàn)干擾組的HDAC3的蛋白表達水平降低,見圖4C、D。
2.5 HDAC3敲低后對HCC細胞增殖的影響確定HDAC3基因干擾效率較高后,課題組進一步探究HDAC3基因敲低對HCC細胞增殖能力的影響。圖5結(jié)果表明,敲低HDAC3后36 h開始可以降低HCC細胞HepG2和Huh7細胞的增殖能力(t=3.32,P=0.04;t=3.501,P=0.02)。
3 討論
細菌毒素因其與靶細胞結(jié)合的特異性和細胞毒性已成為抗腫瘤藥物開發(fā)的熱點,部分細菌毒素已應用于臨床。例如,從白喉桿菌中提取的白喉毒素,在各種臨床試驗中顯示出抗腫瘤活性,包括腎上腺皮質(zhì)癌、膠質(zhì)母細胞瘤、皮膚T細胞淋巴瘤、乳腺癌和宮頸腺癌[7-8]。課題組前期研究[2-3]表明,LukS-PV對白血病細胞、乳腺癌細胞均展現(xiàn)出良好的抗腫瘤作用。進一步研究[9]發(fā)現(xiàn),LukS-PV靶向C5aR誘導白血病細胞凋亡。而Hu et al[10]報道,C5aR在HCC中高表達,且C5aR刺激后能激活ERK1/2通路,促進HCC的侵襲轉(zhuǎn)移。因此,課題組推測LukS-PV可能對高表達C5aR的HCC細胞也有作用。

圖3 不同濃度LukS-PV下調(diào)HDAC3表達
A、B:RT-PCR檢測不同濃度LukS-PV作用HepG2與Huh7細胞后HDAC3 mRNA表達改變;C、D:Western blot檢測不同濃度LukS-PV作用HepG2與Huh7細胞后HDAC3蛋白表達改變;與PBS組比較:*P<0.05

圖4 HDAC3基因敲低驗證
A、B:RT-PCR檢測HepG2與Huh7細胞敲低HDAC3后HDAC3 mRNA表達量改變;C、D:Western blot檢測HepG2與Huh7細胞敲低HDAC3后HDAC3 蛋白表達量改變;與NC組比較:*P<0.05

圖5 HDAC3敲低后對HCC細胞增殖能力的影響
A:HepG2細胞;B:Huh7細胞;與NC組比較:*P<0.05
近年來,在腫瘤發(fā)生發(fā)展過程中起重要作用的表觀遺傳調(diào)控異常逐漸引起關注。乙酰化,尤其是組蛋白乙酰化,通過調(diào)控染色質(zhì)結(jié)構(gòu)的穩(wěn)定性和相關基因的轉(zhuǎn)錄,進而影響細胞的增殖、凋亡和分化。而組蛋白的乙酰化主要受組蛋白乙酰基轉(zhuǎn)移酶和組蛋白去乙酰化酶調(diào)控。HDAC3作為組蛋白去乙酰化家族(HDACs)中的重要成員,已有多項研究[11-13]表明其參與HCC的發(fā)生發(fā)展。朱柯亭 等[11]報道,HDAC3在HCC組織中表達上調(diào),能夠通過上調(diào)p-STAT3促進HCC的復發(fā)。Wu et al[12]報道HDAC3可以與TRAF6結(jié)合增加致癌基因c-myc的穩(wěn)定性,促進其表達。李瑞寶 等[13]報道,伏立諾他衍生物N25能夠通過下調(diào)HDAC3進而誘導HCC細胞凋亡。因此,HDAC3可能是HCC中的一個重要致癌基因,可能成為未來HCC診療的新靶點。LukS-PV可抑制HDAC3表達,這種廣泛的去乙酰化作用導致細胞增殖作用降低,其中可能的機制較為復雜。Lou et al[14]報道HDAC3可以與人附睪蛋白4結(jié)合并活化PI3K/AKT信號通路促進卵巢癌細胞的增殖。Lu et al[15]研究表明沉默HDAC3后胞質(zhì)中ac-STAT3向p-ATAT3的轉(zhuǎn)化過程被破壞,STAT3信號通路被抑制進而抑制HCC細胞的增殖。
本研究首先研究了LukS-PV對HCC細胞增殖的影響,顯示LukS-PV能夠抑制HepG2和Huh7細胞的增殖。為了進一步探索其機制,課題組采用泛乙酰化檢測和定量蛋白組學研究LukS-PV作用后蛋白水平的變化。結(jié)果表明LukS-PV作用于HepG2細胞后,其蛋白整體乙酰化水平增高。定量蛋白組學研究表明HDAC3下調(diào),與乙酰化水平檢測結(jié)果相一致。通過對TCGA數(shù)據(jù)庫中HCC數(shù)據(jù)的分析,顯示HDAC3在HCC中高表達,且高表達HDAC3的患者預后不良,表明HDAC3可能是HCC發(fā)生發(fā)展中的重要致癌基因,其功能可以進一步研究。課題組使用RT-PCR和Western blot驗證了蛋白組學的結(jié)果,LukS-PV可以下調(diào)HDAC3的表達。進一步使用特異性的SiRNA干擾HDAC3的表達后,HCC細胞HepG2和Huh7的增殖能力降低,表明HDAC3可能在HCC細胞增殖調(diào)控過程中發(fā)揮重要的作用。因此,本研究揭示了LukS-PV可能通過下調(diào)HDAC3的表達抑制HCC細胞的增殖,為HCC的精準靶向治療提供了新的研究思路。當然,LukS-PV調(diào)控HDAC3的具體機制仍需進一步深入研究。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
學苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
