2018-2019年舟山近海浮游動物群落結構春季年際變化及其與水團的關系
2020-07-21 05:34:36王慧君許永久俞存根章寒張洪亮蔣日進梁君郭愛朱文斌徐開達劉連為王皓學
海洋學報 2020年6期
關鍵詞:差異
王慧君,許永久*,俞存根,章寒,張洪亮,蔣日進,梁君,郭愛,朱文斌,徐開達,劉連為,王皓學
( 1. 浙江海洋大學 水產學院, 浙江 舟山 316022;2. 浙江省海洋水產研究所, 浙江 舟山 316021)
1 引言
舟山近海海域位于長江口以南,杭州灣以東,受沖淡水(長江、錢塘江兩大江河)以及來自外海的臺灣暖流的強烈影響,水域環境復雜多變。水團交匯帶來大量營養鹽,為該海域群落結構產生顯著影響[1-8]。尤其是,浮游動物具有隨波逐流的生活方式,使之對海洋中水團的變化非常敏感[9]。因此,研究浮游動物群落結構與水團的關系具有重要的意義,不僅對分析浮游動物群落結構、數量分布以及其對海洋環境變化的響應具有科學意義,也可為海洋生態系統結構和功能的修復提供重要的參考。
目前,關于舟山近海的浮游動物群落結構與時空分布有較多的研究報道[6-10]。這些研究大多集中在浮游動物種類組成、數量空間分布、多樣性特征及其影響因素,而對于舟山近海水團動態變化對浮游動物群落結構的影響方面的研究較少。本文根據2018-2019年春季在舟山近海調查所獲得的浮游動物和水團數據,研究了該海域浮游動物的種類組成、數量分布以及與水團的關系,探討浮游動物群落結構年際變化的主要原因,以期為舟山近海生態環境評價以及生態系統修復提供基礎資料和科學依據。
2 材料與方法
2018年4月8-22日和2019年4月11-23日(春季),搭乘“浙普漁43019”進行兩個航次的浮游動物生態調查,調查區域為 29°~32°N,121°~125°E,調查站位分別為41個和37個,近岸平均水深約8~10 m,近海約15~60 m(圖1)。浮游動物樣品采集均采用淺水I型浮游生物網(口徑50 cm、篩絹CQ14,網長145 cm、孔徑0.505 mm)進行,根據網口流量計計算濾水體積,所獲樣品經5%福爾馬林溶液固定帶回實驗室后,去除樣品中的雜物,通過顯微鏡鏡檢進行分類鑒定和計數。將每個站位的浮游動物統一換算為單位水體中的豐度(ind./m3)。使用溫鹽深儀(CTD)同步觀測水溫、鹽度和葉綠素a濃度。浮游動物樣品的采集、保存、運輸和分析均按照《海洋調查規范:海洋生物調查》(GB/T 12763.6-2007)中的規定執行。
2.1 數據處理
2.1.1 浮游動物類群劃分
本文采取張冬融[11]對浮游動物分類的方法,將鑒定的浮游動物主要劃分為橈足類、毛顎類、水母類、糠蝦類、磷蝦類、十足類、浮游幼體、夜光蟲與其他類共9個浮游動物類群(表1)。夜光蟲雖為浮游植物甲藻門,但它能夠攝食浮游生物、硅藻甚至魚卵。最近10多年,夜光蟲在東海暴發頻繁,對浮游生物生態系統影響巨大[12-14],本文考慮將其作為一種特殊的浮游生物,可以與浮游動物對等,在分類中將它單獨作為一類進行研究。所鑒定物種如端足類、多毛類、介形類、海樽類等在所有站位中出現頻率低于5%,本文將其統歸為一類,即其他類進行研究。為降低浮游動物數據的極化程度,減少稀有種的影響,對浮游動物豐度采用對數轉化,,N為原始豐度,Yi為轉換后數值。
2.1.2 浮游動物豐度和優勢度
浮游動物豐度(N)計算公式為
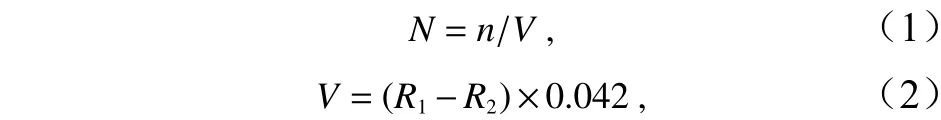
式中,N為單位體積海水中浮游動物的個體數(ind./m3);V為采集浮游動物樣品過濾的海水體積(m3);n為樣品鑒定計數獲得的浮游動物個體數;R1為浮游動物采樣網出水時的流量計讀數;R2為浮游動物采樣網下水前流量計讀數。
浮游動物優勢度(Y)計算公式為
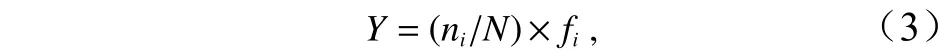
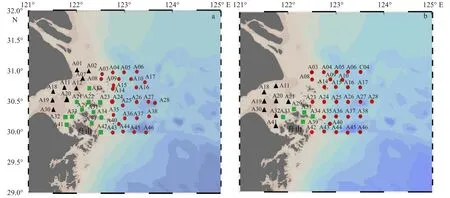
圖1 2018年(a)和 2019年(b)春季調查站位Fig.1 Sampling stations of spring 2018 (a) and spring 2019 (b)

表1 舟山近海海域浮游動物種類組成Table1 Composition of zooplankton in the Zhoushan nearshore waters
式中,N為各采樣站位所有種類的豐度;ni為第i種的豐度;fi為該種在各站位出現的頻率。根據以往學者的觀點,本文將Y大于0.02的種類定為優勢種。環境因子空間分布圖、浮游動物類群豐度分布圖等均使用海洋數據視圖軟件(Ocean Date View)繪制。
2.2 統計分析
根據每個站位的表層溫度(Sea Surface Temperature,SST)和表層鹽度(Sea Surface Salinity,SSS),利用SPSS19.0軟件中的Ward法進行空間站位聚類,據此,進行水團的劃分。根據舟山近海水文條件,將該海域劃分為杭州灣水團(I區)、舟山本島上升流水團(II區)、舟山近海水團(III區);利用 Primer5.0 軟件中的相似性百分比分析(SIMPER)分析不同浮游動物類群、物種在群落結構中的貢獻率。采用單因子相似性分析(ANOSIM)對不同水團間的群落結構的差異顯著性進行檢驗。應用典范對應分析(Canonical Correspondence Analysis, CCA)來研究類群組成及與環境因子關系。使用CCA三變量圖,研究物種組成、站位及環境因子之間的關系。顯著性檢驗根據Monte Carlo 檢驗確定。變量的重要性由變量對應軸之間的相關系數R決定,當|R|≥0.4時,該變量即認為具有重要意義[15]。
3 結果
3.1 水文條件及水團劃分
2018-2019年春季 SST、SSS、表層葉綠素(Chlorophyll a,Chl a)濃度都呈現出近海高,近岸低的趨勢。2018年春季SST整體較2019年低,2018年舟山近海水團SST為14~16℃,上升流水團 SST為12~13.5℃;2019年舟山近海水團SST為15.3~18.3℃,上升流水團SST為13.4~13.8℃;2018-2019年春季舟山近海水團SSS較高(26~35),杭州灣水團SSS低(<15);2018-2019年春季舟山近海Chl a濃度差異較明顯,2018年 Chl a濃度較低(1.5~2.5 mg/m3),而2019 年出現 Chl a 濃度高值(2.5~5 mg/m3)(圖 2)。
根據對SST、SSS的聚類分析結果,可以將調查區域分為3個水團:杭州灣內水團、舟山本島上升流水團、舟山近海水團(圖3)。杭州灣水團為低鹽水,舟山本島上升流水團為低溫水,舟山近海水團為高溫高鹽水。
3.2 舟山近海浮游動物群落結構
2018年和2019年分別鑒定浮游動物50種和31種(表1),2018年鑒定物種如中華哲水蚤、五角水母,拿卡箭蟲、太平洋磷蝦以及橈足類無節幼體,它們分別屬于橈足類、水母類、毛顎類、磷蝦類、浮游幼體。水母類為2018年春季的絕對優勢類群,共鑒定出16種,占浮游動物總種數的32%,橈足類為第2大優勢類群,共鑒定出13種,種數比例為26%。另外其他類 10種(20%),浮游幼體 6種(12%),磷蝦類2種(4%),毛顎類、糠蝦類、十足類各1種(2%)。
與2018年相比,2019年所鑒定出的種數明顯減少,橈足類成為絕對優勢類群,共鑒定出10種,種數比例為32.26%。水母類僅有4種,種數比例為12.90%;另外6大類群鑒定種數由多到少依次為:其他類6種(19.35%),浮游幼體和十足類各 3種(9.68%),糠蝦類2種(6.45%),毛顎類和磷蝦類各1種(3.23%)。夜光蟲在2019年數量巨大,在近海區域分布廣泛(3.23%)。
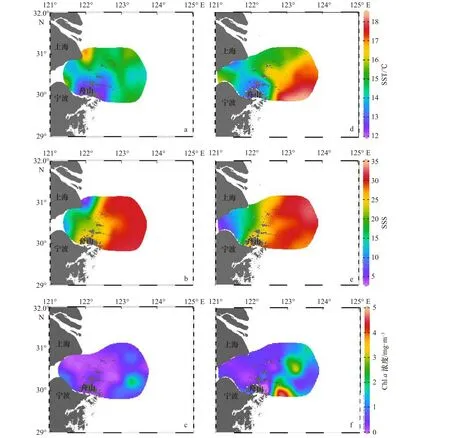
圖2 2018年春季與2019年春季舟山近海海域表層溫度(SST)、鹽度(SSS)、葉綠素a(Chl a)濃度分布與年際變化Fig.2 Surface temperature, salinity, Chl a concentration distribution in the Zhoushan nearshore waters in spring 2018 and spring 2019

圖3 2018 年春季(a)與 2019年春季(b)水團聚類Fig.3 Water mass group clustering of sampling stations in spring 2018 (a) and spring 2019 (b)
水團對春季浮游動物群落結構空間分布格局具有顯著影響(Anosim 檢驗,p<0.05),2018 年春季,Ⅰ區-Ⅱ區、Ⅱ區-Ⅲ區平均相異性分別為48.67%、63.83%,2018年春季所有物種的SIMPER分析表明,引起3個水團區差異的主要貢獻種(貢獻率>10%)為中華哲水蚤。2019年春季,Ⅰ區-Ⅱ區、Ⅱ區-Ⅲ區平均相異性分別為44.76%、64.91%,引起Ⅰ區與Ⅱ區、Ⅱ區與Ⅲ區差異的主要貢獻種為中華哲水蚤(貢獻率分別為52.98%、63.10%)。Ⅰ區與Ⅲ區差異的主要貢獻種為捷氏歪水蚤(Tortanus derjugini)(貢獻率 38.85%)(表2)。水團對浮游動物群落結構年際差異亦具有顯著影響(Anosim 檢驗,p<0.05),Ⅰ區、Ⅱ區、Ⅲ區年際平均相異性分別為:52.21%、50.72%、75.58%。引起2018年和2019年Ⅰ區、Ⅱ區、Ⅲ區以及整個調查區域年際差異的主要貢獻種為中華哲水蚤、夜光蟲和捷氏歪水蚤(貢獻率分別為33.63%、19.15%、12.75%)(表 3)。
3.3 舟山近海優勢種及其空間分布格局
2018年春季優勢種有中華哲水蚤、拿卡箭蟲、小擬哲水蚤、擬長腹劍水蚤(表4)。其中,中華哲水蚤在3個水團區出現頻率均較高(圖4a),在近海海域出現高值,平均豐度達124.93 ind./m3;拿卡箭蟲多分布在舟山近海海域(圖4b),平均豐度13.25 ind./m3;小擬哲水蚤和擬長腹劍水蚤分布區域相似,更多的分布在近海海域(圖4c,圖4d),平均豐度分別為6.58 ind./m3、6.12 ind./m3。與2018年相比,2019年春季水溫的升高,長江徑流量增大,調查海域優勢種發生改變,分別為中華哲水蚤、捷氏歪水蚤、五角水母、夜光蟲。中華哲水蚤僅在上升流水團和近海水團出現頻率較高(圖4e),平均豐度12.28 ind./m3;其次捷氏歪水蚤在杭州灣內較集中(圖 4f),平均豐度 8.17 ind./m3;五角水母在上升流水團數量較大(圖4g),平均豐度2.46 ind./m3。
2018年總豐度時空分布特征為近海(Ⅲ區)高,近岸(Ⅰ區和Ⅱ區)低(圖4i),而2019年分布特征為近海(Ⅲ區)低,近岸(Ⅰ區和Ⅱ區)高(圖4j)。春季浮游動物總豐度年間差異顯著(Anosim 檢驗,p<0.05),兩年浮游動物總豐度分別為:7 152.78 ind./m3,1 007.43 ind./m3。
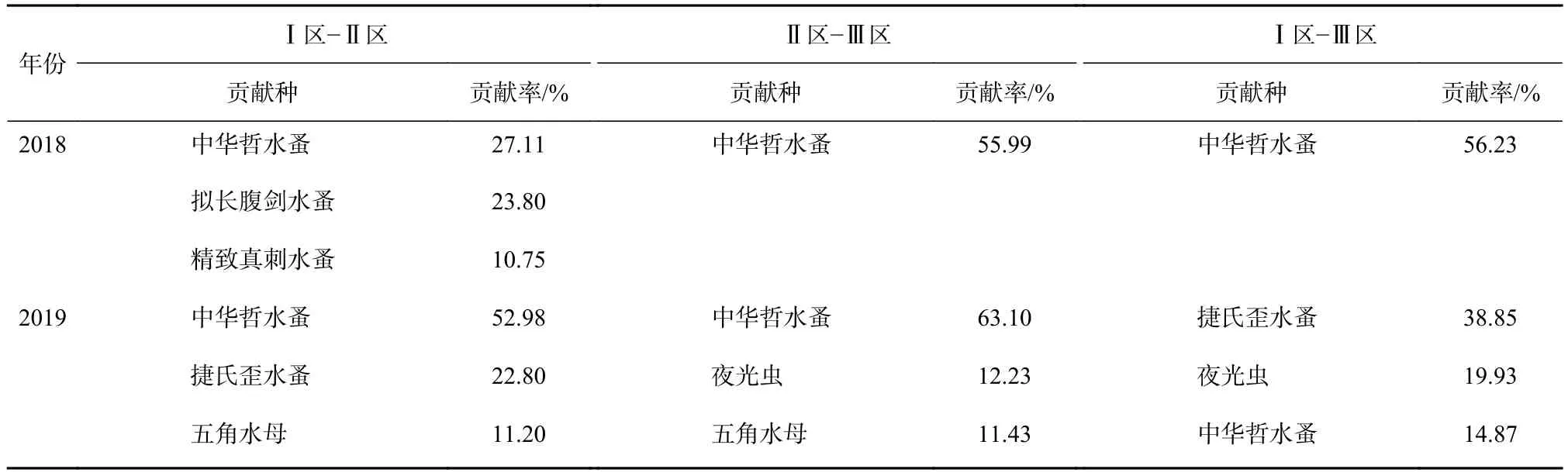
表2 2018年和2019年春季各水團間差異的主要貢獻種和貢獻率Table2 Main contribution species and contribution rate of differences among water masses in spring 2018 and spring 2019, respectively
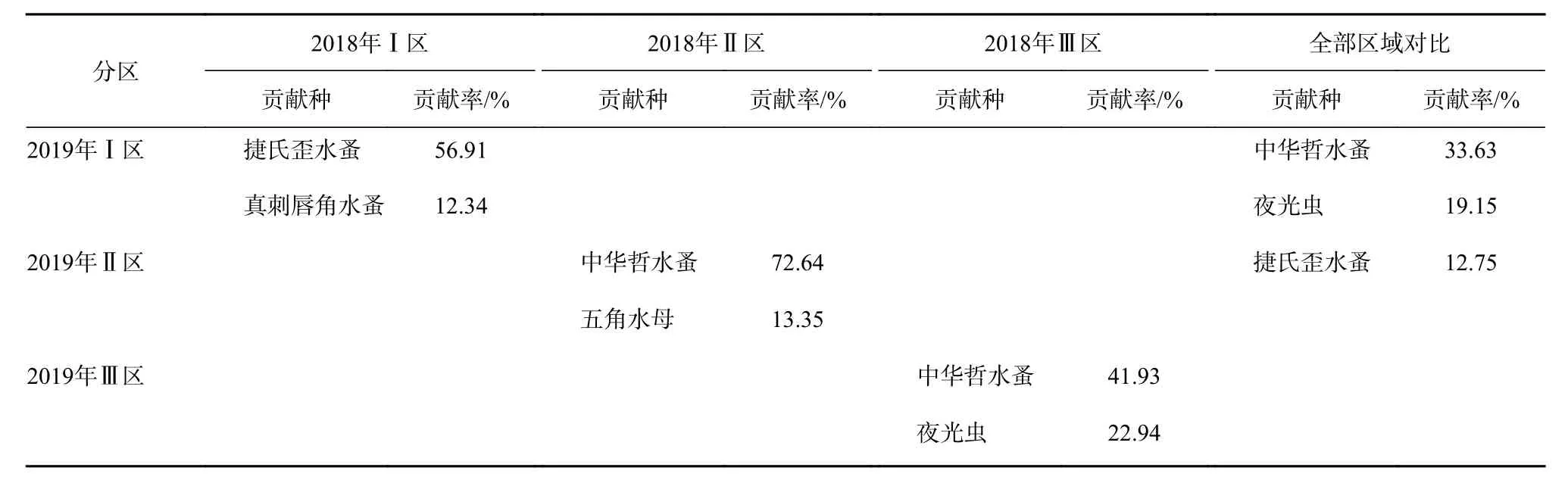
表3 2018年和2019年春季不同水團對浮游動物群落結構組成年際變化和區系格局的影響Table3 The influence of different water masses on the interannual variation and floristic pattern of zooplankton community in spring 2018 and spring 2019
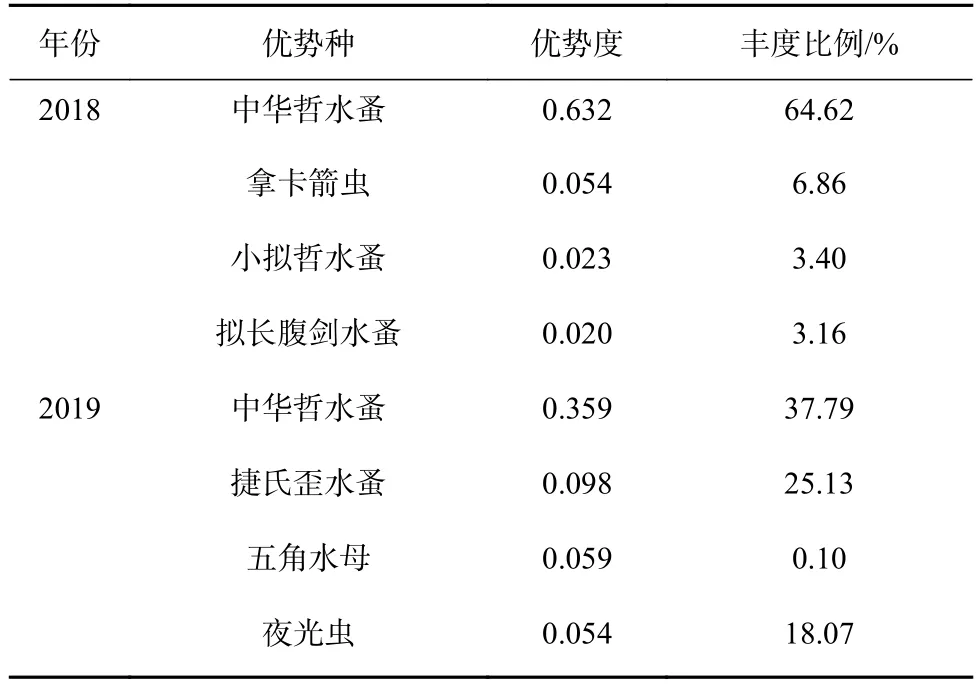
表4 2018年和2019年春季優勢種和優勢度Table4 Dominant species and their dominance in spring 2018 and spring 2019
3.4 舟山近海浮游動物春季優勢種數量年際變化與水團及環境因子的關系
2018年春季與2019年春季各優勢種豐度和環境變量的CCA分析結果顯示(圖5),前4軸與環境變量之間的相關性顯著(F=10.7,p<0.01),第1CCA軸(CCA1,特征值:0.44)和第2CCA 軸(CCA2,特征值:0.37)表現出很高的物種-環境相關性,CCA1和CCA2解釋了物種累計方差百分比的46.14%和物種-環境累計方差總百分比的97.82%,Chl a濃度、海底層溫度(SBT)、SST、海底層鹽度(SBS)和 SSS聚集在CCA1的負軸,與CCA1呈負相關。CCA1主要反映了近海水團和灣內水團的差異。CCA2正負軸兩邊呈現較好的2018年站位和2019年站位的年際差異。捷氏歪水蚤聚集在杭州灣內水團(CCA1的正軸),與SSS、SBS、SST及SBT呈現明顯的負相關性,說明SSS、SST是影響其分布水域的主要因素。同時,捷氏歪水蚤在2019年呈現高值,說明沖淡水勢力較強,灣內SSS較低,導致該河口種類在2019年春季聚集。同時,優勢種夜光蟲聚集在CCA2的正軸,說明2019年春季的高SST、高Chl a濃度是影響夜光蟲分布的主要因素。而中華哲水蚤、擬長腹劍水蚤、小擬哲水蚤、拿卡箭蟲和五角水母則聚集在近海海域,說明暖溫環境是影響其分布的主要因素。這些分隔模式可能反映了浮游動物對棲息地類型的偏好或者其生態習性的適應性。

圖4 2018年和2019年春季浮游動物優勢種分布Fig.4 Distribution of dominant zooplankton species in spring 2018 and spring 2019
4 討論
4.1 春季浮游動物優勢種的年際變化與水團的關系
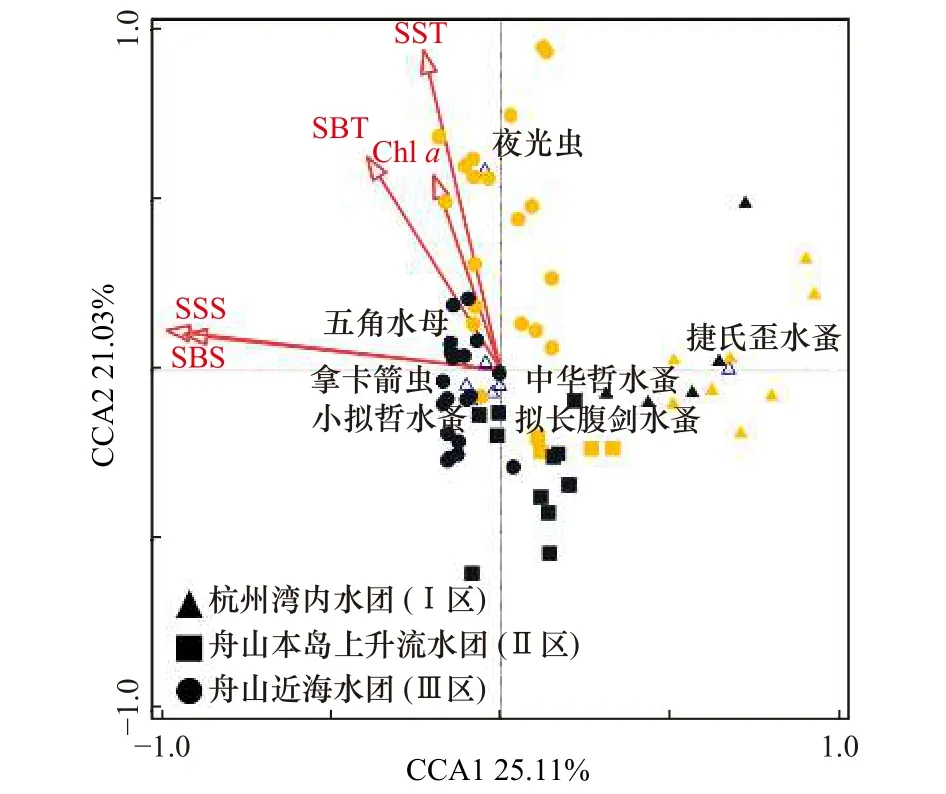
圖5 浮游動物優勢種與環境因子典范對應分析Fig.5 Correspondence analysis between dominant zooplankton species and environmental factors
本研究結果顯示,2018年各水團區域浮游動物豐度由高到低依次為舟山近海水團(平均332.37 ind./m3)、上升流水團(平均11.11 ind./m3)、杭州灣水團(平均10.17 ind./m3)。無論是物種組成還是類群組成,3個劃分區域所有浮游動物組成均具有顯著差異(表2),此外,在近海海域,近海種如中華哲水蚤、拿卡箭蟲豐度最高,顯示出近海水團具有適合近海種棲息的環境。這與以往研究[15-18]相一致。春季水溫的回升,有利于浮游植物的生長繁殖,不僅為浮游動物提供了豐富的餌料,而且為浮游動物攝食浮游植物提供了較好的環境條件[19-21]。
然而,與2018年春季的分布趨勢相反,2019年春季各水團區域浮游動物豐度由高到低依次為:上升流水團(平均 57.52 ind./m3)、杭州灣水團(平均 44.75 ind./m3)、舟山近海水團(平均22.07 ind./m3),相關分析也表明,2019年春季研究海域浮游動物豐度與水溫和鹽度呈顯著負相關(p<0.05)。造成這一差異的主要原因可能為2019年春季近海夜光蟲暴發,大量聚集在近海高Chl a濃度區域,侵占浮游動物原本的生存空間[12-13],導致其他浮游動物如中華哲水蚤、捷氏歪水蚤在其他海域(杭州灣水團、上升流水團)聚集,最終影響浮游動物的分布格局或生物量,這一結果與胡翠林等[14]的研究類似。
2018年春季長江沖淡水徑流量(19 823.64 m3/s),顯著小于2019年(26 387.1 m3/s),本文猜測沖淡水的差異可能會對浮游動物的優勢種的數量變化與空間分布格局造成顯著的影響[22-23]。2018年春季浮游動物大都分布于舟山近海水團,而較少在杭州灣內水團出現,這可能與沖淡水量較小導致的杭州灣內營養鹽的總體水平較低有關。2019年較大的沖淡水量,引起了較多的浮游動物在杭州灣內聚集,同時,舟山近海發生了較強的浮游植物藻華,也引起了部分浮游動物(比如夜光蟲)在近海聚集。因此,2018年和2019年的浮游動物空間分布格局主要體現了餌料對生物聚集的調控。周孔霖[24]也發現了餌料濃度影響浮游動物的生長和聚集,本文的研究結論與其結果一致。
4.2 浮游動物總生物量年際差異的原因
本文結果顯示,2018年春季SSS高、SST低、Chl a濃度低,而浮游動物總豐度大,與此相反,2019年SSS較低、SST較高、Chl a濃度較高,而浮游動物總豐度低(圖2,圖4)。根據食物匹配和溫度有利于生理生長的理論[25-28],2018年的浮游動物總豐度應該較2019年低。本文的研究結果與常規的理論推測相反,造成這一現象可能與兩個因素有關。
一是2019年春季長江沖淡水量異常偏高,為有害赤潮藻暴發提供了豐富的營養鹽,導致適宜生長的夜光蟲暴發。夜光蟲雖然本身不含毒素,但其暴發后,導致其他浮游生物生存環境更加惡劣(比如缺氧、擠占生存空間、浮游動物適宜的餌料數量減少),從而導致2019年春季高營養物質流入有害赤潮藻類,導致大量有害浮游植物不能被浮游動物有效利用,造成浮游動物數量偏低。很多研究[14,29-30]也表明,發生赤潮藻華時,浮游動物生物量不升反降,與本文的研究結果一致。
二是氣候條件所致,2018年春季呈現明顯的拉尼娜現象[31],溫度總體偏低,可能導致偏冷水性浮游動物數量較多,而偏暖溫性浮游動物數量較少,亦有研究(http://www.bom.gov.au/climate/enso/)表明,2019年春季呈現厄爾尼諾現象[31],導致原來棲息于此的偏冷水性浮游動物大量死亡,從而導致總的浮游動物數量偏低。如果這兩年推論成立,本研究結果表明,人類活動和氣候變化會對浮游動物種群數量變化產生顯著的影響,對優勢種的交替具有重要的影響。對于本研究推論的廣泛適用性,還需要結合其他年份的調查數據,進一步的調查驗證。
5 結論
本研究根據2018-2019年春季的浮游動物豐度數據和環境因子數據,發現舟山近海海域存在3個水團:杭州灣內水團、舟山本島上升流水團和舟山近海水團。分析發現,不同水團對春季浮游動物群落結構地理分布格局影響顯著,且對浮游動物群落結構的年際變化具有顯著影響,鹽度是影響春季浮游動物群落結構空間差異的主要因素,溫度、葉綠素a濃度是春季浮游動物群落結構年際差異的主要因素。其次,夜光蟲等浮游生物的暴發對浮游動物分布會造成重要影響。
致謝:感謝“浙普漁43 019”號全體船員在出海采樣過程中的幫助,A17海洋資源與環境班級董軍、官道伊、江丹妮、張晗、龍麗梅等同學參與部分實驗,謹致謝忱。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
