好氧反硝化細菌的鑒定及其脫氮特性研究
2020-08-07 01:24:08龔望寶王廣軍余德光李志斐田晶晶
水生生物學報 2020年4期
朱 云 龔望寶 謝 駿 王廣軍 余德光 李志斐 張 凱 田晶晶
(1.中國水產科學研究院珠江水產研究所,農業農村部熱帶亞熱帶水產資源利用與養殖重點實驗室,廣州 510380;2.上海海洋大學,水產與生命學院,上海 201306)
隨著我國水產養殖業的快速發展,養殖方式從傳統粗放型向現代規模化、集約化轉變[1],養殖過程中營養物質的高投入,導致池塘養殖內源氮素污染嚴重,不僅會惡化水質,致使養殖動物疾病暴發,養殖產品質量和產量下降,而且氮素超標嚴重的養殖尾水排放會導致江河水域環境的污染。因此,如何降低池塘養殖水體中氮素污染成為水產養殖領域的熱點問題。生物脫氮以無污染、高效率的優點被認為是去除水體中氮素的有效途徑之一[2]。傳統的生物脫氮理論認為反硝化是嚴格厭氧的過程[3],氧氣會抑制反硝化還原酶基因的表達和反硝化還原酶的活性,不能發揮微生物脫氮的作用。
20世紀80年代,Robertson和Kuenen[4]首次成功分離出了一株細菌Thiosphaera pantotropha(現命名為Paracoccussp.),報道了該菌能夠在好氧條件下進行反硝化作用,并將此現象命名為好氧反硝化。Bell等[5]對Paracoccussp.的反應機理開展了進一步研究,結果表明,好氧反硝化的發生是由于細胞內存在的好氧反硝化作用酶系。好氧反硝化細菌能夠在有氧的環境中將NO-N和NO-N還原為N2,這就為生物脫氮提供了嶄新的思路[6]。此后,越來越多的好氧反硝化細菌從不同的環境中分離出來,如糞產堿菌(Alcaligenes faecalis)[7]、假單胞菌(Pseudomonadaceae)[8]、不動桿菌(Acinetobacter)[9]、叢毛單胞菌(Comamonas)[10]、紅球菌(Rhodococcus)[11]、芽孢桿菌(Bacillus)[12]和副球菌(Paracoccus)[13]等,其中以假單胞菌屬最多[14—17]。好氧反硝化細菌的發現,為現有的養殖池塘水體中氮素的去除工藝提供了新的研究方向。
本研究從廣東省中山市草魚養殖池塘水體中分離出一株高效好氧反硝化細菌ZS1,對該菌株進行形態觀察、生理生化特性和16S rDNA基因序列分析,探究了該菌株在好氧條件下的脫氮特性以及不同環境因子對菌株ZS1脫氮效率的影響,以期為養殖池塘尾水處理系統的生物脫氮技術工藝提供理論支持及初步的候選菌株。
1 材料與方法
1.1 池塘條件及樣品采集
本實驗在廣東省中山市某養殖場進行,池塘面積約10000 m2,水深1.8 m。該池塘采用輪捕輪放的生態養殖技術,三年不換水,其中主養草魚密度為30000 尾/hm2,規格為1000 g/尾。
采樣時間為2018年3月20日,養殖水溫24.3℃,溶氧為4.5 mg/L,pH為7.7。上午10點用采水器5點采樣法取池塘水樣裝于無菌水樣袋中,置于4℃冷藏箱中,并立即帶回實驗室進行菌株的富集和分離。
1.2 培養基
(1)基礎富集培養基: 酵母浸膏5 g,蛋白胨10 g,NaCl 5 g,pH=7。
(2)反硝化富集培養基[18]: 酵母浸膏3.0 g,蛋白胨5.0 g,KNO3l.0 g,蒸餾水1000 mL,pH=7.0—7.6。
(3)溴百里酚藍(BTB)培養基: KNO31.0 g,C6H5Na3O21.0 g,KH2PO41.0 g,FeSO4·7H2O 0.05 g,CaCl20.2 g,MgSO4·7H2O 1.0 g,1%的溴百里香酚藍 1 mL,蒸餾水999 mL,pH=7.2。
(4)反硝化性能測定培養基: CH3COONa 2.56 g,KNO30.361 g,MgSO4·7H2O 0.2 g,KH2PO4l.0 g,K2HPO45.0 g,NaCl 0.5 g,微量元素溶液1 mL,蒸餾水999 mL,pH=7.4。
(5)微量元素溶液: EDTA 50 g,CaCl25.5 g,ZnSO42.2 g,MnCl2·4H2O 5.06 g,FeSO4·7H2O 5.0 g,(NH4)6MO7O2·4H2O 1.1 g,CuSO4·5H2O 1.57 g,CoCl2·6H2O 1.61 g,蒸餾水1000 mL,pH=7.0。
1.3 菌株的富集、分離和篩選
將養殖場采集的水體10 mL放入裝有90 mL反硝化細菌富集培養基的250 mL錐形瓶中,在搖床上培養(180 r/min,30℃)。24h后,取菌懸液體1 mL,放置到9 mL無菌水的離心管中,混合混勻,然后用涂布平板10倍稀釋法分離好氧反硝化細菌。將平板長出的菌落進行3 次純化,得到純菌。
挑取純化后的純菌株單菌落至液體反硝化富集培養基中至其長到對數期,將其稀釋為細菌濃度為4×108cfu/mL的菌懸液,取1 mL菌懸液接種到裝有100 mL液體培養基的250 mL 錐形瓶中,放置在搖床中培養(180 r/min,30℃),24h后測其NO-N和TN的去除效率,并選取NO-N去除效率在90%以上且總氮去除效率在60%以上的菌株開展進一步分析,共分離得到89株反硝化細菌,其中菌株ZS1脫氮性能最優,使用甘油法[19]將ZS1置于-80℃保存待用。
1.4 菌株的鑒定
從形態學、生理生化和16S rDNA三方面對菌株進行鑒定。
掃描電鏡(SEM)觀察: 取菌液體1.5 mL在1000 r/min 條件下離心10min,使用3%的戊二醛溶液固定24h后,通過掃描電鏡(JSM-5800,Japan JEOL)拍攝觀察菌株形態,在廣東省微生物分析檢測中心測定分析。
采用細菌基因組DNA試劑盒(Omega,USA)提取菌株ZS1 的DNA,然后以其為模板進行PCR擴增,選用細菌通用引物[20],上游引物為27F(5′-AGAG TTTGATCCTGGCTCAG-3′),下游引物為1492R(5′-TACGGCTACCTTGTTACGACTT-3′)。PCR擴增反應體系為: DNA模板1 μL,TaqDNA聚合酶25 μL,27F引物 (10 μmol/L) 1 μL,1492R引物 (10 μmol/L)1 μL,ddH2O(22 μL)。將PCR反應體系充分混合均勻后,在PCR擴增儀(Heal Force,T960,杭州)上進行PCR反應,反應條件為: 94℃預變性5min,94℃變性1min,56℃退火1min,72℃延伸2min,共30個循環,最后72℃延伸8min,4℃ forever。PCR擴增產物 (約1400 bp) 經1.5%瓊脂糖凝膠電泳(EB染色) 確定條帶后,送至上海美吉生物公司進行測序分析,將16S rDNA基因序列在GenBank核酸序列數據庫中進行同源性比較,以Neighbor-Joining法構建系統發育樹。
1.5 菌株ZS1好氧反硝化性能測定
將活化好的菌液按體積比1%接種到以KNO3為唯一氮源的反硝化培養基的錐形瓶中,培養基的初始pH為7.4,碳源為乙酸鈉,C/N為15,培養條件為溫度30℃,轉速180 r/min,設3組平行,置于恒溫振蕩培養箱中培養48h,每隔6h取樣監測菌株的生長、TN、NO-N、NO-N等指標,分析菌株ZS1的反硝化性能。
1.6 不同環境因子對菌株ZS1的脫氮影響
向裝有100 mL已滅菌的反硝化培養基的三角瓶中接入菌株ZS1,研究不同溫度(20、25、30、35、40℃)、C/N值(5、10、15、20、25)、初始pH(6、7、8、9、10、11)、搖床轉速(0、50、100、150、200 r/min)和碳源種類(檸檬酸鈉、乙酸鈉、丁二酸鈉、蔗糖、葡萄糖)對菌株ZS1反硝化特性的影響。以上處理的基本條件為: 溫度30℃、初始pH 7.4、C/N為15、初始懸浮液NO-N含量為50 mg/L、搖床轉速為180 r/min、碳源為乙酸鈉、KNO3為唯一氮源。然后將其中一種條件改變,其余反應條件與基本條件相同,培養24h后,測定培養基中菌株ZS1的生長、NO-N、NO-N和TN的濃度,并計算NO-N和TN的去除率。
1.7 分析方法
在本研究中,NO-N的測定采用N-(1-萘基)乙二胺光度法,NO-N的測定采用麝香草酚分光光度法,TN的測定采用堿性過硫酸鉀氧化紫外分光光度法,菌株的生長采用光度比濁法,用紫外可見分光光度計于600 nm處測量吸光度值(TU-1810,北京通用),pH采用pH計(pHB-3,上海)測定,所有樣品均測定3次,取平均值。脫氮率計算方法[21]如下:

式中,Q為氮的去除率,ct為終末氮濃度(mg/L),c0為初始氮濃度(mg/L),V為氮的去除速率[mg/(L·h)],t為時間。
本研究采用Excel、MEGA7.0和Origin8.5軟件對實驗數據進行統計分析與作圖。
2 結果
2.1 菌株的鑒定
形態特征菌株ZS1革蘭氏染色反應呈陰性,生理生化特性如表1所示: 接觸酶、氧化酶反應陽性;硝酸鹽還原和檸檬酸鹽利用陽性;七葉苷水解和明膠液化陽性;吲哚反應、V-P實驗、淀粉水解、酪素水解、脲酶、ONPG試驗、精氨酸雙水解酶均為陰性。在基礎富集培養基上培養48h后,觀察到菌株ZS1的菌落形態特征為: 圓形,呈灰白色,凸起,邊緣整齊。經掃描電鏡觀察,結果如圖1所示,菌體呈桿狀,長1.5—2.0 μm,寬0.7—0.9 μm,有鞭毛。
16S rDNA序列分析和系統發育分析菌株ZS1的16S rDNA基因序列長度為1333 bp,測序結果已上傳至GenBank數據庫,登錄號為MK615112。系統發育樹如圖2所示,菌株ZS1與Pseudomonas furukawaii同源性達100%。結合菌株的形態學、生理生化特性和16S rDNA基因序列分析結果,將菌株ZS1確定為Pseudomonas furukawaii。
2.2 菌株ZS1的好氧反硝化特性
在以KNO3為唯一氮源的情況下,觀察菌株ZS1在48h內的生長情況及好氧反硝化特性。如圖3所示,在6—18h,可以觀察到菌株ZS1的A600顯著增加,從0.133增加到0.930;培養18h后菌體生長變緩,在18—24h生長處于穩定期;培養24h后,菌體密度呈現減小趨勢,進入衰亡期。在0—24h階段,NO-N濃度迅速下降,從48.93 mg/L降至7.32 mg/L,NO-N去除率為85.04%,去除速率達1.734 mg/(L·h);TN的含量從52.04 mg/L 降低到13.73 mg/L,去除率為73.62%,去除速率達1.596 mg/(L·h)。24h后,菌株進入衰亡期,A600開始下降,NO-N和TN濃度也趨于穩定。培養至48h時,NO-N的含量降低到1.27 mg/L,去除率達到97.40%,去除速率為0.993 mg/(L·h);TN的含量繼續降低到8.40 mg/L,去除率達83.86%,去除速率達0.909 mg/(L·h),且無NO-N積累。

表 1 菌株ZS1的生理生化特性Tab.1 Physiological and biochemical characteristics of strain ZS1
2.3 不同環境因子對菌株ZS1反硝化性能的影響
不同碳源對菌株ZS1反硝化特性的影響如圖4所示,不同碳源對菌株ZS1的生長及脫氮率的影響存在很大差異。在5種不同碳源下,菌株的A600均顯著增多,由高到低的順序為乙酸鈉(0.930)、檸檬酸鈉(0.878)、葡萄糖(0.777)、丁二酸鈉(0.623)和蔗糖(0.013)。當菌株ZS1分別以5種碳源為唯一碳源時,對-N的去除率從高到低依次為乙酸鈉(99.47%)、檸檬酸鈉(99.39%)、葡萄糖(95.13%)、丁二酸鈉(75.49%)和蔗糖(26.20%);對TN的去除率從高到底依次為乙酸鈉(88.52%)、檸檬酸鈉(88.31%)、葡萄糖(78.12%)、丁二酸鈉(55.05%)和蔗糖(3.22%)。菌體生長情況與N和TN的去除率具有一致性,密度越大,對-N和TN的去除率就越高。當以蔗糖為碳源時,菌株ZS1幾乎不能生長,TN去除率僅為3.22%。因此,在實際池塘養殖尾水生物脫氮處理中可以將乙酸鈉、檸檬酸鈉和葡萄糖作為菌株ZS1的最適碳源。
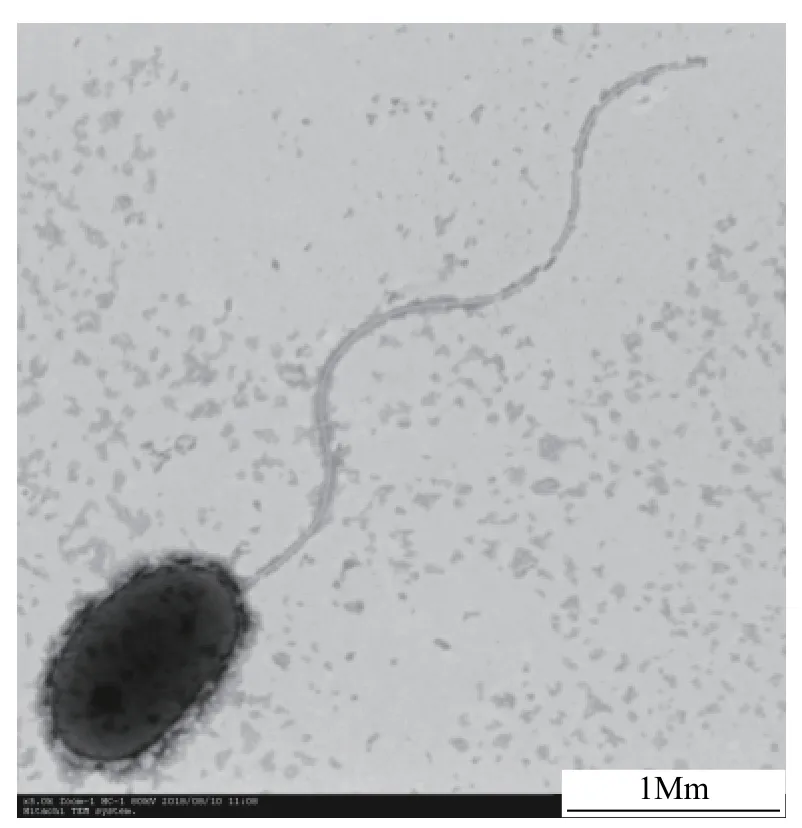
圖1 掃描電鏡下菌株ZS1的形態特征Fig.1 Morphological characteristics of strain ZS1 under a scanning electron microscope
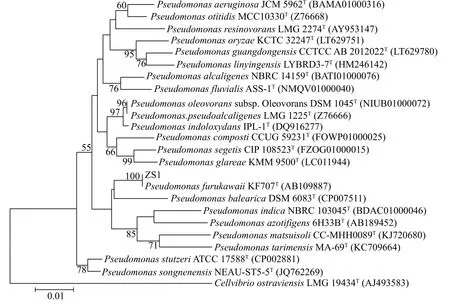
圖2 基于16S rDNA分析的菌株ZS1系統發育樹Fig.2 Phylogenetic tree of strain ZS1 based on 16S rDNA analysis
不同C/N對菌株ZS1反硝化特性的影響如圖5所示,菌株ZS1的A600隨著C/N的增加而升高。當C/N為5時,TN和NO-N去除率均達最低值,分別為19.74%和51.28%,NO-N積累達最高值,為0.11 mg/L;當C/N從5上升至15時,TN和NO-N去除率同步上升達到最高值,分別為88.31%和99.39%;當C/N從15上升至25時,TN和NO-N去除率基本保持不變。因此,菌株ZS1發揮反硝化功能的最適C/N為15—25。
不同pH對菌株ZS1反硝化特性的影響如圖6所示,當培養基的pH為6和11時,嚴重抑制菌株ZS1的生長,A600值僅為0.003和0.001,同時其TN和NO-N的去除率也均最低,分別為6.43%和10.16%。當pH在7—10時,菌株ZS1生長良好,A600在0.734—0.773,其對NO-N和TN的去除率較高,NO-N的去除率在99.34%—99.59%,TN的去除率在60.735%—73.77%。因此,菌株ZS1發揮反硝化功能的最適pH為7.0—10.0。
不同溫度對菌株ZS1反硝化特性的影響如圖7所示,當溫度為20℃時,菌株ZS1的A600為0.048,幾乎停滯生長。當溫度上升到25℃時,NO-N和TN去除率分別為98.42%和71.98%。當溫度為30℃時,NO-N和TN去除率達到最高,分別為99.18%和77.99%。當溫度繼續上升到40℃時,菌株的生長和
不同搖床轉速對菌株ZS1反硝化特性的影響培養基中的溶解氧濃度是通過改變搖床轉速來進行調控。如圖8所示,隨著搖床轉速的升高,菌株的去氮能力逐漸加強。當轉速為50 r/min時,NO-N和TN去除效率極低,分別僅有9.19%和5.03%;當轉速上升至100 r/min時,NO-N和TN的去除率顯著上升,分別達93.33%和78.72%;當轉速繼續上升至150 r/min時,NO-N和TN的去除率達最高值,分別為99.35%和83.09%;轉速升高到200 r/min時,NO-N和TN的去除率保持穩定。因此,菌株ZS1發揮反硝化功能的最適搖床轉速為100—200 r/min。

圖3 菌株ZS1好氧反硝化特征及生長曲線Fig.3 Aerobic denitrification performance and growth curve of the strain ZS1

圖4 碳源對菌株ZS1反硝化性能的影響Fig.4 Effect of the carbon source on the denitrification ability of strain ZS1
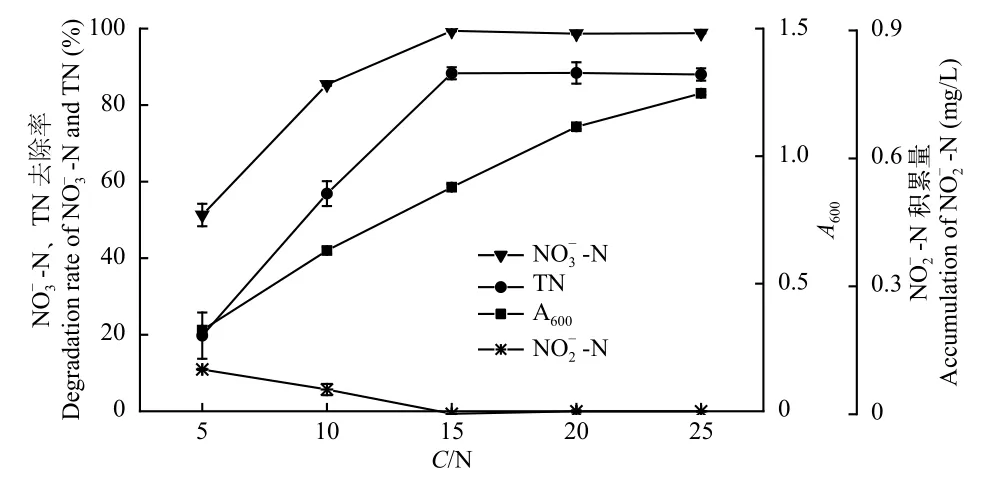
圖5 C/N對菌株ZS1反硝化性能的影響Fig.5 Effect of the C/N ratio on the denitrification ability of strain ZS1

圖6 pH對菌株ZS1反硝化性能的影響Fig.6 Effect of pH on the denitrification ability of strain ZS1
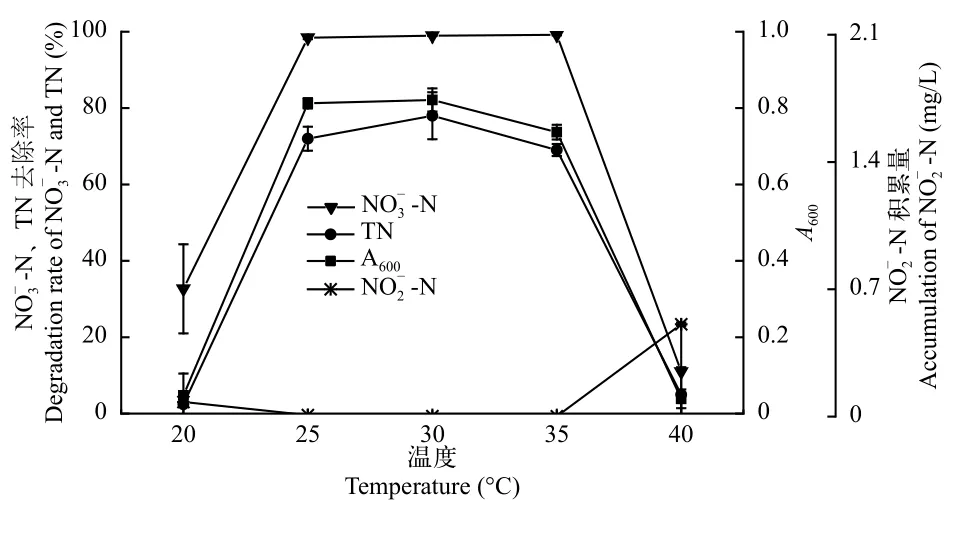
圖7 溫度對菌株ZS1反硝化性能的影響Fig.7 Effect of temperature on the denitrification ability of strain ZS1

圖8 轉速對菌株ZS1反硝化性能的影響Fig.8 Effect of rotation speed on the denitrification ability of strain ZS1
3 討論
3.1 不同碳源對菌株ZS1脫氮性能的影響
有機碳源在菌株的反硝化過程中提供電子和能量,不同碳源因其化學結構和分子量的差異而對菌株的生長和脫氮效果產生不同的影響[21]。在本研究中,當菌株ZS1以乙酸鈉、檸檬酸鈉和葡萄糖為碳源時,菌株生長良好,具有高效的脫氮性能,其NO-N的去除率分別為99.47%、99.39%和95.13%,TN的去除率分別為88.52%、88.31%和78.12%,菌株ZSI基本不能利用蔗糖,本實驗結果與修海峰等[8]、張婷月等[22]和蔡茜等[23]研究結果一致。顏薇芝等[9]發現不動桿菌YN3以葡萄糖和蔗糖為碳源時,菌株基本不增長,檸檬酸鈉是其最佳碳源,氨氮去除率達到99%。這說明相對于糖類,菌株更高效利用有機酸,是因為有機酸是三羧酸循環的中間產物,能被細菌直接利用,而糖類需要先轉化為有機酸再被利用。同時有機酸還能增強周質硝酸還原酶的活性[24]。反硝化反應屬于氧化還原反應,不同碳源對菌株的脫氮性能產生差異,也可能與不同碳源的氧化還原電位有關[25]。
3.2 不同C/N對菌株ZS1脫氮性能的影響
C/N是菌株能否進行完全反硝化的關鍵因素,直接影響細菌的生長和對有機物質的去除效果[23]。大多數研究結果表明,細菌的脫氮效率會隨著C/N的增大而升高,達到一定值后,脫氮效率維持穩定,繼續增大C/N,脫氮效率反而會呈下降趨勢[26—28]。趙丹等[29]發現假單胞菌ZD8在好氧條件下,C/N在4—22時,NO-N的去除率隨著C/N增加而增強,當C/N為22時,其NO-N去除率達到最高,為99.2%,
3.3 不同pH對菌株ZS1脫氮性能的影響
培養基的pH通過影響微生物細胞膜上的電荷,從而決定微生物體內營養物質的轉移方式,進而影響微生物對營養物質的吸收和轉化[32]。pH過高或過低都會導致微生物酶的活性降低或喪失,從而降低菌體的反硝化活性[33]。菌株只有在最適pH下才具有快速生長和高效脫氮的能力[34]。在本實驗中,pH為7—10時,菌株ZS1的脫氮性能最佳,NO-N和TN的去除率分別在99.34%—99.59%和60.73%—73.77%。目前已報道的好氧反硝化細菌中大部分菌株的最適pH為中性或偏堿性,如張淑楠等[35]從地熱水中分離出的一株芽孢桿菌,生長和高效反硝化作用的適宜pH為7—8。黃廷林等[36]篩選出的貧營養菌株A14在pH為7時,對NO-N的去除效果最佳。高喜燕等[37]研究發現菌株在 pH為7.5時的脫氮率最高,但在過酸(pH<5.5) 或過堿(pH>9) 的條件下,菌株幾乎不生長,脫氮性能也最低。
3.4 不同溫度對菌株ZS1脫氮性能的影響
溫度是影響細菌反硝化性能的重要因素之一,主要是通過影響微生物細胞結構從而影響其生長和新陳代謝[38]。許多研究證明,好氧反硝化細菌的最適溫度為20—35℃,如Ye等[39]報道了菌株Providencia rettgeri strainYL的最適脫氮溫度為25℃;修海峰等[8]報道德克斯氏菌株DF2的最適脫氮溫度為30℃;菌株Pseudomonas stutzeriF1在溫度低于20℃和高于35℃時,其生長和脫氮能力均受到抑制[40]。本實驗中菌株ZS1發揮好氧反硝化性能的最適溫度為25—35℃,與上述報道一致。
3.5 不同搖床轉速對菌株ZS1脫氮性能的影響
本實驗通過轉速來調節培養基中的溶解氧,結果表明,轉速過低導致培養基中的溶氧偏低,抑制了菌株ZS1的生長和脫氮性能,當轉速在100—200 r/min時,菌株ZS1的生長和脫氮性能顯著增加。王田野等[28]發現不動桿菌SQ2脫氮性能最佳的轉速為180—220 r/min。易立等[26]報道蠟樣芽胞桿菌CZ1在轉速為160 r/min時,其NO-N去除率達99.29%。菌株Acinetobacter calcoaceticusN7在轉速為230 r/min時,TN去除率達到最大,為89.8%[27]。本實驗結果與上述報道一致,隨著搖床轉速的提高,培養基中的液體形成渦流,延長了空氣的停留時間,加大了空氣接觸面積,使培養基中的溶解氧提高,滿足了菌株的生長代謝,提高了脫氮過程中相關酶的合成能力,導致菌株的脫氮性能增強。因此,在實際生物脫氮處理過程中,要控制適當的溶解氧,以適合菌株的生長,從而提高菌株去氮的效率。
本研究中P. furukawaiiZS1具有高效的好氧反硝化性能,下一步需要深入開展該菌株在實際養殖水體中的應用效果以及該菌株好氧反硝化的分子機制,為池塘養殖尾水處理應用生物脫氮技術提供理論依據。
4 結論
(1)本實驗從中山市主養草魚池塘水體中分離篩選出1株具有高效好氧反硝化能力的菌株,經形態觀察、生理生化特性以及16S rDNA序列分析,鑒定為Pseudomonas furukawaii,命名為P. furuka-waiiZS1。(2)經48h培養后,NO-N的含量從48.93降至1.27 mg/L,去除率為97.40%,去除速率達0.993 mg/(L·h);TN的含量從52.04 mg/L 降低到8.40 mg/L,去除率為83.86%,去除速率達0.909 mg/(L·h),且無NO-N積累。以NO-N為唯一氮源時,菌株ZS1實現最佳好氧反硝化性能的碳源為乙酸鈉、檸檬酸鈉和葡萄糖,溫度為25—35℃,pH為7.0—10.0,C/N為15—25,轉速為100—200 r/min。(3)在好氧條件下,P. furukawaiiZS1具有高效脫氮效率,該菌株為池塘養殖尾水生物脫氮處理提供初步的候選菌株。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
