重口裂腹魚的胚胎發育觀察
2020-08-28 11:34:30胡仁云張運海舒旗林羅武蔣澤民丁勇冉玉鳳譚德清
江蘇農業科學 2020年14期
胡仁云 張運海 舒旗林 羅武 蔣澤民 丁勇 冉玉鳳 譚德清
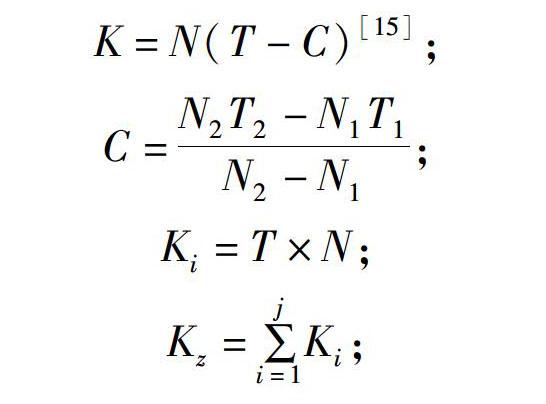
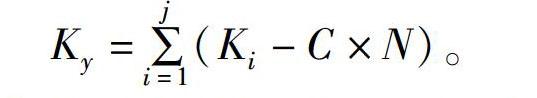

摘要:2017年、2018年對采自毛爾蓋水電站魚類增殖站內的重口裂腹魚(Schizothorax davidi)進行人工繁殖,并對其胚胎發育特征進行觀察。結果顯示,重口裂腹魚魚卵為黃色、微黏性,卵徑為(2.91±0.29) mm,魚卵吸水后直徑為(3.99±0.31) mm;在水溫為(14.5±12) ℃的條件下經217 h 20 min孵化出膜,有效積溫為665.27 ℃·h,胚胎發育生物學零度為(1130±0.06) ℃,初孵仔魚全長為(11.00±0.18) mm。
關鍵詞:重口裂腹魚;胚胎發育;有效積溫;生物學零度;發育觀察
中圖分類號: S917.4;S961.2? 文獻標志碼: A
文章編號:1002-1302(2020)14-0198-05
重口裂腹魚(Schizothorax davidi),別稱重唇魚、重口、細甲魚、雅魚、丙穴魚、嘉魚等,屬鯉形目(Cypriniformes)、鯉科(Cyprinidae)、裂腹魚亞科(Schizothoracinae)、裂腹魚屬(Schizothorax),常生活在流水或急流水的中下層,是以浮游動植物、水生昆蟲幼蟲為主食的雜食性魚類,主要分布于岷江、嘉陵江、烏江、漢江等中、上游,有時冬季也可在長江干流的中、下游發現[1-3]。近年,受水電站建設、垃圾污染、過度利用等因素的影響,總體數量銳減,生存條件日趨惡化,野生重口裂腹魚資源遭到嚴重破壞[4-6]。因此,提高重口裂腹魚的人工繁殖技術,以培育魚苗實施放流活動,對補充和恢復重口裂腹魚野生資源至關重要。目前,有關該魚的研究在早期資源調查、形態發育、細胞組織學等方面已有較多報道[7-11];對重口裂腹魚仔魚發育、眼早期形態發育、鱗片發育及覆蓋過程等方面有部分報道[12],但僅限于重口裂腹魚胚后發育研究,在胚胎發育方面尚未見報道。本研究對重口裂腹魚胚胎發育過程進行較全面的觀察,以期豐富重口裂腹魚生物學研究資料。
1 材料與方法
1.1 試驗材料
本次試驗的材料來源于毛爾蓋水電站魚類增殖站內重口裂腹魚的受精卵。2017年、2018年分別對毛爾蓋水電站魚類增殖站內的重口裂腹魚親本進行人工繁殖,具體做法是在水溫為14 ℃左右時,對發育成熟的親本注射催產劑,待效應時間到達時,分別對雌魚及雄魚采卵和采精,通過干法人工授精獲得受精卵。受精卵均勻分布于用聚乙烯網片制成的孵化框內孵化,網片為40目,尺寸為 40 cm×60 cm×30 cm。
1.2 試驗方法
將孵化框置于循環水養殖缸內,孵化期間每2 h記錄1次水溫,采用循環水自動控溫設備將孵化水溫控制在(14.5±1.2) ℃。胚胎發育觀察時,每次使用小撈網隨機撈20粒卵,使用解剖鏡觀察發育進程并拍照記錄,同時記錄時間、發育特征等,其中胚胎發育時序按60%受精卵發育至同一時期記時。囊胚期之前每30 min觀察1次,原腸胚至神經胚期每2 h觀察1次,以后每5 h觀察1次,直至出膜。胚胎發育分期參考已發表的小裂腹魚和短須裂腹魚胚胎發育等相關的報道[13-14]。
1.3 數據處理
胚胎發育圖片采用Photoshop CS6編輯,試驗數據采用Excel、SPSS Statistics等軟件進行統計分析。積溫根據有效積溫法計算,即
2 結果與分析
2.1 胚胎發育
2.1.1 成熟卵及卵裂階段
由圖1-A至圖1-J可知,重口裂腹魚的魚卵為黃色,也有部分卵為灰色,魚卵直徑為(2.91±0.29) mm,受精卵吸水1 h后卵徑達最大,最大直徑為(3.99±0.31) mm,卵周隙為(0.55±0.09) mm;卵裂階段持續時間為12 h 47 min,分為8個時期,主要特征為顯微鏡下可數細胞增多,形狀可見,分裂溝清晰。
2.1.2 囊胚期階段
由圖1-K至圖1-M可知,囊胚期持續時間為8 h 53 min,分3個時期,囊胚由高變低,開始沿卵黃邊緣向下分裂延展,植物極基本在囊胚早期形成。
2.1.3 原腸胚階段
由圖1-N至圖1-P可知,原腸胚階段持續時間為24 h 18 min,分3個時期,胚層下包卵黃囊的1/2~3/4高度,體積逐漸增大,厚度減小,且厚度出現差異,其中一側集中加厚隆起分化形成胚盾,原腸晚期時初步形成大卵黃栓。
2.1.4 神經胚階段
由圖1-Q至圖1-S可知,神經胚階段持續時間為9 h 6 min,分3個時期,胚體持續分化,并下包至卵黃囊的4/5高度,神經原基逐漸分化為神經管,末期時胚孔封閉,胚胎相位變化結束,為器官系統發育作準備。
2.1.5 器官系統發育階段
器官系統發育階段持續時間為108 h 18 min,分13個時期。孵化55 h 46 min 時肌節出現4~8對,初現腦空泡(圖1-T);孵化67 h 6 min時頭部有1條模糊的眼基線出現,肌節7~10對;孵化84 h 45 min時卵黃囊開始內凹,由球形變為腎形,較細的一端出現1個小芽,進而中腦隆起,肌節9~13對;孵化98 h 20 min時胚體背部出現微弱的不規律的肌肉收縮4~8次/min,肌節28~30對,進而耳囊中央出現1對透光性差的小黑點,即為耳石(圖1-U至圖1-Z);孵化108 h 50 min時,在下頜與卵黃膜之間出現短管狀的心臟原基,胚體尾部拉長,卵黃囊變小,眼晶體變圓,肌節32~37對,再過約10 h吻端與大眼之間形成左右各1個凸起的嗅囊;孵化127 h 10 min后,圍心腔逐步增大,血細胞不明顯,心臟搏動頻率為 35 bpm(beat per minute),眼球黑色素沉積明顯(圖1-AA至圖1-AD)。
2.1.6 出膜期
出膜前胚體劇烈翻滾擺動,卵膜變薄,尾部分化得更長。出膜時,尾部附近的卵膜先破裂,尾部游離到水里并不停地擺動,經5~10 min后,軀干、頭部與卵膜分離,此時卵黃囊明顯變小,心率為65~75 bpm,后期卵膜逐漸溶解消失,孵化結束(圖1-AE至圖1-AF)。出膜期到卵膜破裂之間形態上基本沒有發生變化,在運動能力和力度方面增強,部分出現眼色素。60%魚苗出膜總歷時217 h 18 min。初孵仔魚在解剖鏡下呈淡黃色、半透明,心率為65~75次/min,心室、靜脈竇清晰,心臟內紅細胞明顯,頭部及脊椎結構清晰可見,各時期發育時序見表1。
2.1.7 積溫和生物學零度
試驗在溶氧為(8.5±0.98) mg/L,pH值為7.6±0.28,水溫為(14.5±12) ℃,流速為(5.0±1.5) cm/s的條件下進行。總歷時217 h 20 min,總積溫達到3 121.14 ℃·h時,60%的重口裂腹魚魚苗出膜。其中,卵裂階段所需積溫為200.7 ℃·h,占總積溫的6.4%,分化速度快、相位明顯;囊胚階段所需積溫為134.7 ℃·h,占總積溫的4.3%,囊胚期間細胞由清晰變模糊,由大變小,由少變多;原腸階段所需積溫為358.3 ℃·h,占總積溫的11.5%;神經胚階段所需積溫為 134.9 ℃·h,占總積溫的4.3%;器官系統發育階段所需積溫為1 581.6 ℃·h,占總積溫的50.7%,出膜階段所需積溫為708.2 ℃·h,占總積溫的22.7%。
通過2017年、2018年共3個批次的胚胎發育數據計算出重口裂腹魚胚胎發育的生物學零度(C)為(11.30±0.06) ℃,有效積溫Ky為665.27 ℃·h,相關數據見表2。
3 討論
3.1 重口裂腹魚胚胎發育特點
重口裂腹魚卵為微黏的沉性卵,其胚胎發育進程與大多裂腹魚類相似,但存在以下差異,詳見表3。
(1)重口裂腹魚成熟卵平均直徑為(2.91±0.29) mm, 遠大于塔里木裂腹魚、 小裂腹魚和伊犁裂腹魚的卵徑,大于短須裂腹魚、光唇裂腹魚、細鱗裂腹魚、異齒裂腹魚的卵徑,與四川裂腹魚、齊口裂腹魚、松潘裸鯉、拉薩裂腹魚、拉薩裸裂尻及尖裸鯉的卵徑接近。有研究表明,大卵黃可為仔魚提供更多的營養物質,延長內外營養源轉化時間,以提高野外生存的成活率[24],同時卵的大小直接影響卵裂速度、胚胎正常發育及出膜仔魚的大小[25-26],這也驗證了重口裂腹魚出膜仔魚全長較其他裂腹魚略大的結果。
(2)重口裂腹魚胚胎發育積溫遠高于小裂腹魚、細鱗裂腹魚、伊利裂腹魚、異齒裂腹魚、拉薩裂腹魚、尖裸鯉,而低于短須裂腹魚,與拉薩裸裂尻魚相當;由于各學者計算積溫的方法不同或者胚胎是否在最適溫度下試驗,致使積溫存在較大差異且可比性較低,相比之下生物學零度較有比較意義,本試驗中重口裂腹魚的生物學零度略低于四川裂腹魚和齊口裂腹魚,其他品種的有待研究。
(3)重口裂腹魚胚胎發育過程中耳石先于心臟原基出現,與短須裂腹魚心臟原基出現于耳石之前[14]不同, 與四川裂腹魚[16]相似;耳囊出現于尾芽之前與松潘裸鯉的耳囊出現于尾芽之后[23]不同;耳石在尾芽之后形成與拉薩裸裂尻魚的耳石先于尾芽形成[22]不同,而與拉薩裂腹魚[22]相似;肌肉效應時間比四川裂腹魚早[16]。
3.2 溫度對重口裂腹魚胚胎發育的影響
3個批次胚胎發育平均溫度略有差距,隨溫度升高,破膜時間縮短,說明提高孵化水溫可使魚苗提前出膜,這對規模化生產有重要意義,但不影響孵化率和成活率的最適孵化水溫有待進一步研究。
綜上,四川裂腹魚的產卵場所是水流較緩的河灘[16];松潘裸鯉為流水急流水的中下層魚類,產卵于近岸河灘礫石[3,27];短須裂腹魚于每年的12月至次年3月在流水淺灘處產卵[28];重口裂腹魚生存在峽谷急流河段,每年8—9月在水流較急的礫石河底產卵[2];由于重口裂腹魚產卵的季節、水流速度、底質和孵化環境存在較大差異,推測重口裂腹魚與其他裂腹魚類之間胚胎發育的差異可能與其更好地適應生存環境有關,具體與何種環境因子有關還須要進一步探索。
參考文獻:
[1]葉妙榮,傅天佑. 四川大渡河的魚類資源[J]. 資源開發與保護,1987,3(2):37-40.
[2]丁瑞華. 四川魚類志[M]. 成都:四川科學技術出版社,1994:379-381.
[3]鄧其祥,李 操,吳光舉. 岷江上游的魚類[J]. 四川師范學院學報(自然科學版),2001,22(1):21-25.
[4]范繼輝,劉 巧,麻澤龍,等. 岷江上游水電開發對環境的影響[J]. 四川環境,2006,25(1):23-27.
[5]徐 迅. 毛爾蓋水電站建設生態環境影響研究[D]. 雅安:四川農業大學,2013.
[6]李 明,傅 斌,王玉寬,等. 岷江上游水電開發特點及其空間格局分析[J]. 長江流域資源與環境,2015,24(1):74-80.
[7]楊世勇,陳道春,楊 淞,等. 重口裂腹魚精子的超微結構研究[J]. 安徽農業科學,2010,38(34):19504-19505,19508.
[8]彭 淇,吳 彬,陳 斌,等. 野生重口裂腹魚的性腺發育觀察與人工繁殖研究[J]. 海洋與湖沼,2013,44(3):651-655.
[9]寧 毅. 重口裂腹魚幼魚日糧中蛋白質、脂肪和碳水化合物需求量研究[D]. 南寧:廣西大學,2013.
[10]宋旭燕,吉小盼,楊玖賢. 基于棲息地模擬的重口裂腹魚繁殖期適宜生態流量分析[J]. 四川環境,2014,33(6):27-31.
[11]李忠利,陳永祥,胡思玉,等. 四川裂腹魚和重口裂腹魚形態差異的多元分析[J]. 動物學雜志,2015,50(4):547-554.
[12]嚴太明,周翠萍,李忠利,等. 實驗室飼養的重口裂腹魚仔魚的形態發育與生長吵[J]. 四川農業大學學報,2007,25(4):493-497,封3.
[13]冷 云,徐偉毅,劉躍天,等. 小裂腹魚胚胎發育的觀察[J]. 水利漁業,2006,26(1):32-33.
[14]左鵬翔,李光華,冷 云,等. 短須裂腹魚胚胎與仔魚早期發育特性研究[J]. 水生態學雜志,2015,36(3):77-82.
[15]孫儒泳. 動物生態學原理[M]. 北京:北京師范大學出版社,1987.
[16]陳永祥,羅泉笙. 四川裂腹魚繁殖生態生物學研究(續)Ⅱ胚胎發育的研究[J]. 畢節師專學報,1994(4):1-7.
[17]龔小玲,崔忠凱,吳敏芝,等. 塔里木裂腹魚胚胎和仔魚的發育與生長[J]. 上海海洋大學學報,2013,22(6):827-834.
[18]申安華,李光華,趙樹海,等. 光唇裂腹魚胚胎發育與仔魚早期發育的研究[J]. 水生態學雜志,2013,34(6):76-80.
[19]吳 青,王 強,蔡禮明,等. 齊口裂腹魚的胚胎發育和仔魚的早期發育[J]. 大連水產學院學報,2004,19(3):218-221.
[20]陳禮強,吳 青,鄭曙明,等. 細鱗裂腹魚胚胎和卵黃囊仔魚的發育[J]. 中國水產科學,2008,15(6):927-934.
[21]蔡林鋼,牛建功,張北平,等. 伊犁裂腹魚胚胎及早期仔魚發育的觀察[J]. 淡水漁業,2011,41(5):74-79.
[22]許 靜. 雅魯藏布江四種特有裂腹魚類早期發育的研究[D]. 武漢:華中農業大學,2011.
[23]吳 青,王 強,蔡禮明,等. 松潘裸鯉的胚胎發育和胚后仔魚發育[J]. 西南農業大學學報,2001,23(3):276-279.
[24]Knutsen G M,Tilseth S.Growth,development,and feeding success of atlantic cod larvae Gadus morhua related to egg size[J]. Trans Am Fish Soci,1985,114:507-511.
[25]Hansen T K,Falk-Petersen I B. The influence of rearing temperature on early development and growth of spotted wolffish Anarhichas minor (Olafsen)[J]. Aquaculture Research,2001,32(5):369-378.
[26]Inger-Britt F,Tove K H. Early ontogeny of the spotted wolffish (Anarhichas minor Olafsen)[J]. Aquaculture Research,2003,34(12):1059-1067.
[27]周仰躁,董存有,周學思,等. 四川九寨溝水生生物初步調查[J]. 四川動物,1986,3(2):15-17.
[28]顏文斌. 短須裂腹魚繁殖行為生態學研究[D]. 上海:上海海洋大學,2016.
收稿日期:2019-08-22
作者簡介:胡仁云(1991—),男,云南巧家人,水生動物執業獸醫,主要從事魚類人工繁殖研究。E-mail:1192354947@qq.com。
通信作者:譚德清,博士,副研究員,主要從事魚類營養與生態研究。E-mail:dqtan@ihb.ac.cn。
