平潭島東北部近岸海域大型污損生物群落結構特征
2020-07-21 05:34:32劉坤林和山李眾何雪寶黃雅琴林俊輝牟劍鋒張舒怡馬麗全俊杰王建軍孫軍
海洋學報 2020年6期
劉坤,林和山,李眾,何雪寶,黃雅琴,林俊輝,牟劍鋒,張舒怡,馬麗,全俊杰,王建軍*,孫軍
( 1. 天津科技大學 生物工程學院,天津 300457;2. 自然資源部第三海洋研究所 海洋生物與生態實驗室,福建 廈門361005;3. 廈門大學 嘉庚學院 信息科學與技術學院,福建 漳州 363105)
1 引言
海洋大型污損生物是指棲息、附著和生長在海中船體和其他人為設施表面的動植物的總稱,主要種類包括藻類、水螅蟲、盤管蟲、牡蠣和藤壺等,海洋生物附著或棲息在海中設施表面并產生不利影響的現象和過程稱為生物污損[1-2]。污損生物對海洋經濟開發的負面影響早已引起人們的廣泛關注,其危害主要包括增加船舶的阻力、堵塞管道、加速金屬腐蝕和對聲學儀器的影響等方面[2-3];從生態研究角度來看,污損生物是海洋生態系統的重要組成部分,其在物質循環、能量流動和水體凈化方面具有重要作用[4]。
我國大型污損生物生態學的研究始于20世紀50年代,主要涉及渤海、黃海、東海和南海等海域的多個海灣和港口[5],其中東海沿岸大型污損生物的研究主要涉及物種多樣性分布、附著機理、污損生態效應及物種入侵等方面的內容[2]。近年來,很多研究通過掛板試驗系統分析了寧德海域[6]、湄洲灣[7]、洋山港[5]、北關港[8]等東海港灣的污損生物物種組成和數量分布,部分研究涉及了污損生物的生活方式、攝食功能群及其二者之間的關系。
平潭島,亦稱海壇島,位于福建省平潭市東部,地處臺灣海峽交通要道,地理位置優越,是福建省第一大島,其周邊海域屬于開敞式海灣,受外海流系影響較大。目前平潭島附近海域大型污損生物的報道僅見于1963-1964年島西南部竹嶼口海域的掛板試驗,分析了該海域污損生物的多樣性和優勢種組成[2],而對于遮蔽程度更低、受外海海流影響更大的東北部海域則缺乏相關研究。另外,隨著經濟的快速發展,人類活動的影響日趨嚴重,了解平潭島周邊海域海洋生物群落結構和多樣性狀況可更好地為合理開發利用海洋資源奠定基礎。
因此,本文擬通過在平潭島東北部海域的周年掛板試驗,對該海域污損生物的群落結構、生活方式、攝食功能群現狀和群落的季節更替等特征進行系統研究,分析其群落組成和優勢種的時空變化規律,以期為東海沿岸海洋設施污損生物的防除和海洋生態系統的保護提供相關理論基礎。
2 材料與方法
2.1 調查區域
于2017年6月至2018年5月在平潭島東北部近岸海域(25.578 8°N, 119.857 7°E)進行污損生物周年掛板試驗,調查位置如圖1所示,水深約為20 m。該試驗站位東臨臺灣海峽,西隔海壇海峽,周邊島嶼眾多,地形復雜;主要的水團活動有浙閩沿岸流、臺灣暖流和附近的上升流,加上臺灣海峽的狹管效應,使得附近海域海流復雜,結構形態多變[9]。
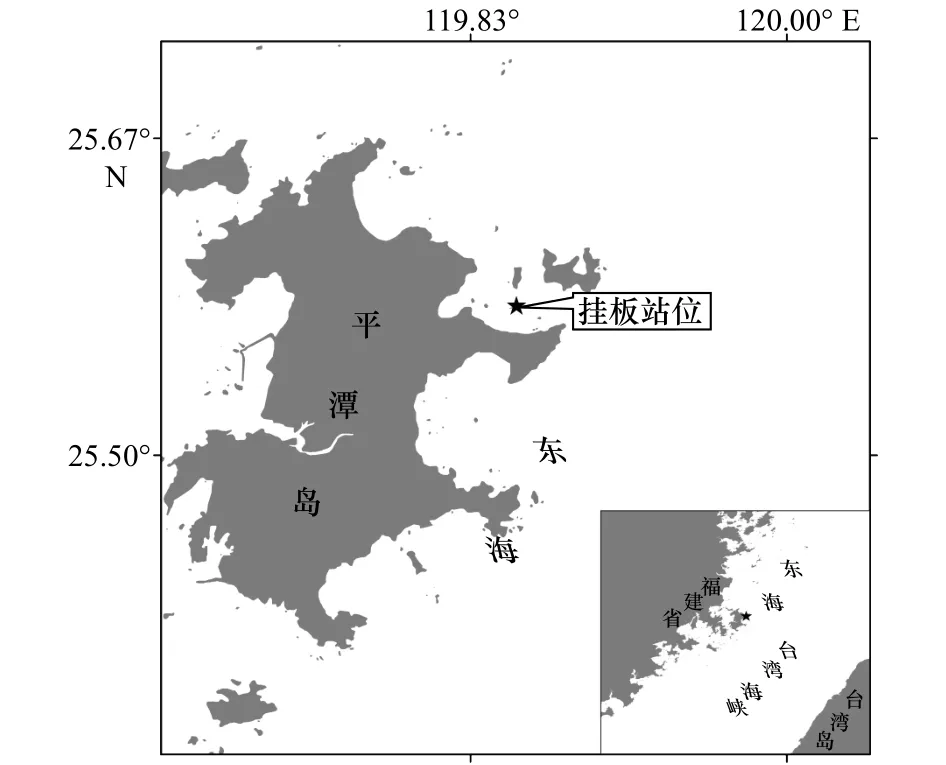
圖1 污損生物掛板試驗站位示意圖Fig.1 The location of macro-fouling investigation
2.2 采樣方法
參照Lin等[6]的方法,采用160 mm(長度)×160 mm(寬度)×20 mm(厚度)的混凝土試板,試板分左右兩面,設A、B兩個平行組掛放。季板、半年板和周年板的收放周期分別按每季、每半年和1周年進行。每組試板均含表層和底層,且試板面與海面垂直;表層試板上沿與水面齊平,底層試板置于水深5 m。全年共回收試板23塊,包括季板13塊,半年板7塊,周年板3塊。樣品處理和分析及資料整理均按《海洋調查規范》(GB/T 12763.6-91)要求進行,其中厚度值為測量試板的4個角落和中央5個點的平均厚度;覆蓋面積率為整個群落覆蓋附著基的百分比率;密度值為逐種記錄單位面積的個體數;生物量為吸去外表水分后的質量。使用YZY4型溫鹽傳感器進行溫度數據的采集,鹽度測量只在2018年3月6日(春季)、2017年6月5日(夏季)、2017年9月2日(秋季)和2017年12月1日(冬季)進行。物種名稱與WoRMS網站(http://www.marinespecies.org/)進行比對并統一。
2.3 數據分析
采用相對重要性指數(Index of Relative Importance,IRI)進行群落優勢種分析[10],IRI≥100 作為優勢種的判別標準,公式如下:
式中,W為物種的生物量占總生物量的百分比,N為物種的豐度占總豐度的百分比,F為該物種的出現頻率。
除大型藻類等初級生產者之外,其他污損生物根據其攝食習性,劃分為5類功能群[11-13]:懸浮物食者、植食者、肉食者、雜食者和碎屑食者。按照生活方式,將污損生物分為固著、半固著和自由活動等3種方式[3]。
利用PRIMER6.0軟件包,基于物種-生物量數據進行Bray-Curtis相似性系數的計算,構建相似性矩陣,用組平均法進行分等級聚合聚類分析和非度量多維標度(Nonmetric Multidimensional Scaling,NMDS),分析之前將生物量數據進行四次方根轉化。采用脅迫系數(Stress)來衡量NMDS分析結果是否具有參考價值:Stress<0.1,可以正確解釋樣本間的相似性關系;0.1<Stress<0.2,具有一定的解釋意義和參考價值;Stress>0.2,不能解釋樣本間的相似性關系[14]。
使用單因素方差分析(One-way ANOVA)進行污損生物物種數、密度和生物量等參數的季節和試板差異。圖表繪制使用ArcMap 10.3軟件。
3 結果與分析
3.1 環境因子
掛板試驗站位的月平均水溫變幅為11.0~27.8℃,9月份(秋季)溫度最高,2月份(冬季)最低(圖 2)。鹽度在4個季節變幅為32.5~34.2,冬季鹽度最低,春季最高,夏、秋兩季鹽度分別為33.23和33.89。
3.2 物種組成
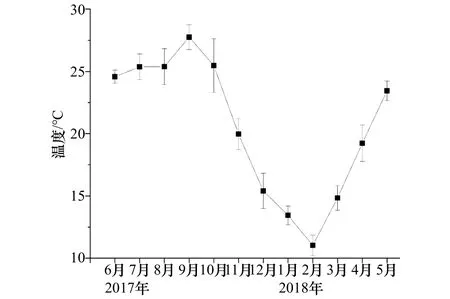
圖2 試驗站位2017年6月至2018年5月溫度變化趨勢Fig.2 The temperature in sampling location during June 2017 to May 2018
23塊水泥試板共鑒定大型污損生物13門92種,其中節肢動物31種、環節動物23種、軟體動物17種,三者構成該海域污損生物的主要優勢類群;另外還有大型藻類5種、刺胞動物5種、扁形動物2種、紐形動物2種、星蟲動物1種、苔蘚動物3種、棘皮動物2種和脊索動物1種(附表A1,附表A2)。
營固著生活的網紋藤壺(Amphibalanus reticulatus)的優勢度最大(IRI=4 116),其次是自由生活的廉形葉鉤蝦(Jassa falcata),二者攝食類型均以懸浮顆粒物為主。其他優勢種分別為固著生活的大室別藻苔蟲(Biflustra grandicella)、三角藤壺(Balanus trigonus)和中胚花筒螅(Ectopleura crocea),自由生活的齒掌細身鉤蝦(Maeropsis serratipalma)、加爾板鉤蝦(Stenothoe gallensis)、細毛背鱗蟲(Lepidonotus tenuisetosus)和四角細身鉤蝦(Quadrimaera quadrimana),以及附著生活的翡翠貽貝(Perna viridis)(表 1)。
3.3 附著量及時空變化
研究海域全年均有生物附著,但附著種類和數量的季節性差異明顯,附著盛期為6月至8月(生物量介于7326.0~12 970.0 g/m2之間),12月至次年 2月為附著淡季(生物量介于39.5~1 580.5 g/m2之間)。表層和底層板附著生物的密度(F=6.281,p<0.05)和生物量(F=4.568,p<0.05)均具有顯著性差異。
表層板中,春季的附著厚度為最高((17±22)mm),秋季最低((4±2)mm);上半年((17±3)mm)和下半年((15±2)mm)的附著厚度差異不大;周年板的平均附著厚度可達(24±10)mm。秋季的平均覆蓋面積為最低(86%±11%),其余季板、半年板和周年板的覆蓋面積均達百分百(表2)。
表層板附著密度不同季節間呈極顯著性差異(F=18.453,p<0.01),其中夏季最高((26 625±7 319)ind./m2),秋季為最低((2 714±2 423)ind./m2);下半年的附著密度((18 863±2 000)ind./m2)明顯高于上半年((5 575±768) ind./m2); 周 年 板 的 附 著 密 度 高 達 (99 625±33 650)ind./m2。在生物附著量(濕質量)方面,夏季最高,可達(11 917.8±1 488.1)g/m2,與其他3個季節有極顯著性差異(p<0.01);下半年((11 049.9±1 413.9)g/m2)也明顯高于上半年((1 061.0±410.9)g/m2);周年板的附著生物量可達(11 005.6±4 113.9)g/m2。節肢動物、藻類、刺胞動物和苔蘚動物為最主要的生物量貢獻類群(表 2)。
底層板中,春、夏、秋、冬季的平均附著厚度差異不大(p>0.05),介于12~15 mm 之間;上半年((10±5)mm)的附著厚度略低于下半年((15±1)mm);周年板的平均附著厚度高達(38±17)mm。除了秋季(74%±37%)和冬季(24%±10%)的試板外,春、夏季板和半年及周年試板的覆蓋面積均達百分百(表3)。
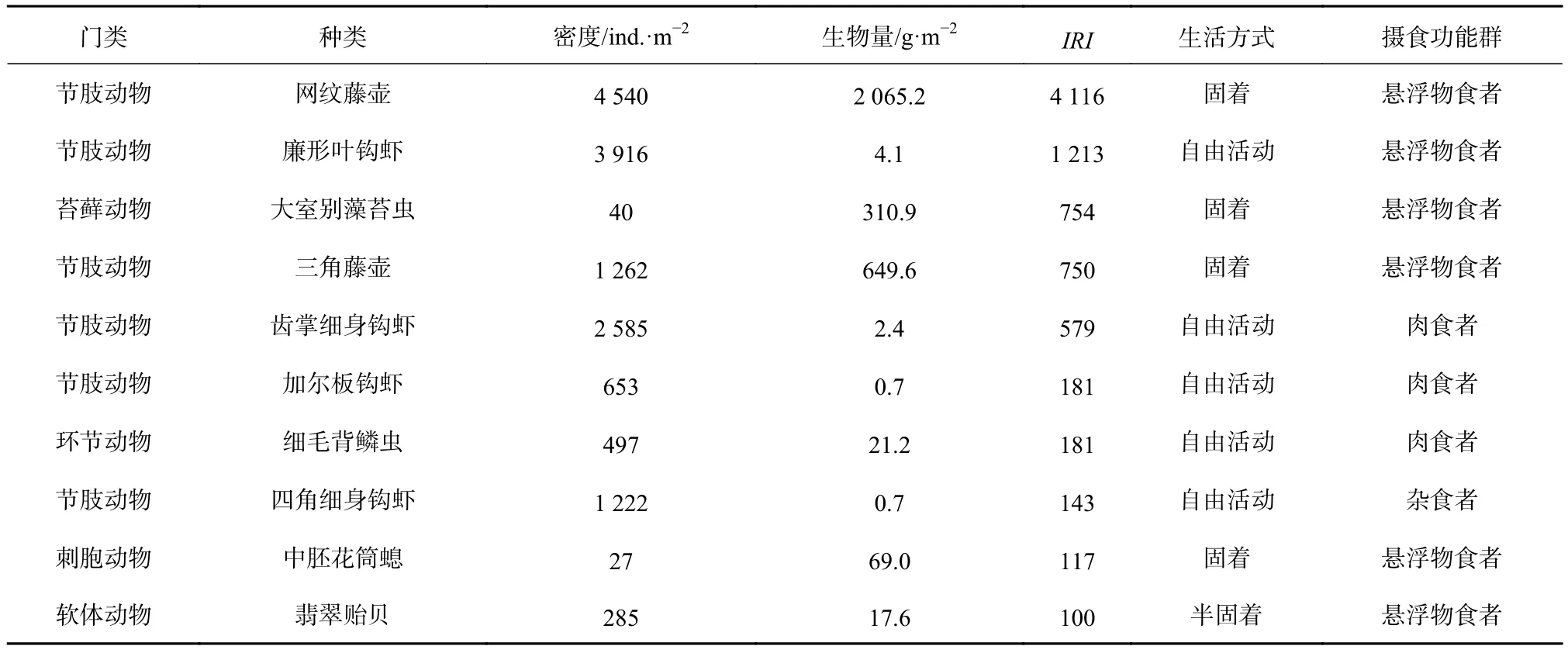
表1 污損生物優勢種Table1 The dominant species of the macro-fouling
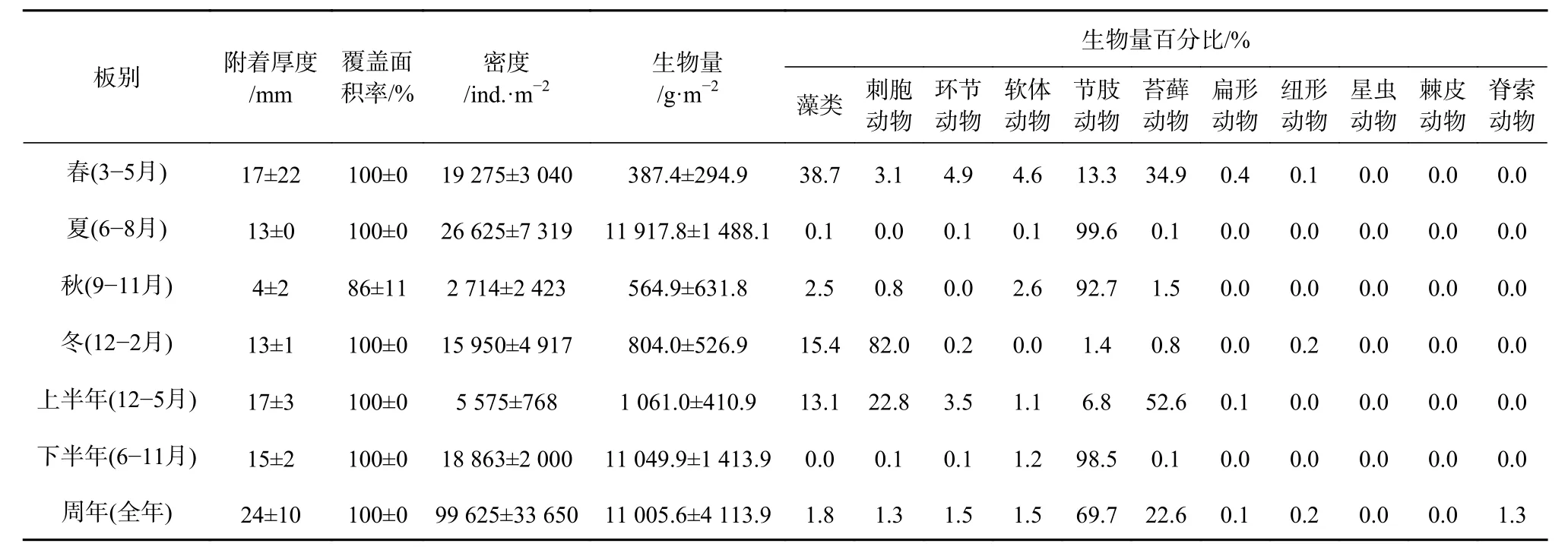
表2 表層板大型污損生物附著狀況Table2 The settlement level of the macro-fouling on the top test panels
底層夏季板的附著密度為最高,可達(35 575±16 228)ind./m2,與其他季節有顯著性差異(p<0.05);下半年的附著密度((16 275±1 520)ind./m2)高于上半年((7 925±1 785)ind./m2);周年板的附著密度為(10 660±1 966)ind./m2。夏季的附著生物量與其他季節有極顯著性差異(p<0.01),可達(9 929.0±3 681.2)g/m2,其中冬季最低,僅為(93.4±36.0) g/m2;下半年的附著生物量((7 584.8±906.2)g/m2)高于上半年((1 099.0±677.4)g/m2);周年板的附著生物量為(4 421.3±505.2)g/m2;節肢動物、刺胞動物和苔蘚動物為最主要的生物量貢獻類群(表 3)。
3.4 群落的季節更替
根據50%的相似度將污損生物群落劃分為5個聚類組(圖3),NMDS顯示Stress值為0.11,分析結果相對可靠;單因素相似性分析表明,5個聚類組間的物種組成呈顯著性差異(Global R=0.989,p=0.001)。聚類組Ⅰ(網紋藤壺群落)出現在表、底層的秋季板上,以固著生活的種類為主,主要出現在溫度較高的夏、秋季(圖4)。聚類組Ⅱ(網紋藤壺-翡翠貽貝群落)出現在表、底層的夏季板和下半年板上,主要附著季節是溫度較高的6-11月,附著強度較大,以固著和半固著生活的種類為主,其中優勢種網紋藤壺附著生物量貢獻率為39.5%,夏季表層板中附著密度和生物量最大可達29 300 ind./m2和12 900.0 g/m2。聚類組Ⅲ(大室別藻苔蟲-細毛背鱗蟲群落)出現在表、底層的春季板和上半年板上,主要附著季節是溫度較低的12月至次年5月,附著強度較小,以固著和活動性生活的種類為主,而且肉食性生物細毛背鱗蟲常棲息于其他固著類生物群落內。聚類組Ⅳ(三角藤壺-大室別藻苔蟲群落)出現在表、底層的年板上,附著強度大,固著、半固著和活動性生活種類均大量存在;優勢種三角藤壺在年板的表層板中附著密度和生物量最大可達10 600 ind./m2和12 720.0 g/m2。聚類組Ⅴ(中胚花筒螅-大室別藻苔蟲群落)出現在表、底層的冬季板上,主要附著季節為溫度較低的冬季,附著強度小,以固著和半固著生活的種類為主。
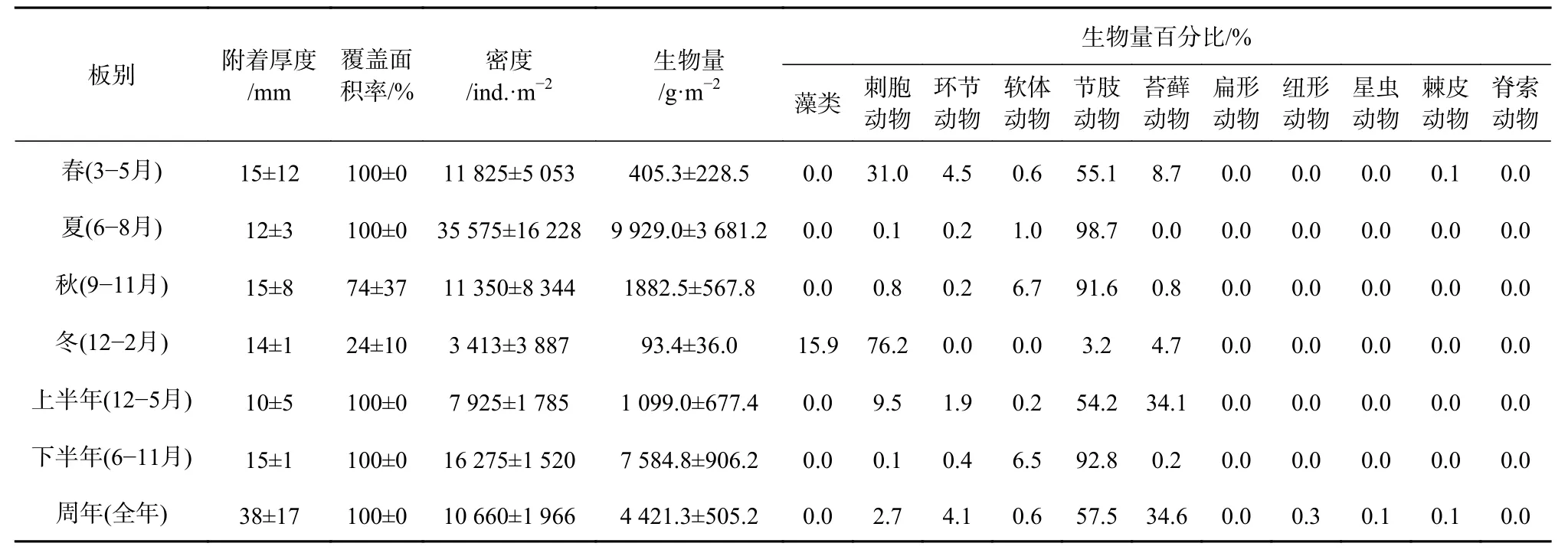
表3 底層板污損生物附著狀況Table3 The settlement level of the macro-fouling on the bottom test panels
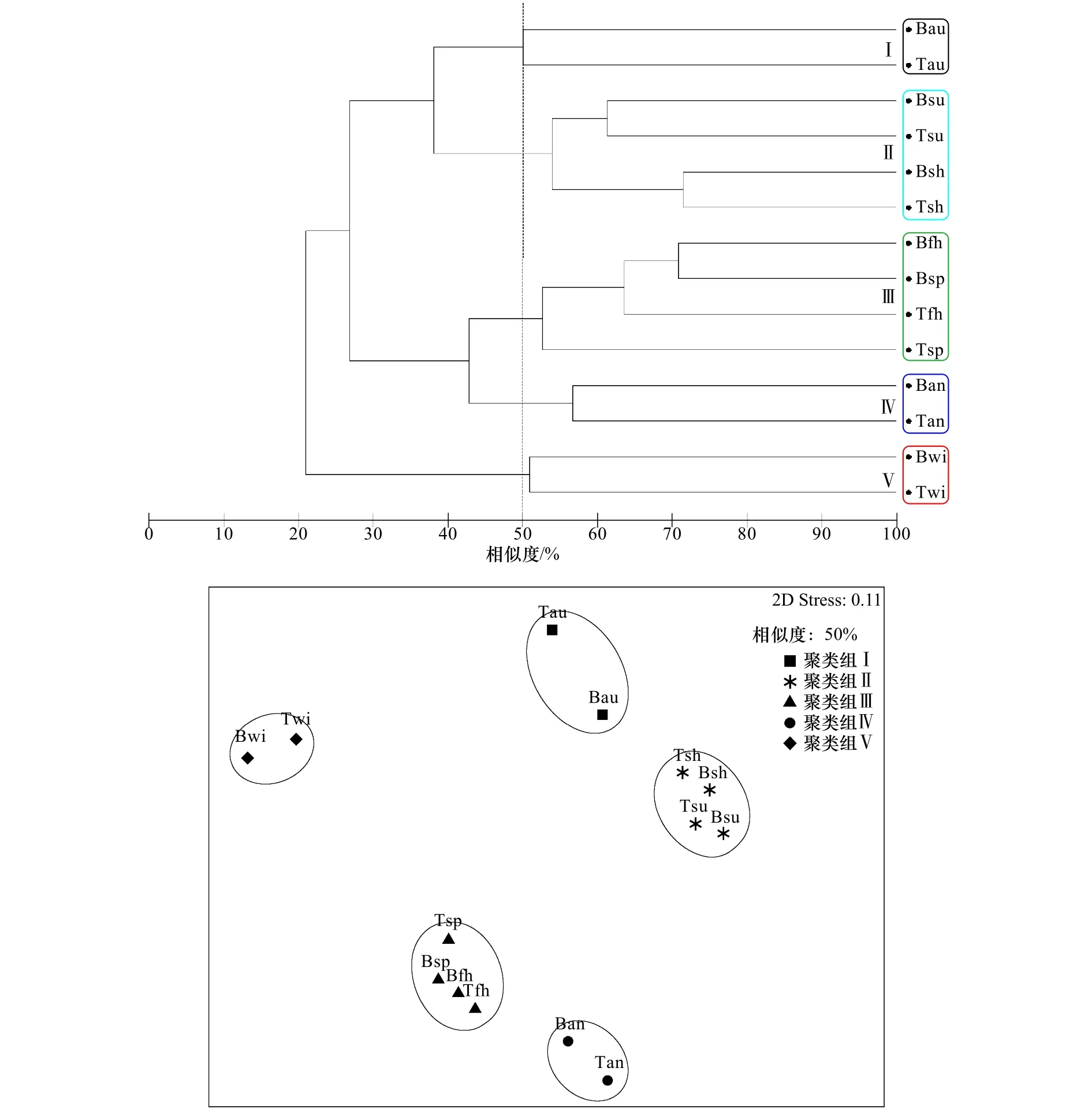
圖3 污損生物群落聚類分析和非度量多維標度排序Fig.3 Cluster and nonmetric multidimensional scaling plots based on biomass of the macro-fouling
3.5 功能群組成
研究海域表、底層試板污損生物均以懸浮物食者為主(圖5),其中春季表層板懸浮物食者占總附著生物量的54.5%,其他季節試板上懸浮物食者的附著量均占總生物量的80%以上。網紋藤壺、三角藤壺、大室別藻苔蟲和中胚花筒螅為懸浮物食者的優勢代表種,其中大室別藻苔蟲和中胚花筒螅的附著高峰期均在12月至次年5月,網紋藤壺則為6-11月,三角藤壺僅在年板上大量附著(圖6)。初級生產者主要出現在春季表層板上,占總附著生物量的38.7%。

圖4 污損生物主要優勢種的附著季節Fig.4 The settlement period and peak time of main fouling organisms
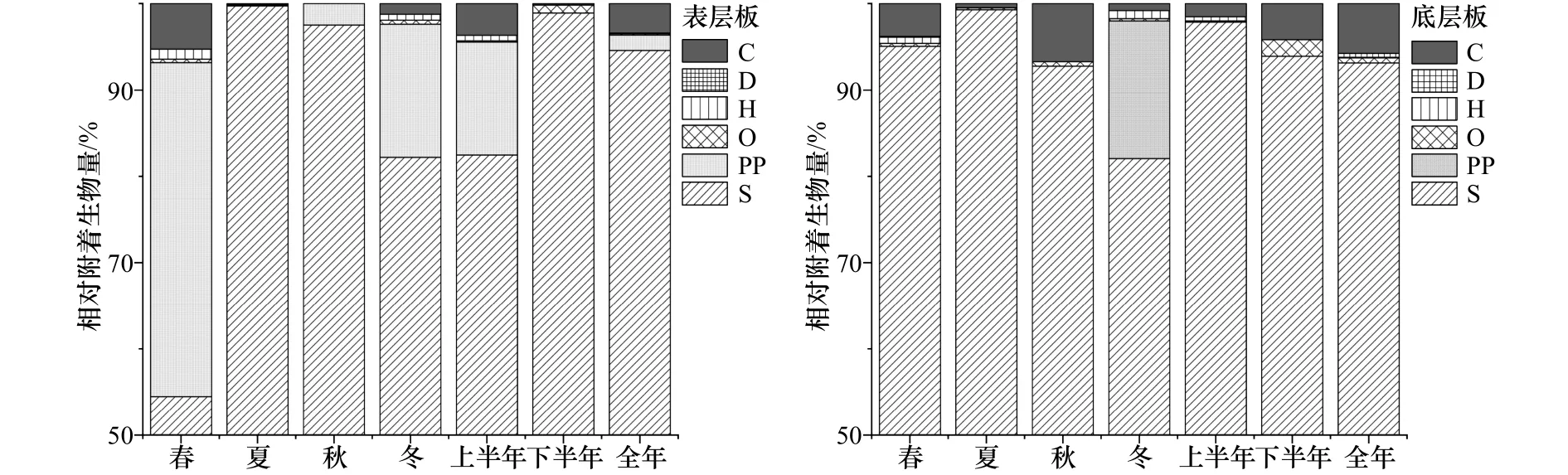
圖5 大型污損生物群落中各攝食功能群所占百分比(相對附著生物量)Fig.5 The relative biomass of the macro-fouling functional feeding groups
污損生物群落按其活動情況和生活方式,可分成固著生活、半固著生活和活動性生活3種類型。在4個季節中表底層板污損生物均以固著型生物為主(圖7),其附著生物量均占總生物量的80%以上。固著生物的食物以懸浮顆粒物為主,優勢種主要為網紋藤壺、三角藤壺、大室別藻苔蟲和中胚花筒螅,其主要分布時期見圖4和圖6。
4 討論
平潭島東北部近岸海域大型污損生物群落種類多且生物量大,以內灣型群落為主,兼具部分外海或高鹽群落特點[2],其中節肢動物和苔蘚動物為最主要的優勢類群。研究海域污損生物種類組成以世界性廣分布暖水種為主,其中固著型生物網紋藤壺是該海域最主要的優勢種,附著季節主要是6-11月,盛期為7-8月,廣泛分布于東海和南海潮間帶至潮下帶區域[15-16];大室別藻苔蟲的分布遍及中國沿海,黃、渤海比南海多[2];鐘巨藤壺廣泛分布于我國長江口外至西沙海域,作為分布于開闊海域的一種巨型藤壺,在內灣很少分布。
黃宗國和蔡如星[2]在平潭島西部海域掛板研究結果顯示,共鑒定污損生物71種,平均生物量為10 785.2 g/m2,優勢種主要為網紋藤壺、總合草苔蟲(Bugula neritina)和管鉤蝦(Corophiidae und.)等,但外海性種類如鐘巨藤壺(Megabalanus tintinnabulum )和紅巨藤壺(Megabalanus rosa)等沒有附著。與本研究結果相比,污損生物物種數較低,生物量較高,主要優勢種則均為網紋藤壺;另外一個明顯的差異是,本研究中出現了鐘巨藤壺等外海性種類。主要原因可能是因為本研究掛板位置位于平潭島東北部海域,離岸相對較遠,鹽度高且較穩定,一般維持在30以上,環境條件有利于部分外海性高鹽種類的生存。
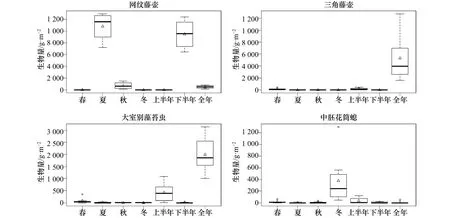
圖6 優勢種附著量狀況Fig.6 The settlement level of dominant species

圖7 不同類型污損生物的相對附著生物量Fig.7 The relative biomass of different lifestyle groups of macro-fouling
黃宗國和蔡如星[2]曾根據夏季底層板的附著量,把中國沿海22個港口的污損生物附著程度分為5個等級,研究海域夏季底層板的附著生物量達(9 929.0±3 681.2) g/m2,屬于第二級,與泉州灣[1](11 295 g/m2)、湄洲灣[7](11 493.21 g/m2)、寧德海域[6](12 353.1 g/m2)、北關港[8](14 313.8 g/m2)和東山灣[17](11 799.0 g/m2)等東海近岸港口污損生物附著狀況相似,遠遠高于受長江沖淡水影響較大的洋山港[5](426.6 g/m2),也明顯大于黃、渤海沿岸的煙臺內港[18]、大連港和青島港[19]等海域,溫度和鹽度應是造成沿海港口污損生物附著量存在較大差異的主要原因。
生物種類溫度屬性的差異是中國沿海各海域之間污損生物差別在“質”方面的表現[2]。東海和南海污損生物種類與黃、渤海,特別是北黃海有很大不同,其中網紋藤壺和三角藤壺廣泛分布于東海和南海的近岸海域,長江口是其在我國近岸海域的地理分布北界,而且越往南其附著期越長,在南海近岸甚至全年均可附著[19]。寒半藤壺(Semibalanus balanoides)、缺刻藤壺(Balanus crenatus)、紫貽貝(Mytilus edulis)和大型褐藻(Laminaria saccharina)等冷水種,呈環寒帶或溫帶分布,其主要分布區域是黃海和渤海沿岸[2-3,20]。本研究調查海域地處亞熱帶,物種種類以亞熱帶暖水種為主,污損性雙殼類中未見紫貽貝、長牡蠣(Crassostrea gigas)等溫水種類出現,取而代之以翡翠貽貝和福建牡蠣(Magallana angulata)等為主。
鹽度是影響污損生物分布的另一關鍵因素[21],不同鹽度區污損生物的種類組成有很大差別。淡水種纖細蓆藻(Phormidium tenue)是河港區域的絕對優勢種;淡水棒螅(Cordylophora lacustris)則是河口區域污損生物的主要優勢種[22]。對于藤壺、泥藤壺(Fistulobalanus kondakovi)適應河口低鹽水域,而網紋藤壺、三角藤壺和鐘巨藤壺等則在東海和南海的高鹽海域出現[2,20]。如果水體鹽度降至18甚至更低,則會妨礙網紋藤壺幼蟲的發育甚至導致其死亡[16]。在本研究中,網紋藤壺、三角藤壺和鐘巨藤壺附著量較大且沒有泥藤壺出現的現象,應與所處海域的鹽度在32以上密切相關。
除此之外,影響污損生物生長的重要因素還應有水體透明度、光照和水深等方面。由于研究海域棲息的附著型藻類多為喜強光藻類(如滸苔和石莼等),水體透明度下降會減弱水下光照和降低初級生產力[23],從而影響藻類的附著[24],因此,本研究中大型藻類一般在表層且光照充足的試板上附著量較大。
因此,大型污損生物群落的形成和發展并非受單一環境因子的影響,而是在溫度、鹽度、水體透明度、水動力條件、地表徑流和養殖等人類活動干擾等因素綜合作用下,最終通過群落結構的演替達到相對穩定的狀態。
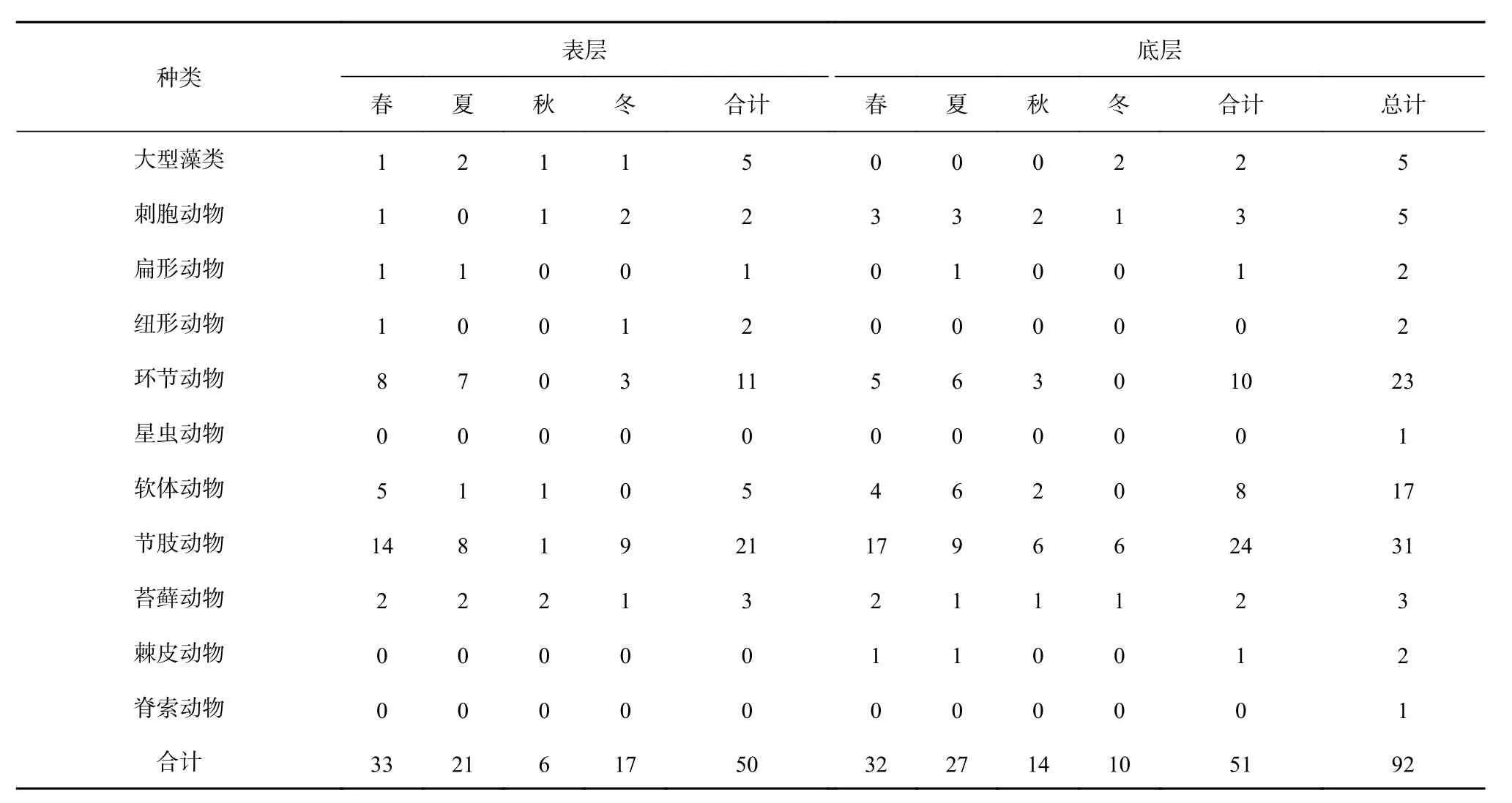
表 A1 大型污損生物種類組成Table A1 The species composition of macro-fouling

表 A2 大型污損生物物種名錄Table A2 The list of macro-fouling organisms

續表 A2

續表 A2

續表 A2
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
