PATZ1通過激活p53依賴性途徑抑制宮頸癌細胞增殖和遷移的機制研究
2020-07-14 10:57:18李秀娟鄭雪絨趙欣媛張露丹張建華李敏敏
山西醫科大學學報 2020年6期
李秀娟,鄭雪絨,趙欣媛,張露丹,張 瑾,張建華,楊 虹,李敏敏,楊 帆
(西安醫學院第二附屬醫院婦產科,西安 710038;*通訊作者,E-mail:medicalzheng01@163.com)
宮頸癌起源于子宮頸上皮,是女性常見的惡性腫瘤,也是世界上最嚴重的惡性腫瘤之一[1]。化療和放療是宮頸癌常見的治療干預手段[2],一般情況下,晚期患者的預后較差,常規治療后的5年生存率約為40%[3]。因此,進一步揭示其分子機制,開發有效的治療策略是非常必要的。
包含POZ/BTB和AT鉤的鋅指蛋白1(PATZ1),也稱為MAZ相關因子(MAZR),鋅指肉瘤基因(ZSG)或鋅指核因子/鋅指蛋白278(ZNF278/Zfp278)是一種轉錄調節因子,已被證明可根據細胞環境來負向或正向調節不同基因的表達[4,5]。幾項研究表明PATZ1在癌癥中的作用[4,5],但其作為癌基因或抑癌基因的癌相關功能仍存在爭議。PATZ1在人惡性腫瘤中的過表達支持了其致癌作用[6,7],而其被siRNAs下調后,則分別阻止或誘導結直腸癌或膠質瘤細胞系的生長或凋亡[8]。此外,研究發現敲除PATZ1的小鼠患上了淋巴瘤和其他腫瘤,這表明PATZ1是淋巴瘤和其他腫瘤中潛在的腫瘤抑制因子[9]。PATZ1在宮頸癌中的表達和作用仍不清楚。因此,本研究分析了PATZ1表達在宮頸癌細胞中的功能并分析宮頸癌細胞增殖和遷移機制。
1 研究對象與方法
1.1 納入與排除標準
納入標準:①確診為宮頸癌鱗狀細胞癌;②診斷前均未接受任何形式的腫瘤特異性治療;③在本院進行了腫瘤組織活檢,能夠獲取可用于研究的組織樣本以及鄰近正常組織(距離腫瘤組織約5 cm)樣本。排除標準:①已經進行了治療;②無法繼續獲得研究所需要的組織樣本。
根據納入與排除標準納入2016-12~2018-12在本院進行了首次手術的患者中,選取20例患者的宮頸癌組織及其鄰近正常組織(距離腫瘤組織約5 cm)。患者平均年齡為(55.16±12.02)歲,范圍為35.2-68.5歲。每位患者均提供書面知情同意書。本研究由西安醫學院第二附屬醫院倫理委員會批準。將樣本冷凍并保存在液氮中備用。患者臨床基線特征見表1。
1.2 細胞培養和轉染
人宮頸癌細胞系(HeLa、SiHa、Caski、ME-180 Ms751和C33A)和正常子宮頸細胞系(Ect1/E6E7)購自中國科學院細胞庫(上海)。HeLa細胞采用DMEM高糖培養基培養,其他細胞均在RPMI-1640培養基中培養,培養基中添加10%胎牛血清。所有細胞培養條件為37 ℃,5% CO2。
通過逆轉錄-聚合酶鏈反應(RT-PCR)分離PATZ1 cDNA,并將其亞克隆到pcDNA載體中以生成pcDNA-PATZ1構建體。然后,通過Lipo3000將pcDNA-PATZ1構建體轉染到HeLa、SiHa及Caski細胞中,構建HeLa/PATZ1、SiHa/PATZ1和Caski/PATZ1作為PATZ1組,同時采用空白載體轉染HeLa、SiHa及Caski細胞構建HeLa/CTRL、SiHa/CTRL和Caski/CTRL作為對照組。進一步孵育48 h后,采用這些細胞進行集落形成、細胞遷移和侵襲及動物致瘤實驗
表1 研究對象臨床基線特征 (例)
Table 1 Baeline of enrolled patients with cervical cancer (cases)

臨床特征納入研究者(n=20)絕經 是8 否12組織學 鱗狀細胞癌6腫瘤大小 <4.0 cm12 >4.0 cm8淋巴結轉移 陰性12 陽性8浸潤深度 <2/312 >2/38FIGO分期 IA2 IB12 II6
。
1.3 RNA提取和實時定量(qRT)-PCR
為了研究PATZ1在宮頸癌腫瘤組織、正常組織及宮頸癌細胞系中表達,從宮頸癌組織、宮頸組織正常樣本和細胞系中提取總RNA,逆轉錄成cDNA。根據標準程序進行逆轉錄,采用Power SYBR Green PCR Master Mix進行qRT-PCR分析。引物序列如下。PATZ1正向:5′-TACATCTGCCAGAGCTGTGG-3′,反向:5′-TGCACCTGC TTGATATGTCC-3′;GAPDH正向:5′-GATCTACCGCATCGACCACT-3′,反向:5′-AGATCCTGTTGGCAAATCTCA-3′。
為了進一步研究PATZ1參與介導細胞遷移抑制的分子機制,分析了HeLa/PATZ1、SiHa/PATZ1和Caski/PATZ1及對照組細胞中在細胞遷移過程的起關鍵作用的一組P53下游基因的表達,包括EpCam,RhoE和Caldesmon。引物序列見表2。
表2 相關基因引物序列
Table 2 Primer sequences of related genes

基因正向引物序列 反向引物序列 EpCam5′-CCATGTGCTG GTGTGTGAA-3′5′-TGTGTTTTAGTTCAATGATGATC CA-3′Caldesmon5′-GAGCGTCGCAGAGAACTTAGA-3′5′-TCCTCTG GTAGGCGATTCTTT-3′RhoE5′-AAAAACTGCGC TGCTCCAT-3′5′-TCAAAACTGGCCGTGTAATTC-3′
1.4 蛋白質提取和蛋白質印跡分析
采用免疫印跡確定PATZ1蛋白在宮頸癌細胞系和正常宮頸細胞中的表達,用含有蛋白酶抑制劑和磷酸酶抑制劑的RIPA緩沖液從培養的正常宮頸細胞(Ect1/E6E7)及6種宮頸癌細胞系(HeLa、SiHa、Caski、ME-180、Ms751和C33A)中提取總蛋白。用BCA試劑盒測量蛋白質濃度。總蛋白在8%-10%聚丙烯酰胺-SDS凝膠電泳中分離,并通過電印跡轉移到硝酸纖維素膜上。將膜用1×TBS,0.1%Tween-20和5%BSA封閉,并與抗GAPDH、抗PATZ1抗體孵育。
1.5 集落形成分析和生長活力測定
為了確定PATZ1的表達是否在宮頸癌細胞的生長中起作用,將pcDNA-PATZ1構建體和空白載體轉染(對照組)的HeLa、SiHa細胞進行了集落形成分析。將細胞以90%的密度接種在培養皿中。培養兩周后,將細胞固定,并用0.1%結晶紫的20%甲醇溶液染色30 min,用PBS洗滌并拍照。為了進一步研究生長抑制的原因,對HeLa/PATZ1、SiHa/PATZ1及各自的對照進行生長曲線和細胞活力測定。將細胞(4×104個細胞/培養皿)鋪在6 cm培養皿中,臺盼藍染色后通過Bürker室計數,以評估細胞活力。
1.6 細胞遷移和侵襲檢測
為了檢測腫瘤細胞遷移能力的變化,進行了細胞劃痕實驗,HeLa/PATZ1作為實驗組,HeLa/CTRL作為對照組。用0.25%胰蛋白酶消化細胞,并調節至5×105細胞/ml細胞懸液的濃度,然后接種到6孔板中并在37 ℃下培養過夜。第2天,將細胞在無血清培養基中培養6 h,達到約95%-100%的融合度,并用20 μl針尖傷細胞單層。然后將細胞用培養基漂洗兩次,并孵育48 h。在0,6,24,30 h,在倒置顯微鏡下拍攝細胞。
為了更好地研究宮頸癌細胞遷移能力,采用Transwell細胞培養室進行測定,HeLa/PATZ1、SiHa/PATZ1和Caski/PATZ1作為實驗組,HeLa/CTRL、SiHa/CTRL和Caski/CTRL作為對照組。用胰蛋白酶/EDTA收集匯合的細胞單層,以1 200 r/min離心5 min,重懸于無血清的培養基中,并鋪板[(3-5)×104個細胞]至8 μm聚碳酸酯膜濾器的上腔室中。下腔室充滿了完全的培養基。然后將細胞在37 ℃,5%CO2中孵育24 h和48 h。擦去過濾器上側的未遷移細胞,并將過濾器反側上的遷移細胞用在20%甲醇中的0.1%結晶紫染色30 min,用PBS洗滌并在光學顯微鏡下拍照。
1.7 動物致瘤實驗
為了研究了Hela/PATZ1,SiHa/PATZ1及其對照組細胞在裸鼠中產生腫瘤的能力。采用6周的雌性裸鼠(購自中科院上海實驗動物中心)皮下注射對照和PATZ1轉染的HeLa和SiHa細胞(2×106個細胞)來評估體內腫瘤生長情況(每組8只)。每3 d測量一次腫瘤體積,計算公式為:體積=長×(寬/2)2。3周后處死所有小鼠進行腫瘤質量測定和分離。小鼠實驗得到倫理委員會的批準。
1.8 統計分析
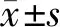
2 結果
2.1 PATZ1在宮頸癌臨床腫瘤組織和細胞系中表達下調
20例宮頸癌標本PATZ1的mRNA表達水平明顯低于相鄰正常組織樣本(見圖1A)。與宮頸正常細胞(Ect1/E6E7)相比,宮頸癌細胞系PATZ1蛋白表達水平明顯降低(見圖1B)。此外,與宮頸正常細胞系(Ect1/E6E7)相比,宮頸癌細胞系HeLa、CaSki、ME-180、Ms751、C33A中PATZ1 mRNA表達水平顯著降低,且差異具有統計學意義(P<0.01),SiHa細胞中PATZ1 mRNA表達水平也顯著降低(P<0.001,見圖1C)。
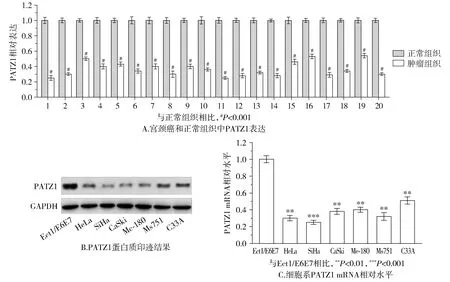
圖1 PATZ1在宮頸癌腫瘤組織和正常組織及宮頸細胞系中表達結果Figure 1 PATZ1 expression in cervical cancer cell lines and tissues
2.2 PATZ1表達抑制HeLa和SiHa細胞集落的生長
與對照組相比,PATZ1組HeLa和SiHa細胞中檢測到集落數明顯降低(見圖2)。與對照組相比,PATZ1組HeLa和SiHa細胞分別顯示出較低的增殖能力,分別在細胞培養7 d或5 d時兩組間差異具有統計學意義(見圖2)。

與CTRL組比較,*P<0.05,**P<0.01,***P<0.001圖2 PATZ1轉染的宮頸癌細胞的細胞集落生長分析結果Figure 2 Cell growth in PATZ1-transfected cervical cancer cells
2.3 PATZ1表達抑制HeLa和SiHa細胞遷移和侵襲
與HeLa/CTRL對比,HeLa/PATZ1的遷移能力顯著降低(見圖3),表明PATZ1可以抑制宮頸癌細胞的遷移。接種后24 h,所有轉染PATZ1的HeLa、SiHa和CaSki細胞侵襲速度都小于對照組細胞(見圖4),結果表明,PATZ1在抑制宮頸癌細胞的遷移和侵襲性中具有關鍵作用。
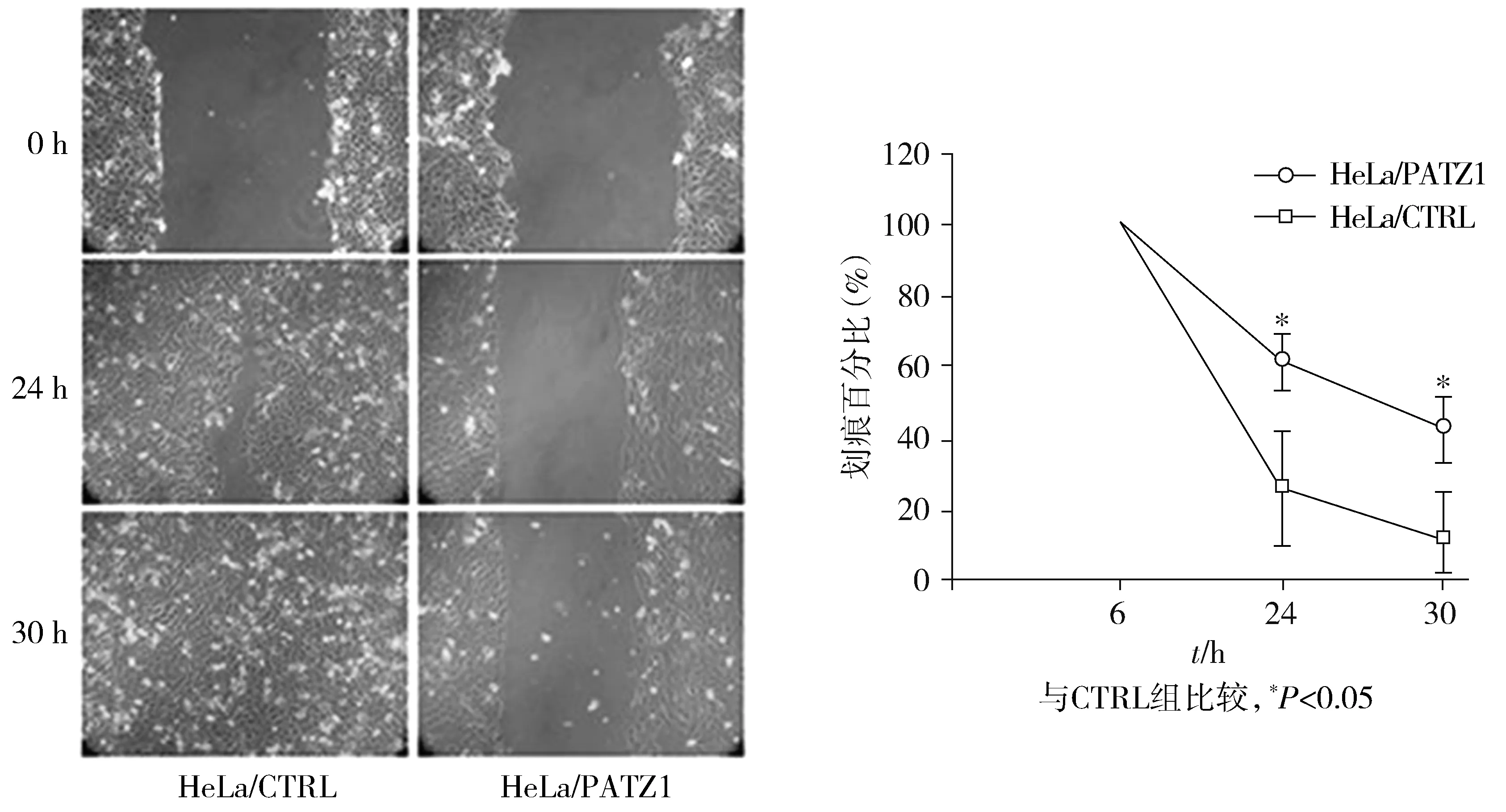
圖3 PATZ1表達抑制HeLa細胞遷移Figure 3 PATZ1 inhibits cell migration of HeLa cells

圖4 PATZ1表達抑制細胞侵襲結果Figure 4 PATZ1 inhibits cell invasion of HeLa,SiHa and CaSki cells
2.4 PATZ1表達抑制宮頸癌細胞的致瘤性
接種空白載體轉染細胞系的小鼠和接種HeLa/PATZ1,SiHa/PATZ1細胞的8只小鼠中均觀察到腫瘤生長。與對照組產生的腫瘤相比,PATZ1組腫瘤大小和腫瘤質量明顯減小(見圖5)。此外與對照組比,HeLa/PATZ1、SiHa/PATZ1組中腫瘤體積較小,分別在第9天和12天兩組間差異具有統計學意義(P<0.05,見圖5)。
2.5 PATZ1表達對p53途徑下游基因EpCam, RhoE和Caldesmon表達的影響
qRT-PCR結果顯示,與CTRL組相比,PATZ1組三種細胞系中EpCam表達下調,RhoE和Caldesmon的表達上調(見圖6),以上結果表明,在HeLa、SiHa和Caski細胞中均存在功能性P53蛋白,至少存在EpCam、Caldesmon和RhoE啟動子且其活性可被PATZ1增強,PATZ1與p53協同作用,抑制宮頸癌細胞的增殖和侵襲。
與各自對應的CTRL組比較,*P<0.05圖5 PATZ1表達抑制小鼠體內腫瘤生長結果Figure 5 PATZ1 expression inhibits tumor growth in mice

與CTRL組比較,*P<0.05,**P<0.01圖6 不同腫瘤細胞系中PATZ1表達對EpCam, RhoE和Caldesmon表達的影響Figure 6 Effect of PATZ1 expression on EpCam, RhoE and Caldesmon expression in different cervical cancer cell lines
3 討論
本研究結果表明,PATZ1在人宮頸癌腫瘤中的表達水平低于鄰近正常組織樣本,宮頸癌細胞中PATZ1表達水平也低于正常宮頸細胞系。轉染PATZ1的宮頸癌細胞中細胞增殖和遷移、集落形成和小鼠腫瘤增殖被抑制。與CTRL組細胞產生的腫瘤相比,PATZ1組腫瘤大小、腫瘤體積和腫瘤質量降低。此外,與CTRL組細胞相比,PATZ1組三種宮頸癌細胞系中EpCam表達下調,RhoE和Caldesmon的表達上調。宮頸癌細胞中PATZ1表達激活了p53依賴性途徑,該途徑抑制宮頸癌細胞增殖和遷移。
已有研究表明,PATZ1和p53之間存在功能性相互作用[10],p53維持轉錄程序通過下調基因(例如EpCam)來阻止上皮-間質轉化(EMT),EMT可促進遷移并侵襲周圍組織,從而引起局部和遠端轉移。而下調的基因可抑制參與穩定細胞-細胞連接的分子(例如E-鈣黏蛋白),或直接抑制黏附機制的成分(如纖連蛋白)的表達[11]。p53調控的基因還包括與抑制足小體形成有關的分子,例如肌動蛋白結合蛋白Caldesmon,它被p53上調[12]。此外,p53還可以上調控制肌動蛋白動力學的分子,例如RhoE,其在抑制伴隨腫瘤細胞遷移和侵襲的細胞骨架變化中發揮作用[11]。因此本研究分析了在細胞遷移過程的起關鍵作用的一組p53下游基因的表達,包括EpCam,RhoE和Caldesmon。結果表明,與對照組比,PATZ1組宮頸癌細胞EpCam表達下調,Caldesmon和RhoE表達均上調,和目前已有研究結論一致。
EMT參與許多生物過程,包括胚胎發育、傷口愈合和癌癥進展[13,14]。越來越多研究表明,EMT在許多類型的癌癥轉移及腫瘤復發中起重要作用[15,16]。因此,鑒定調節這些細胞過程的基因對于靶向癌癥治療具有巨大的潛力。對抗EMT的主要參與者之一是抑癌P53蛋白,其丟失已被證明會影響癌細胞運動性,從而促進癌細胞的侵襲和轉移潛能[17]。
此外,最近研究表明PATZ1能夠與p53相互作用并直接調節p53調控基因的轉錄[18],表明PATZ1可能參與EMT的機制。一致地,本研究發現PATZ1表達導致p53調控基因(EpCam, RhoE和Caldesmon)表達發生變化。后續需要進一步的實驗來更好地闡明PATZ1在不同細胞系中與這些基因結合的動態,以及與功能性P53蛋白存在的關系。
總之,本研究結果表明PATZ1通過調節p53靶基因在宮頸癌中發揮抑癌作用,減少了宮頸癌細胞增殖和遷移以及體內腫瘤的生長。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
學苑創造·A版(2020年9期)2020-10-13 09:41:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
汽車工程學報(2017年2期)2017-07-05 08:13:02
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
