PDIA3對人Jurkat T細胞TCR信號通路的調控
2020-07-23 04:35:38周云劉海龍靳照宇
河北大學學報(自然科學版) 2020年3期
周云,劉海龍,靳照宇
(1.河北大學 生命科學學院,河北 保定 071002;2.北京蛋白質組研究中心 北京 102206 )
T細胞是免疫系統的重要組成部分,在細菌、病毒等病原體的免疫應答,抵抗癌癥、感染以及自身免疫性疾病方面起著至關重要的作用[1-2].T細胞通過識別由抗原呈遞細胞捕獲的外源多肽段, 從而激活T 細胞及下游的一系列免疫反應.活化的T細胞可以誘導包括ζ-鏈相關蛋白70(zeta-chain associated protein 70, ZAP70)磷酸化、細胞因子分泌、轉錄因子NF-κB(nuclear factor kappa-B)、NFAT(nuclear factor of activated T-cells)激活等一系列生物學過程[3].
TCR(T cell receptor)是T細胞表面的標志性受體,它與分化抗原簇3(cluster of differentiation 3, CD3)分子結合,形成TCR-CD3復合物.TCR通路在T細胞激活過程中具有重要作用,特異性的抗體識別TCR后,會激活TCR通路下游的一系列蛋白,調節T細胞的分化和活化等.
通過本實驗室前期的工作,在TCR復合物中發現了調控T細胞受體信號通路的有效分子PDIA3,該分子是蛋白質二硫鍵異構酶A3前體(protein disulfide isomerase A3 precursor, PDIA3),屬于二硫鍵異構酶家族[4].PDIA3是一種巰基-氧化還原酶伴侶蛋白[4],分子質量為58 ku,是信號轉導和轉錄激活因子3信號通路中的一部分[5],在多種細胞,尤其是免疫細胞中表達水平較高,參與細胞信號轉導、主要組織相容性復合物組裝[6-7]、血小板功能[8]、轉錄[9]、骨發育[10]及凋亡誘導等多種重要的生物學過程.有研究表明,PDIA3在卵巢癌、乳腺癌、子宮癌、肺癌和胃癌等多種癌組織中都有表達[4]. 到目前為止,幾乎沒有相關研究表明PDIA3表達與TCR信號傳導途徑之間的關系.由于PDIA3在免疫細胞中高表達,所以筆者提出假說:PDIA3作為重要的免疫調控靶點,在TCR下游通路的級聯反應發揮重要作用.
本文旨在研究PDIA3對Jurkat T細胞中TCR下游通路的調控作用.利用電穿孔法將PDIA3基因的siRNA成功導入Jurkat T細胞中并成功敲低了PDIA3蛋白表達,發現敲低了PDIA3蛋白能夠下調ZAP70蛋白的磷酸化水平,抑制NF-κB信號通路,下調繁殖標記物CD69的表達以及IL-2的分泌,為今后深入研究PDIA3與TCR信號通路下游相關功能蛋白的相互作用提供了重要基礎.
1 材料與方法
1.1 實驗材料
1.1.1 細胞
Jurkat T和Jurkat-NF-κB T細胞株保存于本實驗室.
1.1.2 干涉序列
PDIA3基因的siRNA-794(5'-GGACAAGACUGUGGCAUAUTT-3')以及siRNA-NC(negative control siRNA, siNC)由蘇州吉瑪基因合成.
1.1.3 試劑材料
胎牛血清(fetal bovine serum,FBS)(Gibco USA);1640培養基(Gibco USA);放射免疫沉淀法緩沖液(radioimmunoprecipitation assay buffer, RIPA);電轉緩沖液(HyClone EPB1);電轉杯OC-100(MaxCyte OC-100 processing assembly);一抗:Y319 Rabbit mAb 抗體(CST, 2701S)、PDIA3 Rabbit mAb抗體(proteintech, 15967-I-AP)、ZAP70 Rabbit mAb抗體(CST, 27054S)、甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase, GAPDH) mouse mAb抗體(北京傲銳生物科技有限公司,TA-08);二抗:山羊抗兔IgG/辣根過氧化物酶標記(中杉金橋,ZB-2301);山羊抗小鼠IgG/辣根過氧化物酶標記(中杉金橋,ZB-2305);CD69抗體(invitrogen, 12-0699-42);ONE-Glo熒光素酶報告基因檢測系統(Promega, E606B);The BD OptEIATMReagent Set B(BD, 55-534);BD OptEIATMHuman IL-2 酶聯免疫吸附測定(enzymelinked immunosorbentassay,ELISA) Set(BD, 555190).
1.2 實驗方法
1.2.1 細胞培養
用含有體積分數10% FBS的1640培養基,體積分數 5% CO2, 37 ℃條件下恒溫培養;細胞生長至培養瓶70%開始下一步轉染實驗.
1.2.2 細胞轉染
收取約2×107T細胞,磷酸鹽緩沖溶液(phosphate buffer saline, PBS)清洗1遍,用50 μL電轉緩沖液重懸細胞.將細胞懸液與15 μL siRNA(20 μmol/L)混勻,加到電轉杯OC-100中,根據電轉儀操作手冊進行電轉.結束后,立即取出細胞,移至T25細胞培養瓶中,并吹成液滴狀,放入培養箱中孵育,得到2組干涉后的細胞分別是Jurkat-siNC T和Jurkat-si794 T.孵育40 min后向細胞培養瓶中加入10 mL培養基,并放入培養箱中繼續培養48 h,用于后續的功能實驗.
1.2.3 Western blotting檢測
設置CD3抗體質量濃度梯度分別為10、100、200 ng/mL 3組,每組中加1 μg/mL的CD28抗體協同刺激細胞,得到3組CD3/CD28抗體混合溶液(還有1組不加抗體刺激).收取Jurkat-siNC T細胞和Jurkat-si794 T細胞,用抗體混合溶液于37 ℃刺激細胞20 min.
取抗體刺激的細胞(~2×106)加入RIPA并超聲破碎提取蛋白,取12 μg蛋白經體積分數10% SDS-PAGE電泳,在Bio-Rad電轉移系統中將凝膠蛋白帶轉移到聚偏二氟乙烯膜上,50 g/L脫脂牛奶室溫封閉1 h,利用兔抗人pZAP70(Y319)、ZAP70、PDIA3抗體和鼠抗人GAPDH抗體分別作為一抗,4 ℃過夜孵育,TBST洗3次膜,每次10 min,辣根過氧化物酶標記的羊抗兔IgG、羊抗鼠IgG分別作為二抗室溫孵育90 min,TBST洗3次,Western blotting分析T細胞中目的蛋白表達情況.
1.2.4 流式細胞術檢測CD69的表達
設置CD3抗體質量濃度分別為10、200、500、1 000 ng/mL,均加1 μg/mL的CD28抗體協同刺激細胞.將Jurkat-siNC T細胞和Jurkat-si794 T細胞用CD3/CD28抗體刺激12 h后,再用藻紅蛋白標記的CD69抗體和isotype來識別CD69分子,4 ℃冰箱避光放置30 min,最后通過流式細胞儀檢測CD3/CD28抗體激活的T細胞中CD69的表達情況.
1.2.5 ELISA試劑盒檢測IL-2分泌情況
本實驗設置1個實驗孔和2個副孔,重復3次.用100 μL CD3抗體(5 μg/mL)提前包被96孔板,4 ℃過夜.將包被的96孔板用PBS洗3次.收取Jurkat-siNC T細胞和Jurkat-si794 T細胞,用CD28抗體稀釋液(1 μg/mL)重懸細胞,以每孔約2×105的細胞密度接種到96孔板中.CD3/CD28抗體刺激24 h后,收取上清液,通過ELISA試劑盒檢測細胞上清液中IL-2的分泌情況.
1.2.6 Luciferase assays檢測NF-κB信號通路
本實驗設置1個實驗孔及2個副孔,重復3次.設置CD3抗體質量濃度分別為1、3、5 μg/mL,均加1 μg/mL的CD28抗體協同刺激細胞.將Jurkat-NF-κB-siNC T細胞和Jurkat-NF-κB-si794 T細胞以每孔約2×105的細胞密度接種到96孔板中,用CD3/CD28抗體刺激6 h后,通過ONE-Glo熒光素酶報告基因檢測系統檢測細胞中的熒光信號強度.
1.2.7 數據分析
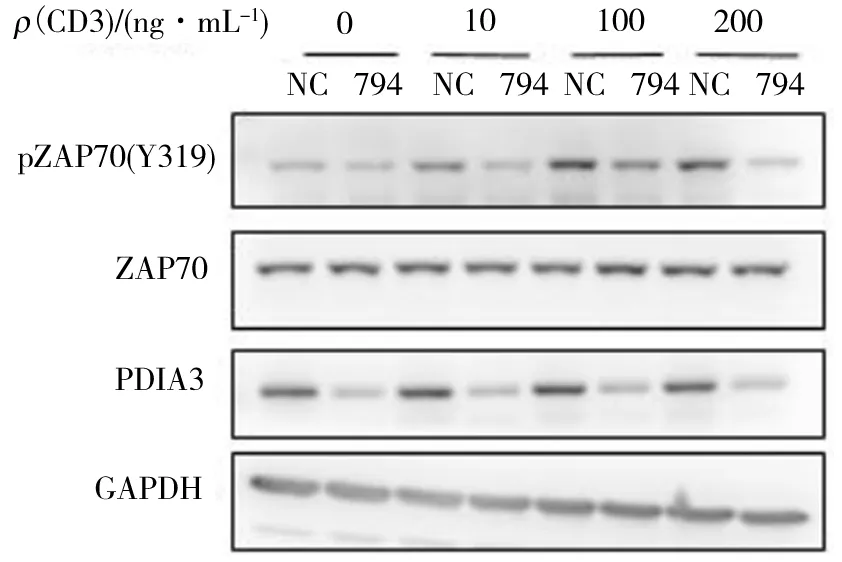
NC.siNC; 794. si794.下同. 圖1 PDIA3調節CD3/CD28抗體激活的ZAP70 蛋白的磷酸化水平Fig.1 PDIA3 regulates the phosphorylation of ZAP70 protein by CD3/CD28 antibodies
使用GraphPad Prism 5進行數據統計分析及柱形圖的繪制.組間比較采用雙尾Student’s t-檢驗,當P<0.05時,組間差異顯著.
2 結果
2.1 敲低PDIA3抑制ZAP70蛋白磷酸化
ZAP70 是一種細胞質蛋白酪氨酸激酶,屬于 Syk 蛋白家族.ZAP70 主要在 T 細胞和自然殺傷細胞中表達,是 T 細胞受體活化所必需的蛋白.為了檢測PDIA3蛋白是否對ZAP70蛋白的磷酸化修飾有影響,進行了siRNA細胞電轉染實驗,電轉后48 h,用不同質量濃度的CD3/CD28抗體協同刺激細胞.通過前期預實驗發現,當CD3抗體質量濃度大于200 ng/mL時,ZAP70蛋白的磷酸化修飾水平(pZAP70(Y319))在實驗組和對照組中幾乎無差別.猜測可能是CD3質量濃度過高,導致T細胞活化效率太高.適當降低CD3抗體的工作質量濃度,分別用10、100、200 ng/mL CD3抗體和CD28抗體激活20 min,結果如圖1所示.由圖1可知,成功敲低了PDIA3的蛋白表達,而且不同質量濃度的CD3抗體刺激對ZAP70蛋白表達沒有影響.在CD3和CD28抗體的刺激下,對照組pZAP70(Y319)蛋白表達水平明顯提高,并且與CD3抗體的質量濃度呈現正相關,表明在抗體刺激下,ZAP70蛋白的磷酸化水平提高,但是在敲低了PDIA3蛋白之后,ZAP70的磷酸化水平幾乎沒有變化. 所以敲低了PDIA3蛋白會抑制ZAP70蛋白磷酸化,從而會影響T細胞活化.
2.2 敲低PDIA3抑制CD69的表達
CD69是T細胞活化后最早表達的表面抗原,是T細胞活化的早期標志物,通常在抗體刺激1~2 h后即可在T細胞表面發現CD69分子.為了探究PDIA3是否會對CD69的表達產生影響,用CD3/CD28抗體刺激細胞,由于前期實驗發現當CD3抗體質量濃度大于1 μg/mL時,超出儀器檢測范圍,所以適當降低CD3抗體的工作質量濃度.設置了不同質量濃度的CD3(10, 200, 500, 1 000 ng/mL)加CD28(1 μg/mL)抗體協同刺激Jurkat-siNC T細胞和Jurkat-si794 T細胞,刺激 12 h后,加入CD69染色抗體來識別并標記細胞膜上的CD69,通過流式細胞術檢測T細胞膜上的CD69的表達情況,結果如圖2所示.由圖2可知,當CD3抗體質量濃度為0也就是T細胞沒有被激活的狀態下,Jurkat-si794 T細胞與Jurkat-siNC T細胞的CD69表達量無顯著性差異(P>0.05),加入CD3/CD28抗體刺激T細胞后,2組細胞的CD69表達量升高,同時Jurkat-si794 T細胞組的CD69表達量顯著低于Jurkat-siNC T細胞組(P<0.05),表明在敲低了PDIA3蛋白會影響CD69的表達.當CD3抗體質量濃度為200 ng/mL時,2組的CD69表達量差異最大.所以在T細胞未活化情況下,PDIA3對CD69的表達沒有影響,當T細胞活化后CD69表達顯著升高,而敲低PDIA3會對T細胞活化通路上CD69的表達產生抑制作用.
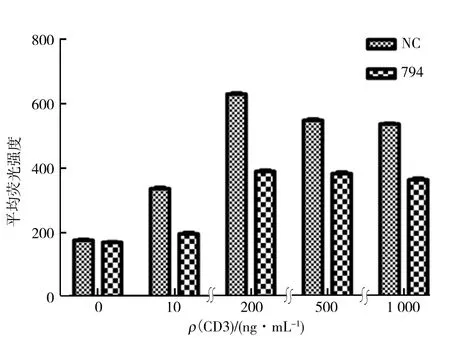
圖2 PDIA3對CD3/CD28抗體激活的Jurkat T 細胞CD69表達的調控Fig.2 PDIA3 regulates the expression of CD69 by CD3/CD28 antibodies in Jurkat T cells
2.3 敲低PDIA3抑制IL-2的表達
IL-2是一種T細胞生長因子,通過TCR信號通路可以調控IL-2的基因表達,屬于TCR信號通路活化下游的一個分子,能促進T淋巴細胞增殖和分化,同時能誘導和增強B細胞、單核巨噬細胞、自然殺傷細胞的活力等,在調控T細胞免疫應答中發揮著重要作用.因此,將Jurkat-siNC T細胞和Jurkat-si794 T細胞用CD3(5 μg/mL)加CD28(1 μg/mL)抗體刺激24 h后,收集細胞培養上清液,通過ELISA試劑盒檢測培養基中的IL-2分泌情況,結果如圖3所示.由圖3可知,敲低PDIA3可顯著抑制IL-2的表達(P<0.01),所以敲低PDIA3后可能對T細胞的生長分化產生影響.
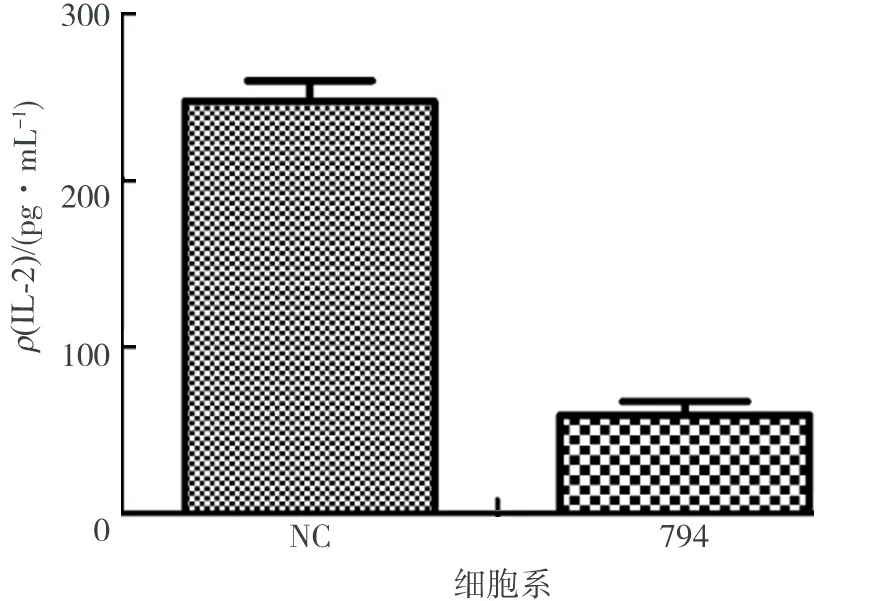
圖3 PDIA3對CD3/CD28抗體激活的 Jurkat T細胞中IL-2分泌的調控Fig.3 PDIA3 regulates the secretion of IL-2 by CD3/CD28 antibodies in Jurkat T cells
2.4 敲低PDIA3對NF-κB信號通路的作用
NF-κB是一種重要的、普遍存在的轉錄因子,主要參與免疫調節及炎癥反應,淋巴細胞發育,細胞增殖、分化、凋亡,腫瘤形成等.如果NF-κB信號通路調控失衡,會導致多種人類疾病,如慢性炎癥、腫瘤和原發性免疫缺陷病等.為了探究PDIA3是否參與調控NF-κB信號通路,用CD3/CD28抗體刺激細胞,實驗發現當CD3抗體質量濃度小于1 μg/mL時,抗體對于細胞NF-κB信號通路的激活作用并不明顯.于是適當提高CD3抗體的工作質量濃度,分別用1、3、5 μg/mL CD3抗體和CD28抗體刺激細胞,通過熒光素酶報告基因檢測系統檢測細胞中的熒光信號強度,結果如圖4所示.由圖4可知,當不加入抗體刺激時,siNC組與si794組的熒光活性較弱且熒光強度無顯著性差異(P>0.05),加入CD3/CD28抗體激活后,實驗組和對照組熒光強度顯著增強,不同質量濃度梯度的CD3抗體刺激細胞,均可以激活NF-κB信號通路,但是敲低了PDIA3后,其熒光強度相比較于NC組具有顯著性差異(P<0.01),所以敲低PDIA3能夠抑制NF-κB信號通路.
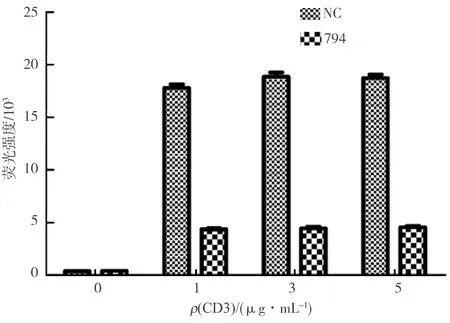
圖4 PDIA3對CD3/CD28抗體激活NF-κB信號通路的調控Fig.4 PDIA3 regulates the NF-κB signaling by CD3/CD28 antibodies
3 討論
PDIA3是PDI家族的成員,主要定位于內質網.一些早期報道發現,PDIA3在腫瘤發生、細胞增殖、細胞凋亡、腫瘤轉移、血管生成及化學抗性中發揮著重要作用[11].有研究表明,早期宮頸癌預后不良與PDIA3的下調有關[12].但是,在乳腺癌發生過程中,發現PDIA3表達上調[13].然而PDIA3在免疫組織中的功能研究尚不清楚.
在本研究中,利用電轉干涉在Jurkat T細胞中成功敲低PDIA3蛋白表達.通過CD3/CD28抗體刺激活化T細胞,增強ZAP70蛋白的磷酸化修飾水平,結果表明,敲低PDIA3蛋白會減弱ZAP70蛋白的磷酸化修飾水平,表明PDIA3與T細胞的活化相關,為了進一步驗證PDIA3與T細胞的活化相關,用CD3/CD28抗體刺激T細胞,并檢測早期活化標志物CD69的表達情況.結果表明,在T細胞未活化狀態下,CD69的表達量比較低,且敲低PDIA3對于CD69的表達量沒有影響.當T細胞受到抗體刺激活化后,CD69的表達量顯著升高,敲低PDIA3會抑制CD69的表達.ZAP70蛋白磷酸化水平的變化和CD69表達量變化表明 PDIA3蛋白參與了T細胞活化過程.T細胞的免疫應答往往還與下游IL-2細胞因子的分泌以及遠端NF-κB信號通路相關,所以本文進一步研究了PDIA3與IL-2細胞因子及NF-κB信號通路之間的關系,發現敲低PDIA3會顯著下調IL-2的分泌.在Luciferase assays實驗中,活化的T細胞會激活NF-κB信號通路,而敲低PDIA3能夠抑制NF-κB信號通路,表明PDIA3蛋白會參與調控T細胞免疫應答的下游通路.
本研究結果顯示,在T細胞中敲低PDIA3可以抑制ZAP70蛋白的磷酸化修飾、 CD69分子表達、IL-2分泌及NF-κB信號通路.因此,猜想PDIA3可能與下游的T細胞活化連接蛋白、含有SH2的白細胞蛋白76 kDa和磷脂酶C-γ1等分子的磷酸化修飾有關.但是PDIA3對T細胞受體信號通路的具體調控機制以及對下游轉錄因子的影響是比較復雜的,需進一步對其機制做深入研究.未來本課題組準備進行磷酸化功能實驗研究,檢測PDIA3分子對TCR信號通路下游的上述分子是否存在調控作用,并構建PDIA3過表達質粒,進行功能回補實驗的驗證.本研究結果為今后對PDIA3與TCR下游一些功能蛋白的相互作用和分子機制研究提供重要基礎.
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
鴨綠江(2021年35期)2021-04-19 12:24:18
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年2期)2019-08-23 08:12:08
電子制作(2018年11期)2018-08-04 03:25:42
海峽科技與產業(2016年3期)2016-05-17 04:32:12
