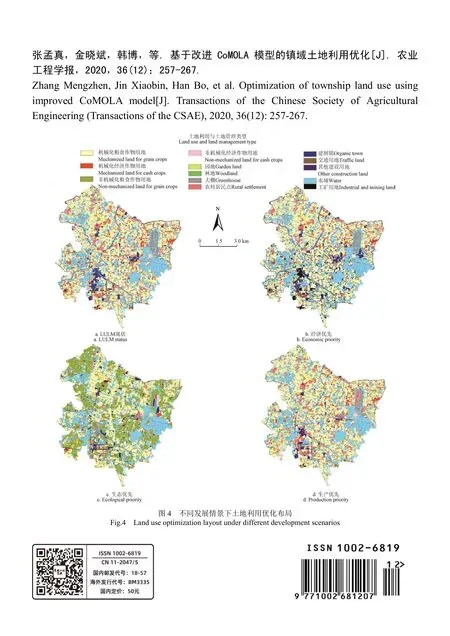
傳統風干牦牛肉加工中肌原纖維蛋白氧化與體外消化性變化
2020-07-25 03:49:28馬紀兵宋艷艷余群力
農業工程學報 2020年12期
馬紀兵,宋艷艷,張 麗,楊 超,韓 玲,余群力
(甘肅農業大學食品科學與工程學院,蘭州 730070)
0 引 言
傳統的風干牦牛肉是將切成條的牦牛肉在低溫條件下自然風干(約 40 d)得到的肉制品。由于受傳統飲食習慣的影響和對傳統文化的傳承,目前這種古老的加工方式在中國牧區仍然流行。而牦牛是這些高海拔牧區的特有畜種,風干牦牛肉含水率約 15%[1],水分活度低于5.0[2],大多數腐敗微生物在此條件下無法生長[3],是牧區傳統的牦牛肉保藏方法。風干牦牛肉蛋白質質量分數為55%[1],屬于高蛋白食品,能夠滿足生活在較高海拔牧區的人們對高蛋白飲食的需求,因此受到牧民們青睞,然而目前對于這種高蛋白肉制品蛋白質消化性還未見報道。因此,有必要探討加工過程中蛋白質的消化率變化以及影響蛋白質水解性的因素,對改善加工工藝和產品質量具有積極促進作用。
肉制品中蛋白質氧化和結構的變化會影響蛋白質被蛋白酶水解的速率,盡管輕度的蛋白質氧化可能導致蛋白質部分解折疊,使蛋白水解酶更容易與底物蛋白結合[4],但在較重氧化條件下,蛋白質與蛋白水解酶結合的位點可能被修飾或蛋白質發生聚集而降低蛋白質被蛋白酶水解的敏感性[5]。在肉制品加工過程中,肌肉暴露于空氣中,氧氣和光等促進了蛋白質氧化程度[6]。另外,肉制品中脂質氧化是促進蛋白質氧化的另一重要原因,脂質氧化的特征是自由基鏈反應,會產生各種氧化副產物,如醛、酮和烷烴[7],α、β-不飽和醛是脂質氧化過程中產生的親電試劑,如丙二醛、4-羥基-2-壬烯醛和丙烯醛可與蛋白質的親核基團反應[8],導致蛋白質的許多結構變化;如修飾的氨基酸側鏈,蛋白質交聯和聚集,這可能會影響蛋白質對胃腸道酶的敏感性[9]。因此,肉類產品中脂質氧化、蛋白質氧化和蛋白質消化之間存在非常密切的關系。風干牦牛肉的加工過程中,生鮮肉長時間暴露于空氣中,其脂質和蛋白質很可能被氧化。但關于風干牦牛肉加工過程中蛋白質氧化修飾及其對蛋白質消化率影響的研究鮮有報道。
本文引入一種特定模擬胃和十二指腸中蛋白質消化的模型[10],以研究風干牦牛加工過程中的營養變化,旨在揭示風干牦牛肉加工過程中脂質氧化、肌原纖維蛋白氧化和蛋白質消化率的變化,并進一步探究蛋白質氧化引起的蛋白質結構變化對蛋白質消化性的影響,以期為改進現有加工技術或進一步開發新加工方法提供參考。
1 材料與方法
1.1 材料與設備
材料:試驗原料肉和風干牦牛肉加工均在甘肅安多清真綠色食品有限公司進行,選取3~4歲公牦牛半膜肌約20 kg作為混合試驗樣品。
試劑:牛血清白蛋白(Bovine Serum Albumin, BSA)購自美國 Sigma公司;胃蛋白酶(牛源):酶活力≥400 U/mg,胰蛋白酶(牛源):酶活力≥1 645 U/mg,α-胰凝乳蛋白酶(牛源):酶活力≥1 500 U/mg,購于美國Sigma Aldrich公司;三氯乙酸(Trichloroacetic Acid,TCA)、2-硫代巴比妥(2-Thiobarbituric Acid, TBA)、四乙氧基丙烷(Tetraethoxypropane, TEP)、2,4-二硝基苯肼(2,4-Dinitrophenylhydrazine, DNPH)、十二烷基磺酸鈉(Sodium Dodecyl Sulfonate, SDS)、5,5′-二硫代雙硝基苯甲酸(5,5′-Dithio Bis-(2-Nitrobenzoic Acid), DTNB)、乙二胺四乙酸(Ethylene Diamine Tetraaceticacid, EDTA)、乙二醇雙(2-氨基乙基醚)四乙酸(Ethylene Glycol Bis(2-Aminoethyl) Tetraacetic Acid, EGTA)、尿素、異硫氰酸胍、硫酸亞鐵、碘酸鈉、無水乙醇、KH2PO4、KCl、K2HPO4、KOH、MgC12、甲醇、氯仿和冰醋酸等均為分析純試劑,購自北京索萊寶科技有限公司。
設備:JRJ-22型實驗室用絞肉機(億翔食品機械有限公司),馬爾文激光粒度儀3000(美國Malvern公司),756P紫外分光光度計(上海光譜儀器有限公司),970CRT熒光分光光度計(蘇州桃梅森科學儀器有限公司),TGL-16M 高速臺式冷凍離心機(長沙湘儀有限公司),XHF-D型高速分散器(寧波新芝生物科技股份有限公司)。
1.2 試驗方法
1.2.1 試驗設計
去除牦牛肉表面的筋膜和可見脂肪,切成橫截面積2 cm×2 cm左右的條狀,長約30 cm,懸掛于風干車間的鐵絲上并且相互之間保持1~2 cm的距離,加工車間可以自然通風并避免陽光直射,在甘肅省夏河縣1-3月較低的環境溫度下進行自然風干(溫度?15~?10℃,相對濕度50%~75%)。將一部分肉條切成小塊,并迅速置于液氮中冷凍作為初始樣品,其余在自然風干 2、4、6、8、10、15、20、25、30和40 d時采集樣品并置于液氮中,后轉移到?80℃超低溫冰箱保存,每個指標測定重復3次。
1.2.2 指標測定
POV(Primary Oxidation Value)測定:按照《食品安全國家標準食品中過氧化值的測定》GB 5009.227-2016方法[11]。
TBARS(Thiobarbituric Acid Reaction Substrates)測定:參考Rasinska等[12]提出的方法并略作修改。準確稱取10.0g樣品,加入20 mL 20%(質量分數)TCA均質。混合液離心6 min(3 500×g,4 ℃),用定性濾紙過濾。取5 mL濾液與5 mL 20 mmol/L TBA溶液混合,置于沸水浴中加熱20 min,以5 mL TCA和5 mL TBA混合液作為對照。冷卻至室溫并在532 nm處測定吸光度。TEP用于標準曲線繪制,以吸光度值為橫坐標x1,TEP溶液濃度為縱坐標y1,標準曲線為y1=0.743x1+0.043(R2=0.999 4)。樣品中TBARS的質量分數表示為mg/kg(以丙二醛計)。
肌原纖維蛋白提取:根據 Wu等[13]的方法并略作修改。稱取肉樣3.0 g,加8倍體積鹽溶液(20 mmol/L磷酸鉀緩沖液,0.1 mol/L KCl,2 mmol/L EGTA,2 mmol/L MgCl2,pH值6.8)均質,4 ℃條件下離心10 min(1 000×g),棄去上清,重復以上步驟 3次。沉淀用 8倍體積100 mmol/L KCl溶液溶解后,4 ℃離心 10 min(1 000×g)棄上清,重復 2次。采用雙縮脲法定量蛋白,以吸光值為橫坐標x2,以牛血清蛋白質量濃度(mg/mL)為縱坐標y2做標準曲線,曲線方程為y2=0.061 3x2+0.000 6(R2=0.999 7)。
羰基測定:依據 Berardo等[6]的方法略作修改。用100 mmol/L KCl溶液將肌原纖維蛋白濃度調整為5 mg/mL,取2份0.5 mL蛋白液,一份加2 mL含0.2%DNPH的2 mol/L HCl溶液,另一份加2 mL 2 mol/L HCl溶液作為對照,室溫下避光反應1 h(每10 min旋渦振蕩一次)。分別添加 2mL 20%三氯乙酸沉淀蛋白,離心(8 000×g,5 min)。沉淀用2 mL乙酸乙酯:乙醇(體積比1:1)清洗,重復3次。分別加3 mL 20 mmol/L磷酸鹽緩沖液(含6 mol/L鹽酸胍,pH值6.5)后置于37℃水浴中保溫30 min溶解沉淀,離心(8 000×g,5 min),在370 nm處測定上清液吸光度。采用下式計算羰基含量
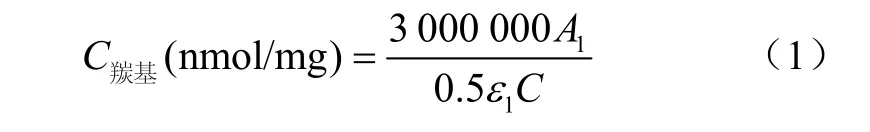
式中A1為 370 nm處吸光值;C為蛋白質質量濃度,mg/mL;ε1為摩爾消光系數 22 000 mol/(L·cm)。
總巰基測定:按照 Cui等[14]的方法稍作修改,用25 mmol/L磷酸鉀緩沖液(pH值6.25)調整蛋白濃度為2 mg/mL。取0.5 mL依次加入2 mL尿素-SDS溶液(含8.0 mol/L尿素,30 g/L SDS,0.1 mol/L磷酸鉀緩沖液,pH值7.4)和0.5 mL 10 mmol/L DTNB試劑(含0.1 mol/L磷酸鉀緩沖液,pH值7.4),室溫下反應靜置15 min,取上清液在412 nm下測定吸光度,25 mmol/L磷酸緩沖液代替蛋白液用于空白對照。采用下式計算總巰基含量
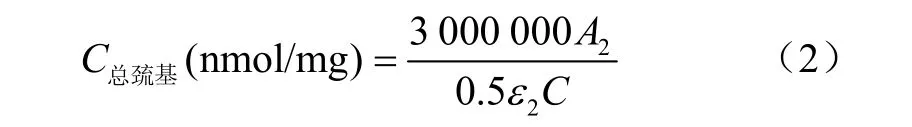
式中A2為 412 nm 處吸光值;ε2為摩爾消光系數13 600 mol/(L·cm)。
二硫鍵測定:按照 Ma等[15]的方法進行測定。用25 mmol/L磷酸鉀緩沖液(pH值6.25)調整蛋白質量濃度為5 mg/mL。取100μL蛋白液與3 mL新配制的NTSB溶液混合,在室溫避光反應25 min,然后在412 nm下測定吸光度,25 mmol/L磷酸鉀緩沖液代替蛋白液用于空白對照。采用下式計算二硫鍵含量
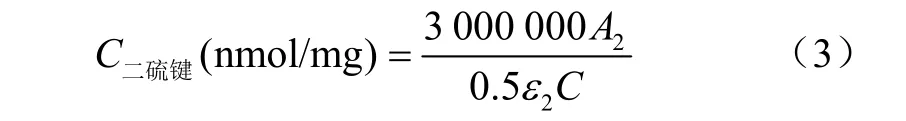
二聚酪氨酸測定:參考崔文斌[16]的方法并略加修改。將蛋白質溶于高離子強度緩沖液(0.6 mol/L,pH值6.0,20 mol/L磷酸鹽)中。濾紙過濾除去殘留脂肪和不溶性物質,濾液用雙縮脲法測蛋白濃度,熒光光度法測定濾液中二酪氨酸含量。測定條件為:發射波長420 nm(狹縫5 nm),激發波長325 nm(狹縫5 nm)。測定結果用熒光強度除以蛋白濃度,表示為相對熒光值。
蛋白質粒徑測定:采用Sun等[17]的方法,用馬爾文激光粒度儀3 000測定蛋白質粒度的分布,粒徑類型設定為非球形,密度為0.95 g/cm,分散劑為水。相對折射率和吸收率分別設定為1.54和0.001。獲得的參數如下:D4,3表示按體積計算的平均粒徑,Dx(10)、Dx(50)和Dx(90)分別表示占樣品10%、50%和90%的蛋白質所達到的最大粒徑。
蛋白質體外消化性測定:參考Wen等[18]的方法,略作修改。用33 mmol/L pH值1.8的甘氨酸緩沖液將蛋白液稀釋至4 mg/mL,取3 mL與6 mL胃蛋白酶(5 U/mg)混合,總共6份,在37℃下進行酶解90 min后用1 mL 15%的三氯乙酸終止反應,離心(6 000×g,7 min),用雙縮脲法定量其中3份剩余的不溶蛋白質量(m1,mg)。其余3份用33 mmol/L pH值8.0的甘氨酸緩沖液沖洗兩遍,然后用3 mL 33 mmol/L pH值8.0的甘氨酸溶解,加入1 mL胰蛋白酶(6.6 U/mg)和α-胰凝乳蛋白酶(0.33 U/mg)的混合液,在37℃下酶解60 min,立即加入1 mL 15%的三氯乙酸終止反應,然后離心(8 000×g,7 min),用雙縮脲法定量剩余的不溶蛋白質量(m2,mg)。用下式計算胃蛋白酶和胰蛋白酶對蛋白質的水解率。

1.3 數據分析
采用Excel 2010軟件對數據進行處理(計算平均值和標準偏差)及繪圖;采用SPSS Statistics 19.0軟件對數據進行顯著性和Pearson相關性分析,采用單因素方差分析 Duncan法比較風干牦牛肉加工不同時間點之間的差異,所有統計分析的顯著水平均為P<0.05。
2 結果與分析
2.1 脂質氧化變化
風干牦牛肉加工過程中POV和TBARS值變化如圖1所示。
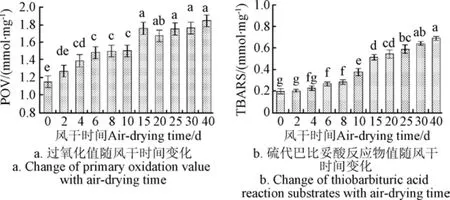
圖1 風干牦牛肉加工過程中脂質氧化變化Fig.1 Change of lipid oxidation in air-dried yak meat during processing
POV值反映了脂質氧化初期主要氧化產物氫過氧化物的含量,氫過氧化物是誘導蛋白質發生氧化的促進因子之一[7]。隨風干加工時間的延長,POV值呈現先增加后趨于穩定的趨勢(圖 1a),在第 15天時達最大值1.76 mmol/kg,15 d以后變化不顯著(P>0.05)。氫過氧化物不穩定,容易形成丙二醛等次級氧化產物,其含量用TBARS值表示。Mariutti等[19]認為氫過氧化物分解的主要途徑是2個氧原子之間的裂解,這2個氧原子分別產生一個烷氧基和一個羥基自由基,然后該烷氧基可以進一步通過β-裂解產生脂質二級氧化產物。TBARS值的變化如圖 1b所示,在整個風干加工過程中其值呈逐漸增加的趨勢,尤其在10 d以后顯著增加(P<0.05),最大值達到 0.69 mg/kg。Ripoll等[20]指出肉類變質的TBARS閾值水平為2~2.5 mg/kg,本試驗TBARS值未達到該范圍。
2.2 蛋白質氧化變化
羰基是蛋白發生氧化的標志性產物,羰基含量增加代表蛋白氧化程度的增加[6]。圖2中顯示了風干牦牛肉加工過程中羰基含量變化。
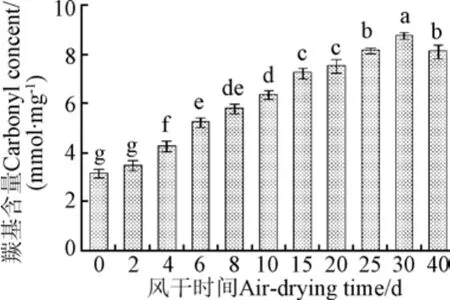
圖2 風干牦牛肉加工過程中羰基含量變化Fig.2 Change of carbonyl content in air-dried yak meat during processing
羰基化合物含量隨風干時間的延長呈逐漸上升趨勢。初始時為3.15 nmol/mg,至40 d時達到8.11 nmol/mg,增加了4.96 nmol/mg。肌肉蛋白質的氧化羰基化是一個不可逆的非酶促化學修飾[13],蛋白質側鏈羰基基團的形成主要來自賴氨酸、脯氨酸、精氨酸等的直接氧化,這類敏感性氨基酸先在自由基的攻擊下形成氨基自由基,氨基離子再通過水合反應形成氨基酸側鏈羰基衍生物[21]。因此,脂質氧化形成的氫過氧化物和丙二醛等自由基促進了風干牦牛肉蛋白質羰基化。
2.3 蛋白質交聯變化
圖 3反映了風干牦牛肉加工過程中總巰基、二硫鍵以及二聚酪氨酸含量的變化。
在整個風干加工過程中,總巰基含量顯著下降(P<0.05),和原料肉相比,至40 d時降低了31.03 nmol/mg(圖 3a)。Lund等[22]也報道了類似的研究,他發現豬肉在冷藏期間總巰基數量顯著減少。風干牦牛肉中二硫鍵含量(圖 3b)則呈顯著增加趨勢(P<0.05),肌肉蛋白質中半胱氨酸的巰基(-SH)容易被氧化形成二硫鍵(-S=S-),導致巰基含量下降和新的交聯蛋白分子產生[23]。酪氨酸易受活性氧自由基攻擊形成二聚酪氨酸,其含量可用相對熒光值來表示[16],二聚酪氨酸含量變化如圖 3c所示,在風干牦牛肉加工過程中呈逐漸增加趨勢,在0~30 d內變化顯著(P<0.05)。以上結果表明,風干牦牛肉加工過程中蛋白質氧化后形成了二硫鍵和二酪氨酸,使蛋白質分子發生交聯[10]。
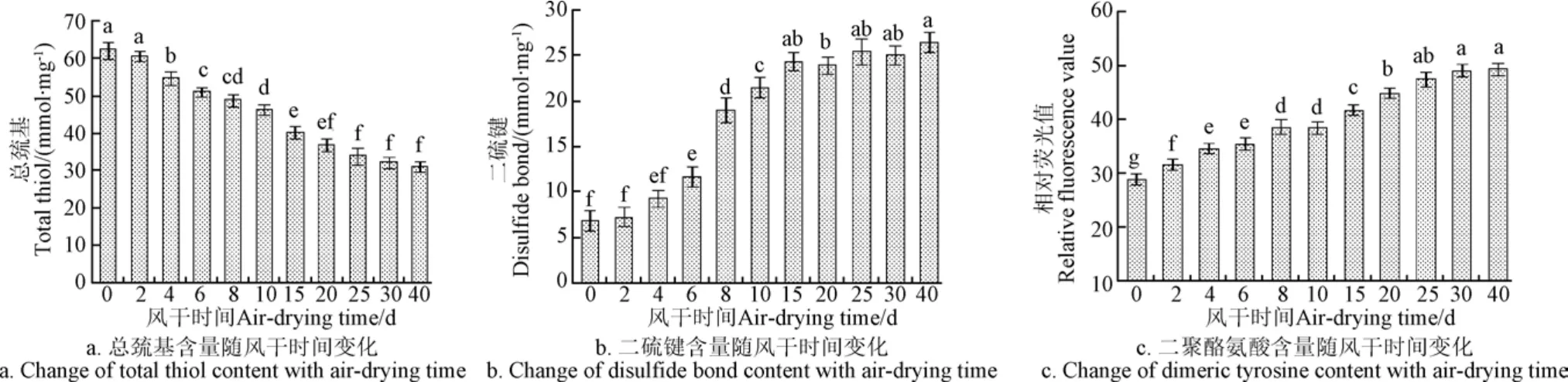
圖3 風干牦牛肉加工過程中蛋白質交聯變化Fig.3 Change of protein cross-linking in air-dried yak meat during processing
2.4 蛋白質粒徑大小分布變化
風干牦牛肉加工不同時間點肌原纖維蛋白粒徑分布變化如表1所示。
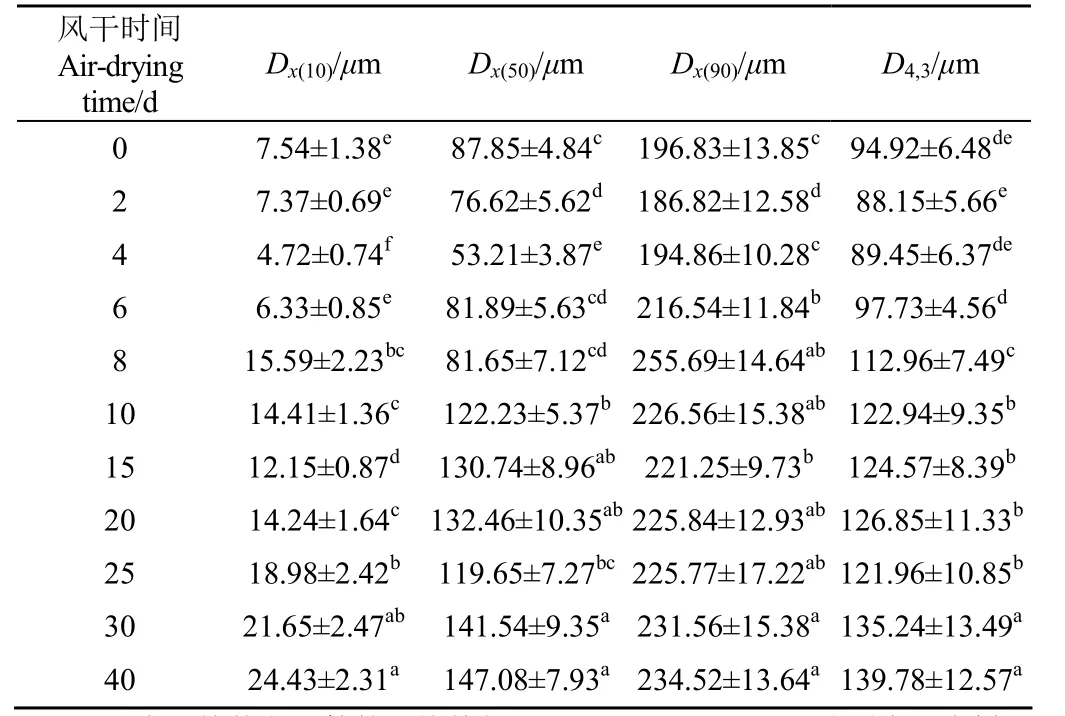
表1 風干牦牛肉加工過程中肌原纖維蛋白粒徑分布變化Table 1 Change of the myofibrillar protein particle size distribution in air-dried yak meat during processing
在風干加工最初的4 d內,Dx(10)、Dx(50)、Dx(90)和D4,3均顯著降低(P<0.05),在第4天時的值分別為4.72、53.21、194.86和89.45μm;在4~40 d內則顯著增加(P<0.05),至40 d時分別為24.43、147.08、234.52和139.78μm。Dx(10)、Dx(50)、Dx(90)和D4,3在第一階段的降低可能是由于牦牛肉中內源酶對蛋白質的水解作用,使高分子量蛋白質降解為小分子蛋白質或肽以及游離氨基酸[17]。在第 4天后,Dx(10)、Dx(50)、Dx(90)和D4,3均逐漸增加,表明牦牛肉風干加工過程中產生了大分子蛋白質。這一現象可能是肌原纖維蛋白分子間形成的二硫鍵或二聚酪氨酸交聯物導致小分子蛋白質結合生成了大分子蛋白質[24]。
2.5 蛋白質體外消化性變化
風干牦牛肉在加工過程中其肌原纖維蛋白體外消化率變化如圖 4所示,用胃蛋白酶對蛋白質的水解來模擬胃消化,胰蛋白酶和α-胰凝乳蛋白酶共同對蛋白質的水解來模擬腸消化過程。
隨著風干時間的延長,胃蛋白酶對肌原纖維蛋白的水解率逐漸降低(P<0.05),由初始的62.36%降低至40 d時的33.92%,40 d時顯著低于15 d(P<0.05),極顯著低于0~8 d(P<0.01)。胰蛋白酶對蛋白質的水解率則呈現逐漸增加趨勢(P<0.05),在加工初始時,胰蛋白酶消化率為4.78%,至40 d時升高至13.64%。Santé-Lhoutellier等[5]發現如果胃蛋白酶在早期對蛋白質的水解作用較弱時,則留給了胰腺蛋白酶足夠的反應底物,在后期胰蛋白酶和α-胰凝乳蛋白酶的降解作用會增強。然而胰蛋白酶和胃蛋白酶總水解率在整個加工過程中顯著降低(P<0.05),至40 d時降低至47.56%,降低了19.58%,40 d時顯著小于30 d(P<0.05),極顯著小于0~15 d(P<0.01)。可見蛋白質的消化性主要取決于胃蛋白酶的水解作用。
2.6 各指標間的相關性分析
風干牦牛肉在加工過程中,脂質氧化、蛋白氧化、蛋白粒徑以及蛋白消化率各指標間的 Pearson相關系數(r)和顯著水平如表2所示。
由表2可知,風干牦牛肉加工過程中,POV和TBARS與羰基、二硫鍵以及二聚酪氨酸呈極顯著的正相關關系(P<0.01),與巰基則呈極顯著的負相關關系(P<0.01)。Wang等[8]報道鯉魚肌纖維蛋白中的羰基化合物隨著丙二醛濃度的增加而增加[9]。Estévez[23]強調蛋白質與非蛋白質羰基化合物的共價結合是導致蛋白質羰基化的途徑之一。Stadtman等[25]指出,肉中某些敏感氨基酸(如賴氨酸,脯氨酸,精氨酸等)側鏈殘基被一類活性基團攻擊而直接氧化是產生羰基的主要原因之一,例如氫過氧化物。此外,丙二醛可與組氨酸殘基 N末端形成席夫堿,并與谷氨酰胺的親電性ε-氨基反應形成二氫吡啶[26],這些生化反應可使蛋白質產生羰基化合物。
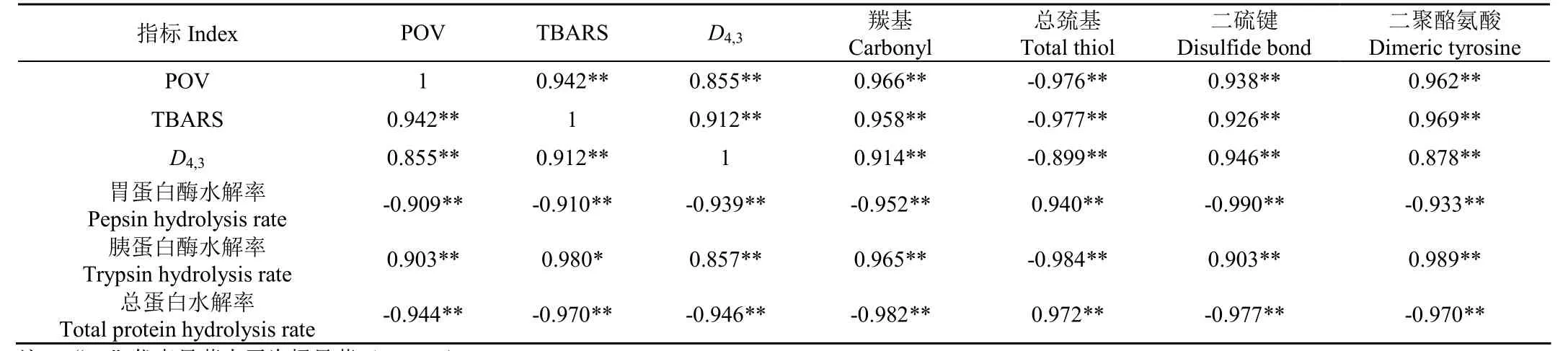
表2 各指標間的Pearson相關系數(r)Table 2 Pearson correlation coefficients (r) of various indexes
風干牦牛肉在加工過程中,二硫鍵和二聚酪氨酸的形成使蛋白質分子發生了交聯[10]。丙二醛本身是親核交聯劑[27],它可以與蛋白質中的 2個賴氨酸殘基反應形成雙席夫堿二亞胺交聯物,這可能導致蛋白質分子內或分子間的交聯[28]。一般認為自由基誘導的蛋白質氧化可發生二硫鍵和二聚酪氨酸的交聯。自由基可以從半胱氨酸的巰基中奪取氫原子,然后 2個巰基結合形成二硫鍵并交聯[10]。酪氨酸氧化后形成的二聚酪氨酸是肉類加工中另一種常見的交聯形式[22]。Zhang等[29]認為在自由基存在條件下,二聚酪氨酸的生成可以通過兩步催化氧化來實現,首先,酪氨酸的單電子氧化產生酪氨酸自由基,其次,2個單體酪氨酸自由基的組合形成二聚酪氨酸。總之,風干牦牛肉加工過程中脂質氧化促進了蛋白質氧化,導致蛋白質分子發生交聯。
總蛋白水解率與羰基、二硫鍵以及二聚酪氨酸呈現出極顯著的負相關關系(P<0.01),與總巰基變化則表現出極顯著的正相關關系(P<0.01)。D4,3與二硫鍵以及二聚酪氨酸表現出極顯著的正相關關系(P<0.01)。可見牦牛肉在自然風干過程中,其肌原纖維蛋白消化性隨蛋白氧化程度的增加而降低。根據Rysman等[30]的研究,蛋白質的氧化修飾使蛋白水解變得困難,因為α-酮酰基衍生物與N末端氨基酸的反應會阻止蛋白質水解酶對蛋白質的降解。Stadtman[25]認為肉類產品蛋白質中芳香族氨基酸(酪氨酸等)是被蛋白酶水解的切割位點之一,此類氨基酸發生氧化修飾會阻止蛋白質的降解。此外,Santé-Lhoutellier等[10]報道蛋白質氧化后發生分子內或分子間的交聯,如二硫鍵、二聚酪氨酸等的生成引起蛋白質聚集,最終導致蛋白質的消化率下降。因此,從蛋白質氧化角度考慮,風干牦牛肉加工過程中其蛋白質消化率下降的原因有2個方面,一是部分氨基酸側鏈被修飾,二是蛋白質交聯物的生成使蛋白質分子粒徑變大或聚集。以上變化均可引起蛋白水解酶與蛋白質的結合能力減弱,導致蛋白質水解率降低。總之,風干牦牛肉傳統的加工工藝耗時過長,導致其脂質、蛋白質氧化程度增加和蛋白質消化性降低。今后可以引進熱風干等加工技術以縮短加工時間;另外,在風干牦牛肉加工前先將牦牛肉采用抗壞血酸棕櫚酸酯、維生素E、茶多酚等可食性抗氧化劑以及一些具有抗氧化作用的果蔬汁進行腌制處理,以期有效降低脂質氧化和蛋白質氧化程度和極大限度保證產品營養品質。
4 結 論
1)傳統風干牦牛肉加工過程中,蛋白質消化性顯著降低(P<0.05),而脂質氧化程度顯著增加(P<0.05),并且蛋白質羰基化合物、二硫鍵及二聚酪氨酸交聯物含量均呈顯著增加趨勢(P<0.05)。
2)牦牛肉在自然風干過程中,蛋白質消化性與蛋白質羰基、二硫鍵、二聚酪氨酸以及脂質氧化產物之間呈極顯著負相關關系(P<0.01),脂質氧化似乎促進了蛋白質氧化進而導致肌原纖維蛋白消化率降低。
進一步要進行的工作是研究新的加工技術以縮短加工時間,或將可食性天然抗氧化劑以及一些具有抗氧化作用的果蔬汁應用于風干牦牛肉的加工中,以期降低脂質、蛋白質氧化程度和保證產品營養價值。