刺槐體細胞胚發生的同步化調控
2020-07-23 04:35:36習洋孫宇涵李云
河北大學學報(自然科學版) 2020年3期
關鍵詞:質量
習洋,孫宇涵,李云
(1.國家知識產權局專利局專利審查協作北京中心 醫藥生物部,北京 100160;2.北京林業大學 生物科學與技術學院,北京 100083)
體細胞胚發生是體細胞向胚胎發生途徑轉變的重建過程,也是證明細胞全能性理論的經典實例.
建立一個高頻率、同步化發生的體細胞胚培養系統,對深入揭示植物胚胎發育機制以及在植物遺傳轉化和人工種子等方面應用均很重要[1].早在1973年Okamura[2]就在胡蘿卜細胞培養中采用低溫處理和饑餓處理相結合的方法獲得了較好的細胞同步化結果.陳春玲[3]通過控制培養基中2,4-二氯苯氧乙酸(2,4-dichlorophenoxyacetic acid,2,4-D)的質量濃度來調控龍眼體胚的發育,研究表明經過不同質量濃度2,4-D處理后的材料染色體數目正常,且仍然保持著很強的體細胞胚發生能力,通過對龍眼整個體細胞胚發生過程的研究,根據不同發育階段材料對培養基中2,4-D的質量濃度需求不同,實驗設計了多種同步化培養基并分別獲得了非胚性愈傷組織、胚性愈傷組織Ⅰ( 1 mg/L 2,4-D) 、胚性愈傷組織Ⅱ( 0.8 mg/L 2,4-D) 、胚性緊密結構、不完全胚性緊密結構、球形胚、玻璃化早期子葉形胚、成熟子葉形胚等 8 個階段的同步化調控材料.目前,通過調控培養得到同步化的植物體細胞胚仍是尚未解決的難題,進行過細胞同步化和體細胞胚發生的同步化培養研究的僅有胡蘿卜、水稻、枸杞、龍眼等少數幾種植物[1].
本實驗室前期利用不同時期的刺槐未成熟合子胚誘導愈傷組織,研究了幼胚胚齡和不同植物生長調節劑對于體細胞胚誘導的影響[4],并建立了刺槐胚性細胞懸浮系[5].本文是在已建立了刺槐體細胞培養體系基礎上,進一步對刺槐胚性愈傷組織的體細胞胚發生及同步控制進行了研究,建立了刺槐體細胞胚發生及同步控制的培養體系,以期有助于生理生化和分子水平上的刺槐體細胞胚發生機制研究.
1 材料與方法
1.1 實驗材料
由刺槐55 d胚齡的未成熟合子胚誘導得到的嫩黃綠色、松散易碎、顆粒狀的刺槐胚性愈傷組織由北京林業大學生物科學與技術學院李云課題組提供.
1.2 低溫處理控制體細胞胚同步化
將多次繼代的胚性愈傷組織接種到體細胞胚誘導培養基中,置于4 ℃的低溫條件下分別處理0、7、14、21、28 d后轉移到正常溫度條件下培養,7 d后觀察體細胞胚生長情況,且每7 d統計體細胞胚的發生數目.體細胞胚誘導培養基為MS(Murashige & Skoog)+NAA(naphthylacetic acid) 0.5 mg/L+ BA(benzyladenine ) 0.5 mg/L+2-嗎啉乙磺酸 (MES) 500 mg/L+谷氨酰胺 500 mg/L+水解酪蛋白 500 mg/L+蔗糖30 g/L+瓊脂6 g/L[4].
1.3 蔗糖質量濃度控制體細胞胚同步化
將多次繼代的胚性愈傷組織接種到附加不同質量濃度(0,20,30,50,70 g/L)蔗糖的體細胞胚誘導培養基中,每7 d觀察體細胞胚生長情況,并統計體細胞胚的發生數目. 體細胞胚誘導培養基為MS+NAA 0.5 mg/L+ BA 0.5 mg/L+MES 500 mg/L+谷氨酰胺 500 mg/L+水解酪蛋白500 mg/L +瓊脂6 g/L.
1.4 外源植物生長調節劑質量濃度控制體細胞胚同步化
將多次繼代的胚性愈傷組織接種到附加不同質量濃度(0,0.1,0.3,0.5,1.0 mg/L)2,4-D的體細胞胚誘導培養基中,每7 d觀察體細胞胚生長情況,并統計體細胞胚的發生數目. 體細胞胚誘導培養基為MS+BA 0.5 mg/L+谷氨酰胺 500 mg/L+水解酪蛋白500 mg/L+蔗糖30 g/L+瓊脂6 g/L.
1.5 懸浮培養控制體細胞胚同步化
選擇嫩黃綠色、顆粒狀、生長較快的胚性愈傷組織,置于培養基成分為1/2 MS+MES 500 mg/L+NAA 2.5 mg/L+BA 0.5 mg/L+蔗糖30 g/L的懸浮培養基中,得到分散度較好、穩定性較強且具有胚性的懸浮培養細胞[5].
用吸管吸取懸浮液,將懸浮培養的胚性細胞用50目篩(孔徑0.28 mm)過濾,將留在篩上的細胞團均勻地接種到表面墊有濾紙的MS+NAA 0.5 mg/L+ BA 0.5 mg/L+MES 500 mg/L+谷氨酰胺 500 mg/L+水解酪蛋白500 mg/L+蔗糖30 g/L+瓊脂3 g/L半固體愈傷組織的誘導培養基上,進行看護培養.使用SPSS 17.0進行數據分析,每個處理3個重復.
2 結果與分析
2.1 低溫對體細胞胚發生同步化的影響
將前期誘導得到的胚性愈傷組織在4 ℃條件下低溫處理,經過42 d的恢復培養,發現隨著低溫處理時間的延長,體細胞胚子葉期同步化率逐漸降低(圖1).低溫處理7 d的子葉體細胞胚同步化率最高,為37.3%,而低溫處理14 d的子葉體細胞胚同步化率僅為27.3%,與低溫處理7 d的子葉體細胞胚相比延長低溫處理時間(21 d和28 d)的子葉體細胞胚同步化率均顯著降低(P<0.05).
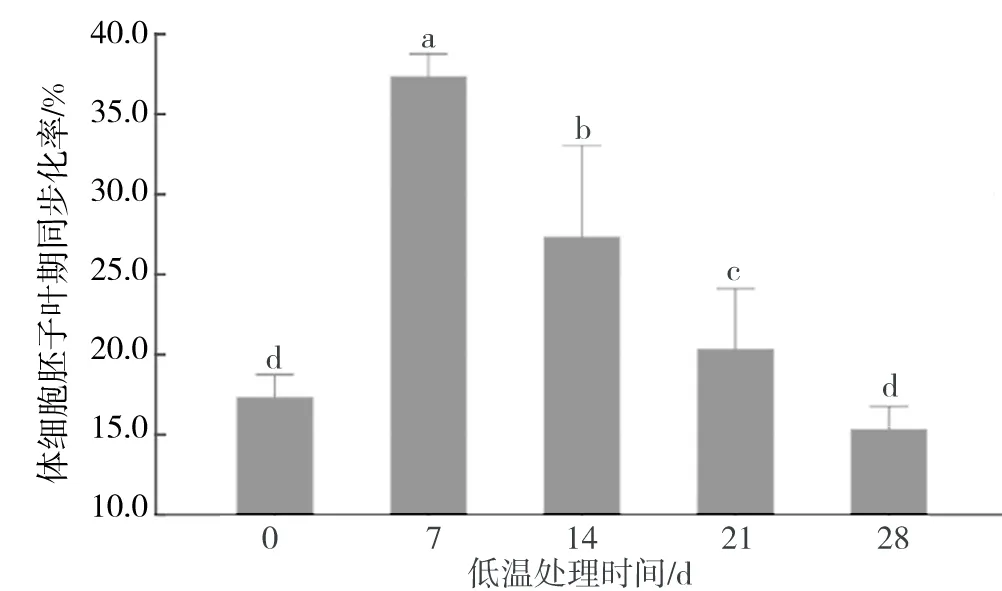
圖1 不同低溫處理時間對體細胞胚子葉期同步化率的影響Fig.1 Effects of different freezing time on the synchronization of cotyledon somatic embryos
實驗觀察發現,低溫處理7 d的胚性愈傷組織顏色接近正常,愈傷組織基部邊緣有輕微褐化的現象.隨著低溫處理時間延長,胚性愈傷組織的褐化現象漸漸加重(表1).當低溫處理21 d時胚性愈傷組織出現明顯的褐化現象,整個愈傷組織塊呈現褐色.而低溫處理28 d時其褐化現象與低溫處理21 d時差別不大,只是個別部位褐化更為嚴重.這一現象可能是由于低溫處理對刺槐胚性愈傷組織造成的傷害主要取決于溫度變化,而當低溫處理溫度一定時,處理時間的長短對刺槐胚性愈傷組織造成的傷害沒有顯著差異.然而,較長的低溫處理時間會抑制刺槐胚性愈傷組織的體細胞胚發生.統計實驗數據,分析發現低溫處理7 d的子葉體細胞胚同步化率與對照有顯著差異 (P<0.05).

表1 不同低溫處理時間對體細胞胚發育的影響
胚性愈傷組織經過低溫處理后畸形胚的發生率明顯降低(表1).徐元紅[6]在平貝母體細胞胚誘導和植株再生的過程中發現,未經低溫處理(0~5 ℃冰箱中處理60 d)的體細胞胚會發育成為畸形胚.Mahesh[7]認為低溫處理與低培養溫度綜合作用,可明顯提高體細胞胚的發生數量,且有利于體細胞胚的正常發育.從表1中可以發現,隨著低溫處理時間的延長,畸形胚的發生率逐漸降低,但下降幅度不大.綜合分析低溫處理實驗結果,在低溫(4 ℃)條件下將胚性愈傷組織處理7 d最有利于刺槐體細胞胚的同步化.
2.2 蔗糖質量濃度對體細胞胚發生同步化的影響
將經過多次繼代的胚性愈傷組織接種到5種不同蔗糖質量濃度的體細胞胚誘導培養基上,培養基為MS+NAA 0.5 mg/L+ BA 0.5 mg/L+MES 500 mg/L+谷氨酰胺 500 mg/L+水解酪蛋白500 mg/L+瓊脂6 g/L.培養28 d后觀察體細胞胚的發育情況,56 d后統計子葉期體細胞胚的同步化情況和畸形胚的發生情況及體細胞胚的敗育情況.
如圖2所示,在蔗糖質量濃度升高的培養基中誘導得到的體細胞胚其同步化率遠高于對照組(P<0.05).當蔗糖質量濃度為50 g/L時,其子葉體細胞胚的同步化率最高,可達35.0%,與蔗糖質量濃度為30 g/L的對照組的體細胞胚同步化率(17.3%)相比有顯著差異(P<0.05).Bomal[8]在對黑云杉的體細胞胚誘導實驗中發現,滲透壓升高導致的水分饑餓能夠促進體細胞胚的同步化發生.本實驗中得出相同的結論.當培養基中的蔗糖質量濃度升高到50 g/L時,培養基中的碳源豐富,能夠滿足胚狀體發生的需要,與對照組相比,此時的培養基滲透壓升高,水分相對不足,胚性愈傷組織在水分饑餓的條件下漸漸停止增殖分裂,隨后細胞開始分化,向胚狀體方向發展.當培養基中的蔗糖質量濃度降低為20 g/L時,體細胞胚的同步化率也略有升高,為18.3%,與對照組相比差異不顯著.分析原因,可能是由于培養基中的蔗糖質量濃度較低,此時體細胞胚的發育主要受到能量供給的限制,而滲透壓的調節作用處于次要地位造成的.當培養基中無蔗糖存在時,其子葉體細胞胚的同步化率顯著低于有蔗糖條件下的子葉體細胞胚的同步化率.這可能是由于愈傷組織轉移到無糖培養基后,其能量供應不足,組織停止發育,使得體細胞胚生長緩慢甚至褐化死亡.
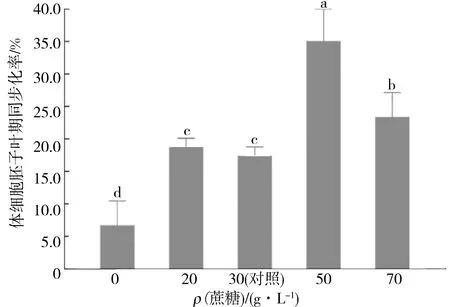
圖2 不同蔗糖質量濃度對體細胞胚子葉期同步化率的影響Fig.2 Effects of different sucrose mass concentration on the synchronization of cotyledon somatic embryos
在植物體細胞胚的誘導培養過程中,長久以來沒有克服的技術難題是無法抑制畸形胚的發生,本研究中也存在同樣的問題.如圖3所示,畸形胚的發生率隨著蔗糖質量濃度的改變而發生變化.當培養基中的蔗糖質量濃度過高或過低時都會導致畸形胚的發生率顯著升高.培養基中蔗糖質量濃度降為0時,畸形胚發生率達到最高,為78.3%,分析原因,可能是因為在不含蔗糖的培養基中培養基滲透壓過低,供應細胞合成的碳源不充足等原因導致的.然而,統計分析顯示培養基中蔗糖質量濃度為20和50 g/L時體細胞胚畸形率彼此之間無顯著性差異.
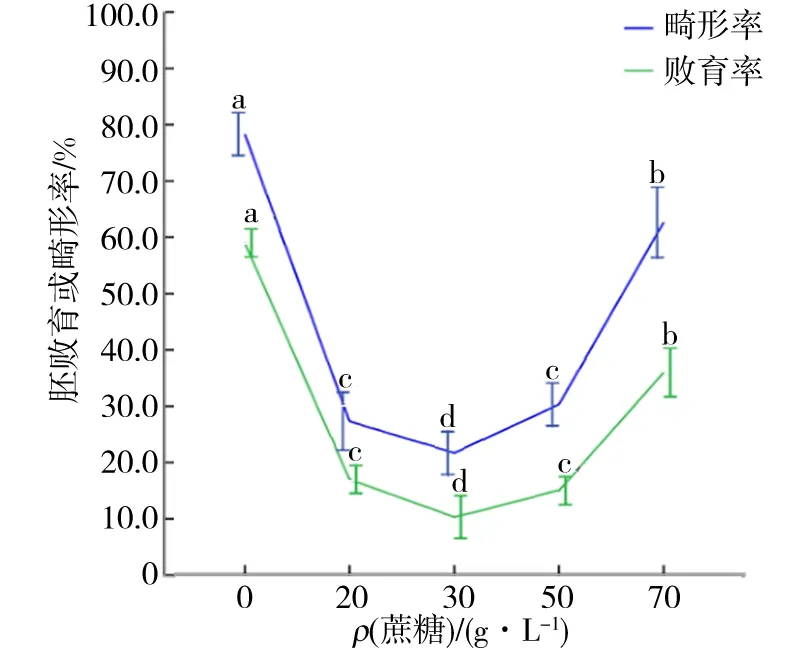
圖3 不同蔗糖質量濃度對體細胞胚敗育和畸形發生的影響Fig.3 Effects of different sucrose mass concentration on the abortive somatic embryogenesis and abnormal embryogenesis
實驗中發現,體細胞胚的敗育現象存在于體細胞胚誘導、同步化及體細胞胚成苗的全過程中.觀察實驗中處于不同蔗糖質量濃度培養條件下的刺槐體細胞胚的敗育情況,結果發現培養基中蔗糖質量濃度降為0時,敗育率最高,為59.0%(圖3).這可能是由于培養基中不添加蔗糖時胚性細胞難以增殖,而此培養條件適宜非胚性細胞的增殖,此時,愈傷組織中的胚性細胞逐漸喪失胚性,轉化為非胚性細胞,因此在這種無蔗糖的培養條件下,恢復培養物中胚性細胞的體細胞胚發生潛能是不可能的.提高培養基中的蔗糖質量濃度雖然也會使體細胞胚的敗育率增高,但是這種影響遠低于蔗糖質量濃度降低時的影響.當培養基中的蔗糖質量濃度為50 g/L時,體細胞胚的敗育率為15.0%,且大部分敗育的胚狀體都分布在與培養基接觸的部分.
綜合上述實驗結果,培養基中蔗糖質量濃度為50 g/L時最適宜調控刺槐的體細胞胚同步化.
2.3 外源植物生長調節劑質量濃度對體細胞胚發生同步化的影響
將經過多次繼代的胚性愈傷組織接種到含有不同2,4-D的體細胞胚誘導培養基上,培養基為MS+ BA 0.5 mg/L+MES 500 mg/L+谷氨酰胺 500 mg/L+水解酪蛋白500 mg/L+蔗糖30 g/L+瓊脂6 g/L.培養28 d后觀察愈傷組織上體細胞胚的發育情況,培養56 d后統計刺槐子葉期體細胞胚的同步化發生情況和畸形胚的發生情況.
實驗結果如圖4所示,2,4-D不僅對體細胞胚的誘導起決定作用,還可以控制體細胞胚的同步化發育,隨著培養基中2,4-D質量濃度的降低體細胞胚的同步化率相應升高.當培養基中2,4-D的質量濃度為0.1 mg/L時,體細胞胚的同步化率最高,為43.6%.劉選明[9]在對雜交水稻的體細胞胚同步化研究中發現2,4-D能夠影響胚性細胞的活力,高質量濃度的2,4-D顯著抑制體細胞胚的發生和發育,使胚性愈傷組織進入周而復始的增殖周期而無法分化成胚狀體.當培養基中不含2,4-D時,體細胞胚的同步化率有所下降,但此時的同步化率與2,4-D質量濃度為0.3 mg/L時差異不顯著.
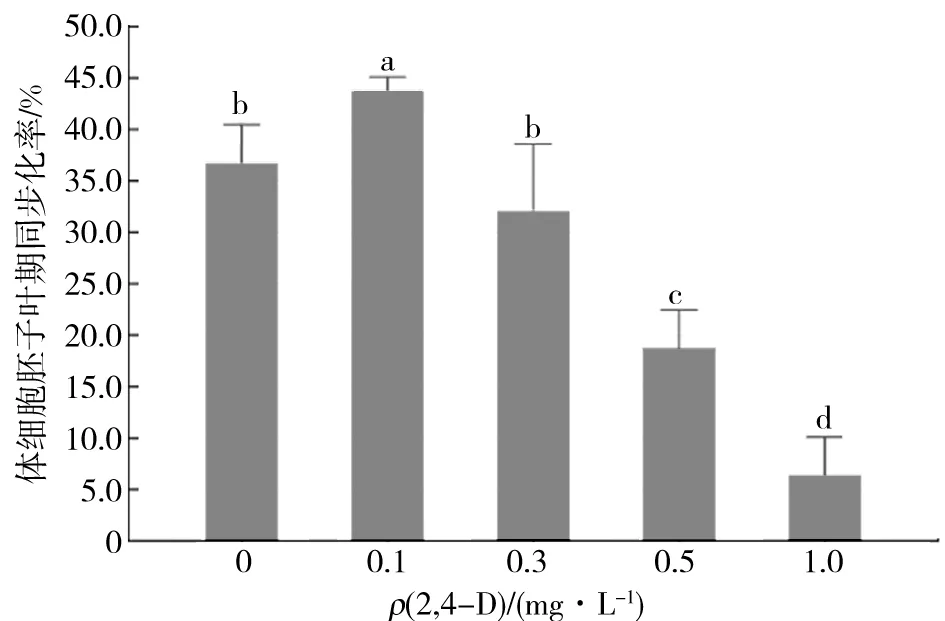
圖4 2,4-D質量濃度對體細胞胚子葉期的同步化率的影響Fig.4 Effects of different 2,4-D mass concentration on synchronization of cotyledon somatic embryo
袁澍等[10]研究發現2,4-D通過與細胞內的激素受體蛋白結合而對某些控制植物體胚發生的特異表達基因起調控作用,它可以激活某些控制細胞胚性的相關基因,增加胚胎發生特異蛋白的表達量;此外,2,4-D對畸形胚的發生有較好的抑制效果,在適宜的質量濃度下,2,4-D能夠抑制多種畸形體細胞胚的發生.本研究也得出了相同的結論,隨著培養基中2,4-D質量濃度的升高,畸形胚的發生率逐漸降低.但是2,4-D質量濃度為0.1~0.5 mg/L時的畸形胚發生率差異并不顯著(圖5).
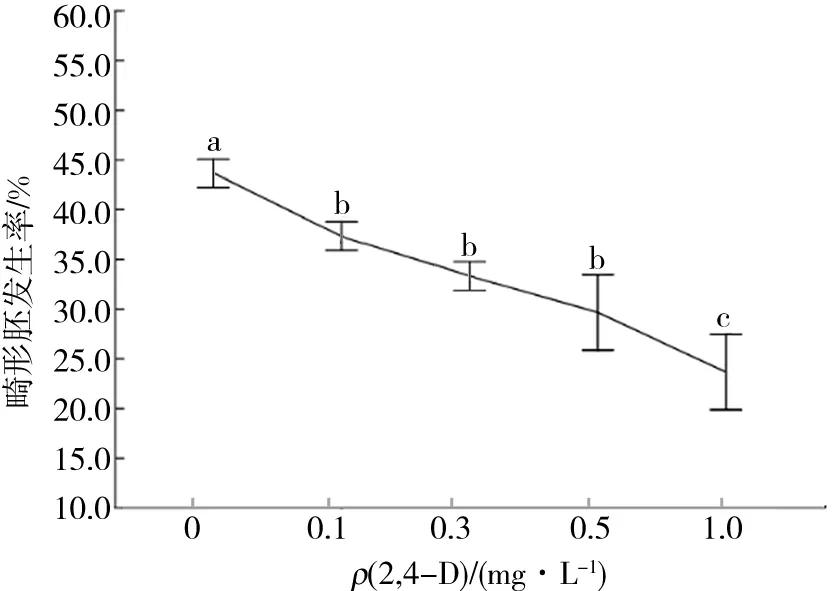
圖5 2,4-D質量濃度對畸形胚發生率的影響Fig.5 Effects of 2,4-D mass concentration on abnormal embryogenesis
綜合上述實驗結果,培養基中的2,4-D質量濃度為0.1 mg/L時最適宜體細胞胚同步化的調控.
2.4 懸浮培養細胞的體細胞胚同步化誘導
圖6a-i顯示了各發育階段的同步化體細胞胚.將懸浮培養的胚性細胞用50目篩(孔徑0.28 mm)過濾,將留在篩上的細胞團接種到鋪有濾紙的半固體體細胞胚誘導培養基上(圖6g),培養基為MS+NAA 0.5 mg/L+ BA 0.5 mg/L+MES 500 mg/L+谷氨酰胺 500 mg/L+水解酪蛋白500 mg/L+蔗糖30 g/L+瓊脂3 g/L.培養7 d后觀察到細胞團的數量明顯增多,14 d左右可以發現有少量體細胞胚發生(圖6h).培養21 d左右,球形胚大量發生(圖6i),占到總胚狀體數目的67.3%.在得到的胚狀體中,有部分球形胚進入了魚雷胚期和子葉胚期,但二者的數目僅占總胚數的20.4%和12.3%(表2).隨著總胚數的逐漸增加,球形胚數量逐漸減少,魚雷期和子葉期的體細胞胚數量逐漸增加,但球形胚期的體細胞胚仍占主要地位.實驗中沒有觀察到典型的大量心形胚的發生時期.這可能是由于心形胚的發育時期較短,不易觀察.培養至28 d時,總體細胞胚數的增加開始減緩,球形胚向魚雷胚和子葉胚的轉化也逐漸停滯,畸形胚的數量顯著增加.分析原因可能與營養物供給不足有關.將培養21 d的球形胚轉接至新鮮的墊有濾紙的誘導培養基上繼續培養能夠有效降低畸形胚的發生比例,得到大量同步化的正常發育的體細胞胚.

a.同步化的球形體細胞胚;b.同步化的魚雷胚;c.同步化的子葉胚;d.起始密度為2 g時的懸浮培養細胞團;e.懸浮培養 21 d后的胚性細胞團;f.懸浮培養21 d后的胚性培養物;g.接種到看護培養基上的懸浮培養物;h.看護培養14 d后 得到的球形胚;i.看護培養21 d后得到的球形胚和個別魚雷胚.圖6 同步化的各發育階段的體細胞胚Fig.6 Synchronized somatic embryo at different developmental stages
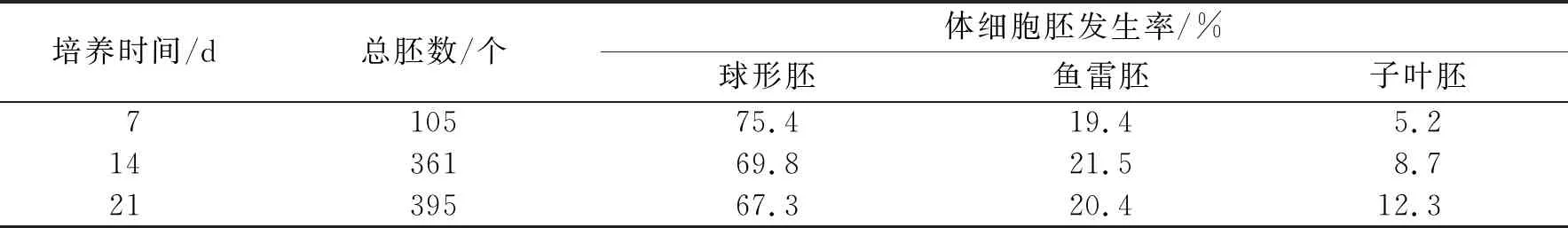
表2 看護培養下不同時期的體細胞胚發生率
表中數據為每個時期統計3個培養皿中體細胞胚數目.
3 討論
1)低溫可以導致細胞發育的停滯,即形成溫度傷害,這種溫度傷害常可以用來調控體細胞胚的同步化發生.李鳳榮[11]研究了不同低溫處理時間下蠶豆根尖細胞分裂的同步化情況,結果發現,低溫處理時間對細胞的同步化率有顯著影響.周愛文[12]的研究表明,溫度對于植物細胞的有絲分裂具有顯著的影響,在一定溫度范圍內,隨著溫度的下降細胞分裂速度減緩、細胞周期延長,相應地延長了細胞分裂期的時間,分裂指數相應提高,細胞的同步化率也相應升高.當培養溫度恢復至正常水平時,大量細胞即進入DNA的合成期,隨后出現一個明顯的細胞分裂高峰.低溫處理可以提高紅豆杉懸浮培養細胞的同步化程度,以4 ℃處理24 h后,再恢復培養24 h,紅豆杉懸浮細胞分裂指數可達到10.26%[13].毛春娜[14]在對半夏懸浮培養細胞的同步化調控中采用了低溫處理的方法,并發現低溫處理可以顯著提高細胞的同步化率,有效地提供高細胞分裂指數.本實驗結果也顯示,低溫處理可以促進刺槐體細胞胚的同步化發生,且低溫處理后的刺槐體細胞胚大小均一、發育良好,畸形胚的發生率明顯降低.低溫處理操作簡單、方便,且短暫的低溫處理一般不會引起細胞DNA和染色體的變異,目前,低溫培養在動物細胞和植物細胞培養領域是倍受青睞的同步化調控方法.
2)蔗糖作為碳源,為植物細胞的呼吸代謝提供必要的能源,同時蔗糖還是培養物與培養環境維持滲透壓平衡的主要調節者.在植物細胞培養方面,目前的研究大多集中在蔗糖對植物胚性愈傷組織的影響上,而針對蔗糖在體細胞胚同步化調控方面的研究卻很少.本實驗發現,在添加不同質量濃度蔗糖培養基中,刺槐體細胞胚發生的同步化水平存在顯著差異.這說明蔗糖質量濃度對刺槐體細胞胚的同步化發生有著重要影響.分析原因,可能是由于不同發育階段的植物體細胞胚對滲透壓有著不同的需求,因此可以通過調節培養基中蔗糖的質量濃度來調節滲透壓從而控制體細胞胚的發育,使其停滯在某一發育階段[15].但是長期處于高蔗糖質量濃度的培養基中會極大地降低愈傷組織的胚胎發生能力,導致畸形胚的發生和胚狀體敗育.本實驗綜合了蔗糖質量濃度對畸形胚和胚狀體敗育的影響,認為培養基中蔗糖質量濃度為50 g/L時最適宜調控刺槐的體細胞胚同步化,此時的體細胞胚同步化率達到35.0%.
3)在調控植物體細胞胚的同步化發育方面外源植物生長調節劑也發揮著重要作用.楊映根[16]在青杄體細胞胚發生的實驗中發現,在含有脫落酸(ABA)的分化培養基中青杄子葉期體細胞胚的同步化率可達80%以上.Merkle[17]在對針葉樹體細胞胚的同步化發生研究中也發現,經過ABA處理后可產生大量高質量同步化的成熟體細胞胚.2,4-D是誘導愈傷組織必不可少的生長激素[18],2,4-D對內源激素的調節和平衡起重要作用[19],57.7%的雙子葉植物和幾乎所有的單子葉植物在體細胞胚的誘導階段均使用了2,4-D[20].低質量濃度的2,4-D不僅能夠促進體細胞胚的發生,而且還能夠調控體細胞胚的同步化.在本實驗中,低質量濃度的2,4-D能夠顯著促進刺槐體細胞胚的同步化發生,且同步化率可達43.6%.然而2,4-D促進體細胞胚同步化的機制尚不清楚,還有待于進一步的研究.
4)植物細胞懸浮培養具有促進胚性細胞大量快速增殖、形成均一的小細胞團和單細胞等優點,是建立體細胞胚同步化的良好實驗體系[21].本實驗通過建立刺槐的胚性細胞懸浮培養體系,探索了植物生長調節劑和培養方法對體細胞胚同步化的影響.細胞懸浮培養對刺槐體細胞胚的同步化調控是有效的,與固體培養相比,懸浮培養時胚性愈傷組織能夠更均勻更充分地吸收營養[22-23],有利于體細胞胚胎發生的同步化[24].在一些植物的體細胞胚同步化誘導中,懸浮培養往往和ABA處理[25-26]、肌醇饑餓[27]、密度梯度離心[28-29]等方法結合使用,以實現體細胞胚的同步發育.本實驗中將懸浮培養和半固體看護培養相結合,將懸浮培養16 d的胚性細胞接種到墊有濾紙的半固體培養基上,使其與培養基充分均勻接觸,解決了固體培養時通常存在的培養物與培養基接觸不均勻的問題.結果表明,當懸浮培養物在半固體培養基上培養21 d左右時,球形胚大量發生,占到總胚狀體數目的67.3%.
4 展望
目前,能夠成功進行體細胞胚同步化的植物品種還很少,對于植物體細胞胚同步化條件的優化還有待進一步研究. 本研究探索了刺槐體細胞胚同步化發生的培養條件,并綜合畸形胚和胚狀體敗育的情況得出了各方法誘導刺槐體細胞胚同步化發生的最適條件,希望能夠為其他植物種類的體細胞胚同步化調控提供參考.通過對體細胞胚發生中調控基因的研究,進一步明確外源培養條件在體細胞胚發生各階段的調控機理,必將對于更多植物品種的體細胞胚發生同步化,以及利用體細胞胚技術制備轉基因受體奠定基礎,為實現通過基因修飾進行體細胞胚同步化控制創造條件.
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
