浙江省紅楠生境地群落數量分類和環境解析
2020-09-17 00:01:18袁位高吳初平王志高朱錦茹黃玉潔
生態學報 2020年15期
任 晴,袁位高,吳初平,王志高,江 波,朱錦茹,黃玉潔,葉 明,方 慶
1 浙江農林大學 林業與生物技術學院,杭州 311300 2 浙江省林業科學研究院,杭州 310023 3 壽昌林場,建德 311600 4 建德林場,建德 311600
紅楠為樟科潤楠屬的常綠闊葉喬木,是亞熱帶和暖溫帶地區常綠闊葉林的主要建群種或者伴生種,常散生于山地闊葉混交林中[1-2]。紅楠不僅被廣泛用作綠化樹種和用材樹種,因其十分復雜的樹體化學成分,還多用于香料提取、藥品制備等領域[3-5]。但由于紅楠天然資源及其生態環境的嚴重破壞,使該植物逐漸成為漸危樹種[6]。國內對紅楠群落的研究主要集中在局部區域的群落調查。黃宗安[7]和曾繁茂[8]調查發現福建省尤溪縣境內的紅楠種群呈隨機分布,發展穩定。陳子林等[9]發現紅楠種群在浙江省六十田自然保護區呈聚集分布狀態。廖承川等[10]發現浙江九龍山國家級自然保護區紅楠群落以樟科和山茶科植物為主,并且群落中紅楠將長期處于優勢地位。但是以上研究僅直觀的反映局部區域紅楠的生境與分布狀況,并不能歸納出紅楠的典型生境和解釋環境因子對紅楠種群分布及發展的影響。另外紅楠林是浙江省典型森林類型,研究紅楠林環境與結構是天然林恢復、珍貴樹種等林業工程建設中急需解決的科學問題,對指導林業生態建設具有重要意義。
植被群落的數量分類和排序是揭示群落與環境關系的重要工具,雙向指示種分析(TWINSPAN)和典范對應分析(CCA)是目前國際上常用的研究方法[11-12]。楊國棟等[13]通過TWINSPAN 分類和DCCA排序研究了四川省寶興縣東拉山短絲木犀(Osmanthusserrulatus)植物群落的演替變化趨勢;商天其等[14]采用TWINSPAN 分類和CCA方法對嵊州市公益林群落進行數量分類與排序,發現以馬尾松(Pinusmassoniana)為建群種的群落是該區域主要的優勢群落類型;李婷婷等[15]對東北闊葉紅松林區的CCA排序結果顯示其變化格局主要受溫度和降水的影響。目前關于紅楠群落的排序研究鮮有報道。
探究群落內部優勢種之間的關系,對揭示群落結構特征及反映群落的穩定程度具有重要意義[16-17]。梁林峰等[18]對山西省文冠果(Xanthocerassorbifolia)群落優勢種的種間關聯性進行定量研究,發現群落整體結構不穩定,優勢物種處于演替變化過程中;俞筱押等[19]對貴州茂蘭國家級自然保護區內瀕危植物四藥門花(Loropetalumsubcordatum)群落各層次優勢種群的種間聯結性進行了分析,發現四藥門花在該群落中表現出較強的獨立性;高浩杰等[20]對舟山桃花島次生林優勢種紅楠的種內和種間競爭強度進行定量分析,發現其受到的種內和種間競爭壓力均衡。但是紅楠與群落物種間的關聯關系未見研究。
總體來說,目前關于紅楠群落的研究多集中于局部區域的種群分布和群落結構方面,而群落類型、群落分布與環境關系、群落內部種間關系等研究較為缺乏,群落結構、環境因子對紅楠分布及生長的影響尚不能解釋。本研究從覆蓋浙江省11個轄區下屬大部分縣級行政區的3181個監測樣地中,僅篩選出172個有紅楠分布的樣地,說明樣地環境、群落結構等對紅楠在浙江省的分布及生長有重要影響。因此本文運用雙向指示種分類(TWINSPAN)、典范對應分析(CCA)并結合種間聯結,對浙江省172個紅楠生境地群落進行分析,探討群落類型、環境因子、種間關系對浙江省紅楠分布及生長的影響,以期在省級層次上更為全面的為紅楠天然資源的恢復和發展提供科學依據。
1 研究區概況
浙江省境內地形起伏較大,西南、西北部地區群山峻嶺,中部、東南地區以丘陵和盆地為主,東北地區地勢較低,以平原為主。浙江省屬亞熱帶季風氣候,年平均氣溫15—18℃,光照較多,年平均日照時數1710—2100 h,降雨豐沛,年平均雨量980—2000 mm,空氣濕潤,氣候資源配制多樣。浙江省物種組成豐富,植被類型主要為常綠闊葉林、針葉林、針闊混交林、落葉闊葉林、常綠落葉闊葉混交林等[1-2]。
2 研究方法
2.1 樣地設置與調查
2009—2016年間設置的覆蓋浙江全省范圍的3181個監測樣地,每5年調查1次,本文采用最近1次(2013—2016年)調查數據,篩選出有紅楠分布的樣地共172個(表1、圖1)。樣地面積400 m2(20 m×20 m),調查內容包括胸徑≥5 cm個體的胸徑、高度、冠幅等物種數據,海拔、坡度、坡向、土層厚度等環境數據。氣候數據通過樣地所在縣氣象站獲取。
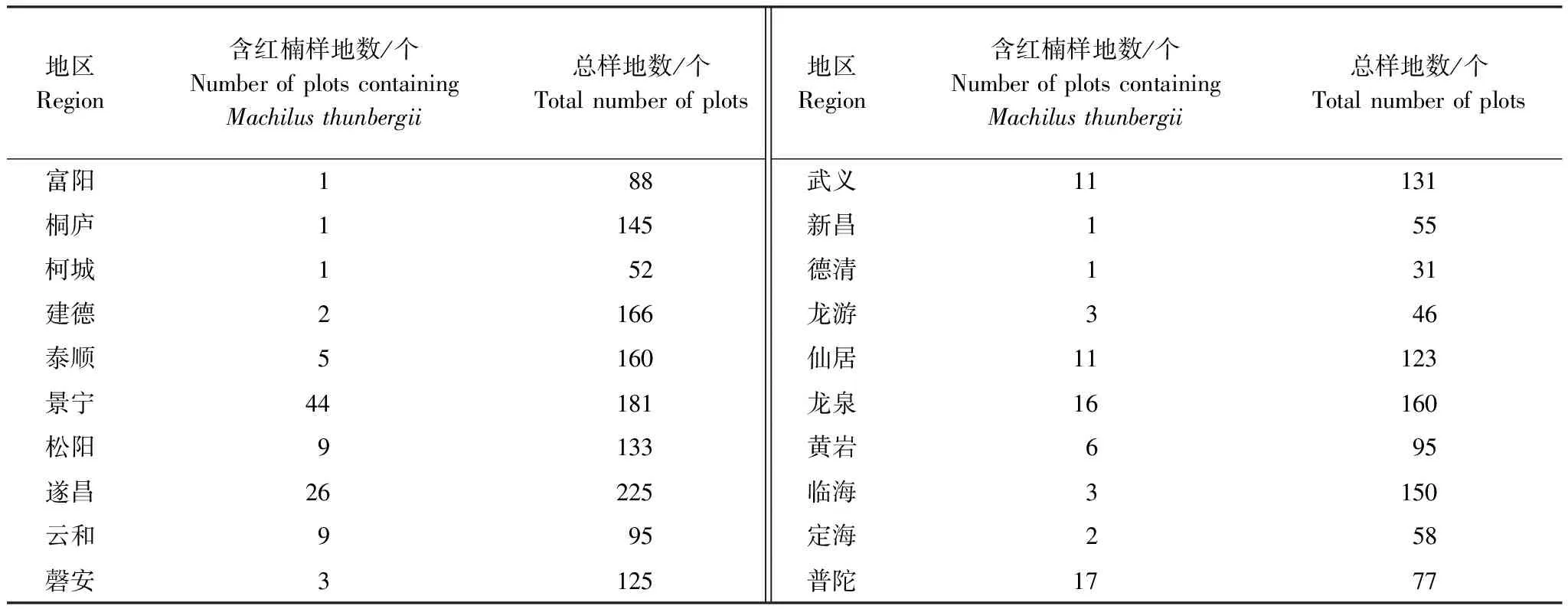
表1 浙江省含紅楠樣地的分布
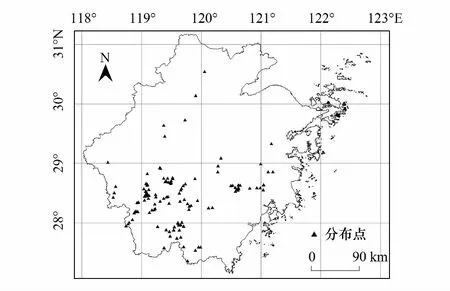
圖1 調查樣地分布圖
紅楠分布的172個樣地大部分集中于浙西南丘陵山地區域。樣地各環境因子的主要特征如表2所示,84.3%的紅楠分布在坡度為17°—35°的范圍內,95.35%的紅楠分布在海拔1000 m以下的范圍內。另外枯落物厚度5 cm以下、腐殖質厚度5—10 cm、土壤厚度30—50 cm的紅壤條件下有較多分布。
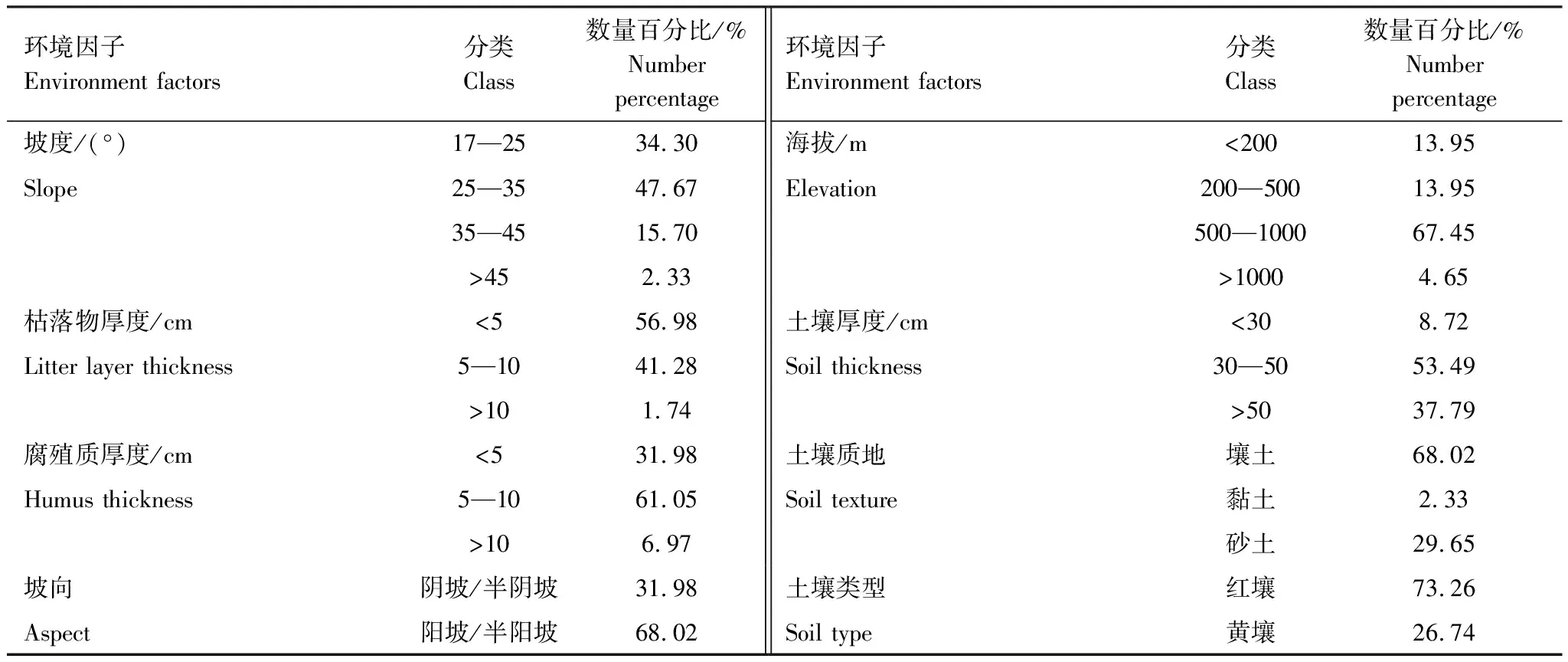
表2 紅楠分布區各環境因子的總體特征
2.2 數據處理
2.2.1重要值
重要值指標計算在Exccel中完成。計算公式為[21-22]:
相對多度=某一植物種類株數/所有植物種類株數×100%;
相對顯著度=某一樹種的胸徑平方和/所有樹種的胸徑平方和×100%;
相對頻度=某一樹種的頻度/所有樹種的頻度×100%;
喬木重要值=(相對顯著度+相對頻度+相對多度)/3。
2.2.2雙向指示種分析TWINSPAN
分類軟件采用WinTWINS 2.3,依據每個樣地所有物種重要值數據,整理成物種編號為橫坐標,樣地號為縱坐標的矩陣數據[23]進行載入分析。
2.2.3典范對應分析CCA
采用國際通用軟件CANOCO 4.5完成CCA排序。CCA要求兩個數據矩陣,一個是環境數據矩陣,一個是物種數據矩陣[23-24]。本文選取年均氣溫、年均降水量、坡度、坡向、海拔高度、土壤類型、土壤質地、土壤厚度、腐殖質厚度、枯落物厚度建立環境因子矩陣,坡向、土壤類型、土壤質地以數字等級表示,其他環境指標以實際觀測值表示[13-15]。植被數據矩陣為P×N維的物種重要值矩陣,其中P為植物的種數,N為樣地數。
2.2.4種間聯結
根據物種在樣方內是否出現的二元數據矩陣作為原始數據,以卡方檢驗(2)為基礎,結合聯結系數(AC)和共同出現百分率(PC)對群落優勢種進行種間聯結分析[17-19]。在計算2及聯結系數AC時,為避免a、c、d為0,導致2及AC出現無法比較關聯程度的情況,把a、c、d為0的數值都加權為1。由于本研究的取樣方法為非連續性取樣,使用非連續性數據的Yates連續校正公式計算2值。
式中n為取樣總數。當ad>bc時, 兩物種為正關聯, 存在較弱的競爭;當ad
聯結系數(AC)計算公式為:

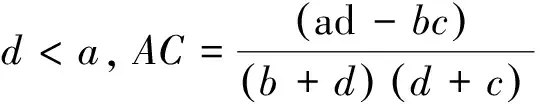
AC的值域為[-1, 1],其值越接近1,種對間的正聯結性越強;其值越接近-1,物種間的負聯結性越強;其值越接近0,種對間的聯結性越弱;當AC值為0,種對間完全獨立。
共同出現百分率(PC)計算公式為:
PC的值域為[0, 1], 其值越接近1,種對之間的正聯結關系越強;其值越接近0, 該種對的正聯結關系越弱。
3 結果與分析
3.1 群落物種組成與重要值
172個樣地中記錄到54科120屬167種植物,其中喬木119種,灌木或小喬木48種。物種組成以殼斗科、樟科、薔薇科、桑科、大戟科等科的植物為主(圖2),其中尤以殼斗科(18種)和樟科(18種)植物最為顯著。對整理好的172個樣地所有物種數據統一進行重要值計算,重要值大于1%的共有16個物種(表3)。
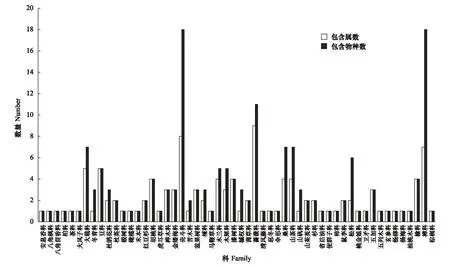
圖2 物種科屬分布圖
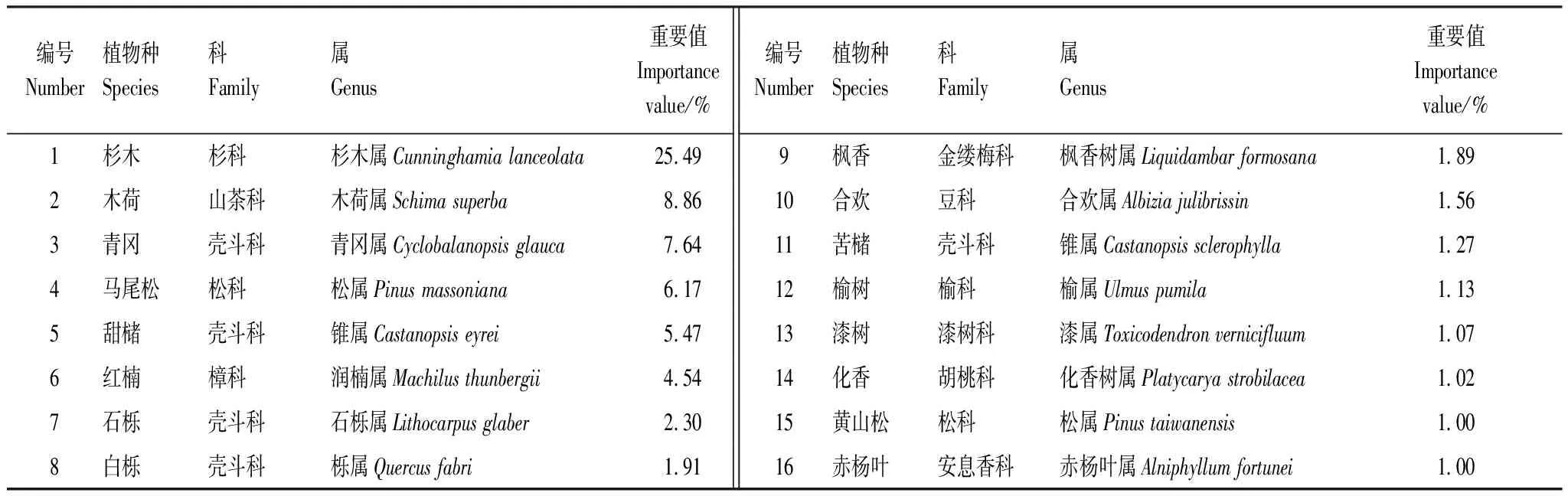
表3 16個優勢種列表
3.2 TWINSPAN分類結果與群落類型
根據TWINSPAN的分類結果(圖3),采取第5級水平的劃分,結合生態學意義,得到根據不同優勢種確定的8種群落類型:
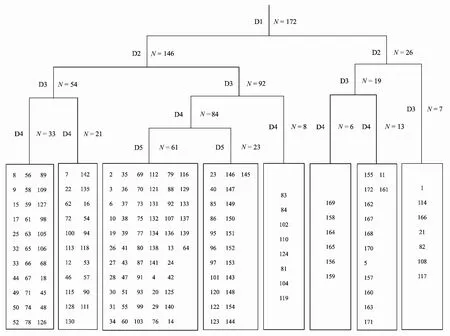
圖3 浙江省172個紅楠生境地群落的雙向指示種分類樹狀圖
Ⅰ為杉木+紅楠+木荷林,共計33個樣方,伴生種有青岡、甜櫧、苦櫧、白櫟、楓香、漆樹、合歡等。該群落分布于年均氣溫17.2℃,年均降水量1673 mm的地區,坡度22°—38°的陽坡或半陽坡,海拔高度350—950 m,平均海拔676.2 m,土壤為紅壤或黃壤。
Ⅱ為杉木林,共計21個樣方,伴生種有紅楠、木荷、甜櫧、石櫟、赤楊葉、漆樹、三角槭(Acerbuergerianum)等。該群落分布于年均氣溫17.3℃,年均降水量約1593 mm的地區,坡度25°—44°的半陽坡,海拔高度300—900 m,平均海拔616.62 m,土壤為紅壤。
Ⅲ為青岡+甜櫧+木荷林,共計61個樣方,伴生種有紅楠、石櫟、烏岡櫟(Quercusphillyraeoides)、冬青(Ilexchinensis)、杜英(Elaeocarpusdecipiens)、赤楊葉、漆樹等。該群落分布于年均氣溫17.2℃,年均降水量約1622 mm的地區,坡度25°—47°的半陽坡或半陰坡,海拔高度400—1300m,平均海拔731.8m,土壤為紅壤或黃壤。
Ⅳ為馬尾松+木荷林,共計23個樣方,伴生種有青岡、甜櫧、苦櫧、紅楠、黃山松、烏岡櫟、楓香、石櫟等。該群落樣地分布于年均氣溫17.4℃,年均降水量1570 mm的地區,坡度25°—42°的陽坡或半陽坡,海拔高度100—800 m,平均海拔440.1 m,土壤類型為紅壤。
Ⅴ為甜櫧+紅楠+青岡林,共計8個樣方,伴生樹種主要有石櫟、香樟(Cinnamomumcamphora)、木荷、山礬(Symplocossumuntia)、三角槭、白櫟。該群落分布于年均氣溫17.1℃,年均降水量約1544 mm的地區,坡度25°—45°的陰坡或半陽坡,海拔高度550—1250 m,平均海拔831.1 m,土壤為砂質黃壤。
Ⅵ為青岡+楓香林,共計6個樣方,伴生種有合歡、白櫟、紅楠、冬青、化香、黃檀(Dalbergiahupeana)、山礬、木荷等。該群落分布于年均氣溫16.1℃,年均降水量約1228 mm的地區,坡度17°—25°的半陰坡或半陽坡,海拔高度45—135 m,平均海拔70.2 m,土壤為紅壤。
Ⅶ為紅楠+榆樹林,共計13個樣方,伴生種有香樟、黃檀、化香、野桐(Mallotusjaponicus)、臭椿(Ailanthusaltissima)、冬青、山礬等。該群落分布于年均氣溫16.1℃,年均降水量約1242 mm的地區,坡度17°—33°的半陰坡或半陽坡,海拔高度24—220 m,平均海拔87.1 m,土壤為紅壤。
Ⅷ為紅楠+白櫟林,共計7個樣方,伴生種有苦櫧、黃檀、化香、鹽膚木(Rhuschinensis)、山胡椒(Linderaglauca)等。該群落分布于年均氣溫16.4℃,年均降水量約1489 mm的地區,坡度22°—35°的陰坡或半陽坡,海拔高度75—800 m,平均海拔437m,土壤類型為紅壤。
3.3 CCA排序
運用Canoco 4.5軟件對紅楠生境地的環境數據和所有物種數據進行排序。DCA排序結果表明,所有軸的梯度長度最大為4.214,應選擇單峰模型(CCA)。4個排序軸的特征值分別為0.513、0.377、0.239、0.188,前3個排序軸特征值占總特征值的85.7%,包含了絕大部分信息,表明本研究選取的環境因子對群落分布具有較好的解釋意義。
CCA排序(表4)結果說明環境因子對群落中物種分布具有顯著影響(P=0.002)。物種CCA排序軸第一軸特征值為0.274。物種-環境相關性系數為0.759。前四軸物種-環境關系方差累計貢獻率為89.8%,其中前兩軸就達到76.4%,說明CCA前兩軸已能較好地反映出研究區群落樣地、物種與其環境之間的關系。根據10個環境因子與CCA排序軸的相關性分析(表5)可知,海拔高度與CCA第1軸極顯著負相關,與CCA第1軸相關性最強,相關系數為-0.7608,其次是坡度,相關系數為-0.6405,與CCA第1軸呈顯著負相關關系。第二軸與土壤類型的相關系數最高。
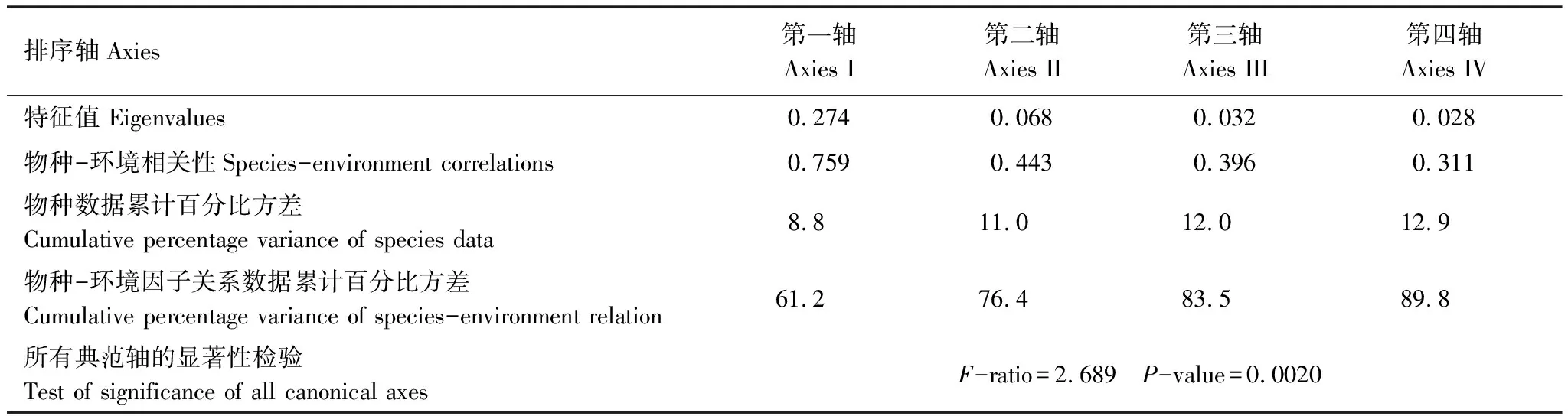
表4 CCA排序軸的特征值和累積百分比
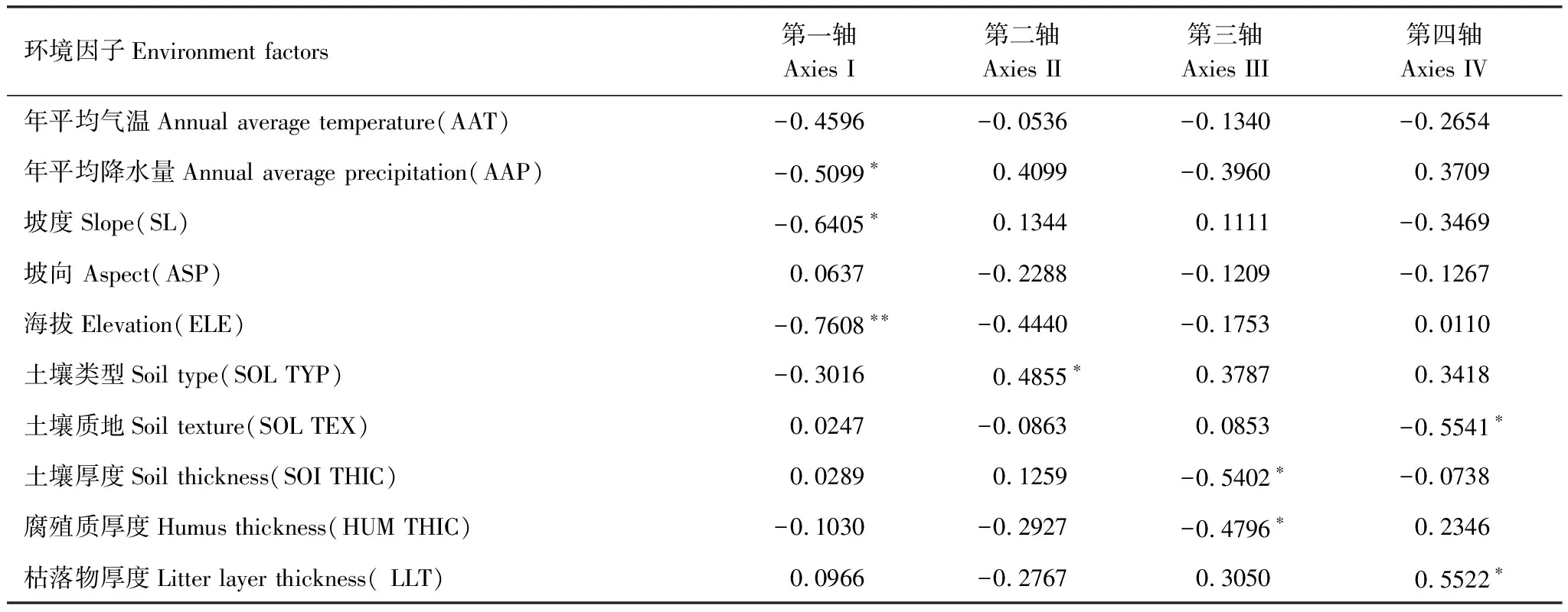
表5 環境因子與排序軸的相關關系
3.3.1樣地與環境因子排序
通過CanoDRAW分別對樣地、優勢種與環境因子關系進行分析繪制(圖4、圖5)。由圖4可知,第一軸反映了紅楠生境地群落所在生境的海拔、坡度及年平均降水量的情況,即沿著CCA第一軸從左往右,海拔、坡度逐漸降低,光熱條件逐漸變好,年均降水逐漸減少,海拔為主要影響因子。第二軸基本上表現了群落所在生境土壤類型的變化趨勢,即沿著CCA第二軸從上往下群落物種對紅壤的傾向減小,對黃壤的傾向增加,但紅壤類型下樣地數更多,群落多樣性更加顯著,表明浙江省內有紅楠分布樣地土壤多為紅壤類型。圖中8個群落類型未能有清晰地界線區分,存在明顯的聚集和交錯現象,這可能是由于樣地內物種組成和環境條件的相似性導致的。
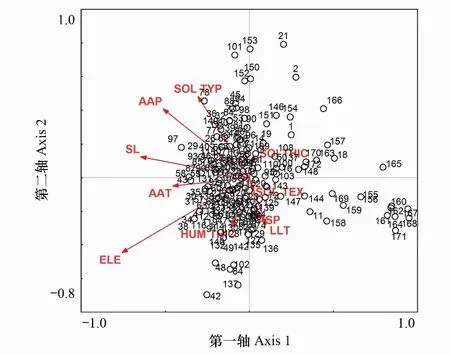
圖4 樣地與環境因子排序圖
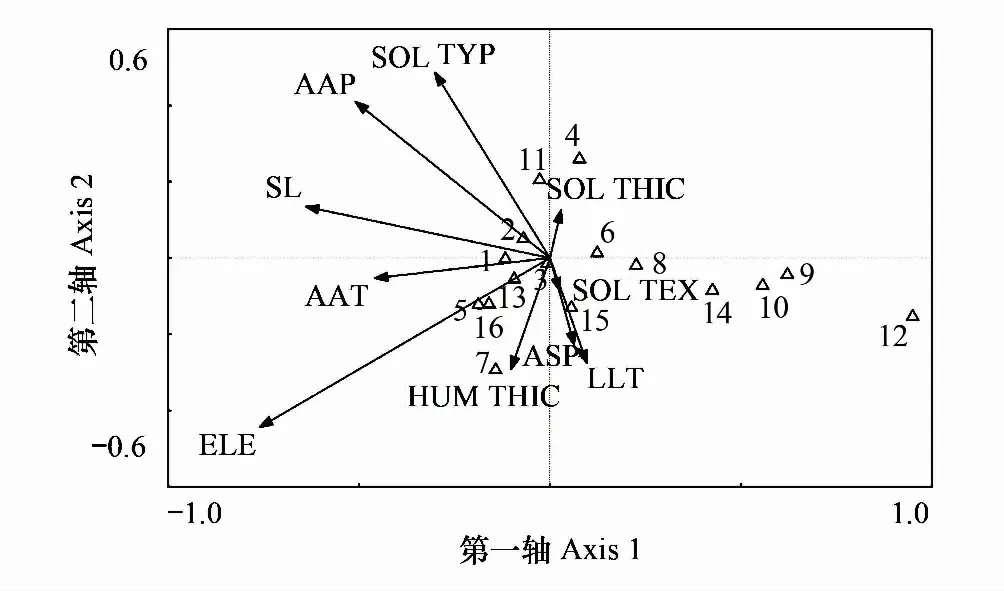
圖5 優勢種與環境因子排序圖
3.3.2優勢種與環境因子排序
由圖5可知,榆樹、楓香、合歡分布于第一軸右端,其生境為坡度平緩、光照條件較好、年均降水量相對較少的低海拔區域;而第一軸的左端并沒有物種的傾向性分布,說明16種優勢樹種并不適生于高海拔地區,紅楠與白櫟第一軸位置接近,說明二者對海拔、坡度要求相近。土壤類型與第二軸相關性最高,沿CCA的第二軸由下往上對土壤要求從黃壤向紅壤轉變。紅楠與木荷、杉木、白櫟在第二軸位置接近,說明這些物種對土壤要求相近。
3.4 優勢種種間關系
種間聯結能夠更直觀地反映種群之間相互關系, 多物種間總體關聯性在一定程度上反映了群落的穩定性[14]。
聯結系數AC結果(表6、圖6)中,共39個正聯結種對,34個負聯結種對,正負關聯比為1.1∶1,無關聯種對有47個。紅楠與甜櫧的AC值結果為極顯著正關聯,與木荷、青岡為顯著正關聯,與杉木為不顯著正關聯,與黃山松為不顯著負關聯;黃山松-合歡、榆樹-苦櫧、化香-苦櫧、赤楊葉-白櫟的AC值結果為極顯著負關聯。
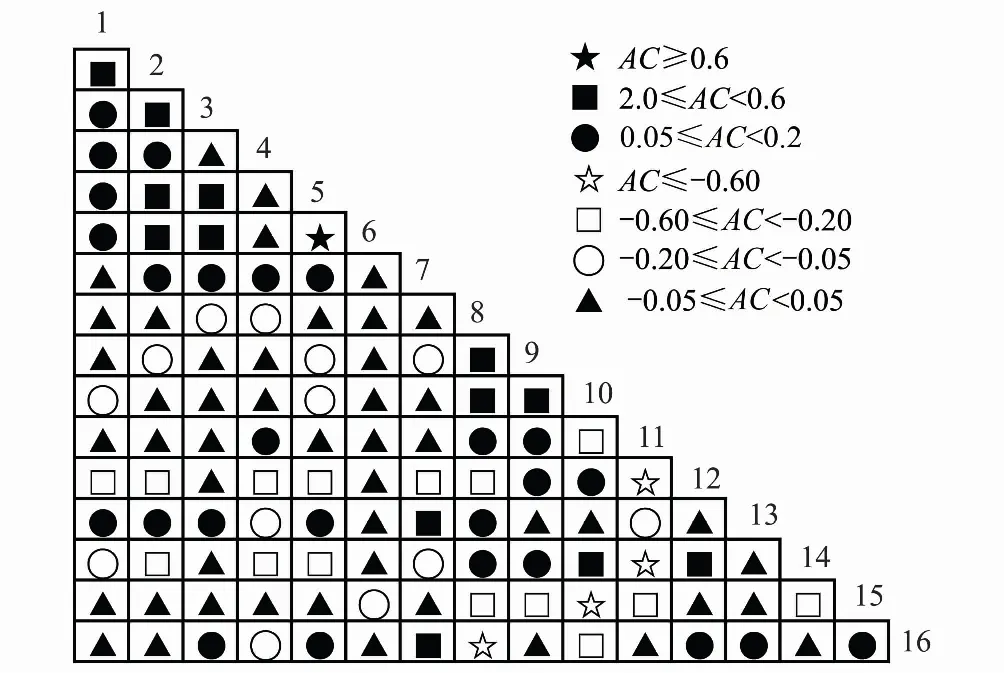
圖6 優勢種種間關聯的AC半矩陣圖
共同出現率PC結果(表6、圖7)中顯著率(包括極顯著)為8.3%,不顯著正關聯的有50個種對。無關聯的有60個種對,占50%。由此可見,主要優勢種間的正聯結程度不高。紅楠-杉木、紅楠-青岡、紅楠-木荷、杉木-木荷、木荷-青岡顯示為極顯著正關聯。顯著正關聯的種對分別是紅楠-甜櫧、杉木-甜櫧、青岡-甜櫧、木荷-甜櫧、杉木-青岡。
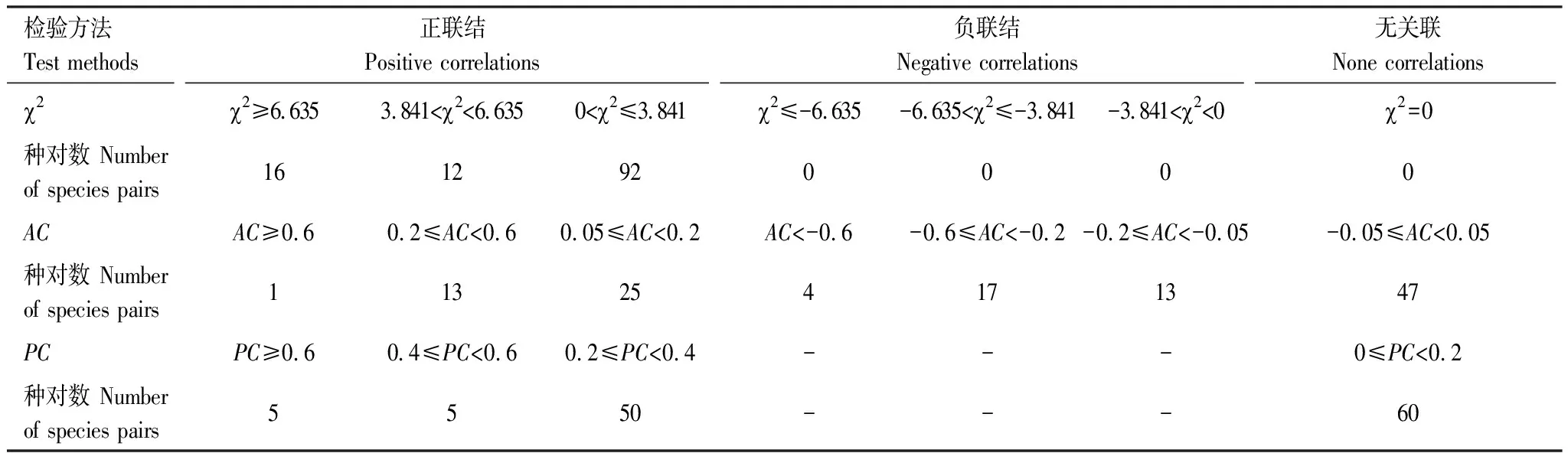
表6 16個優勢物種的卡方檢驗、聯結系數(AC)和共同出現百分率(PC)
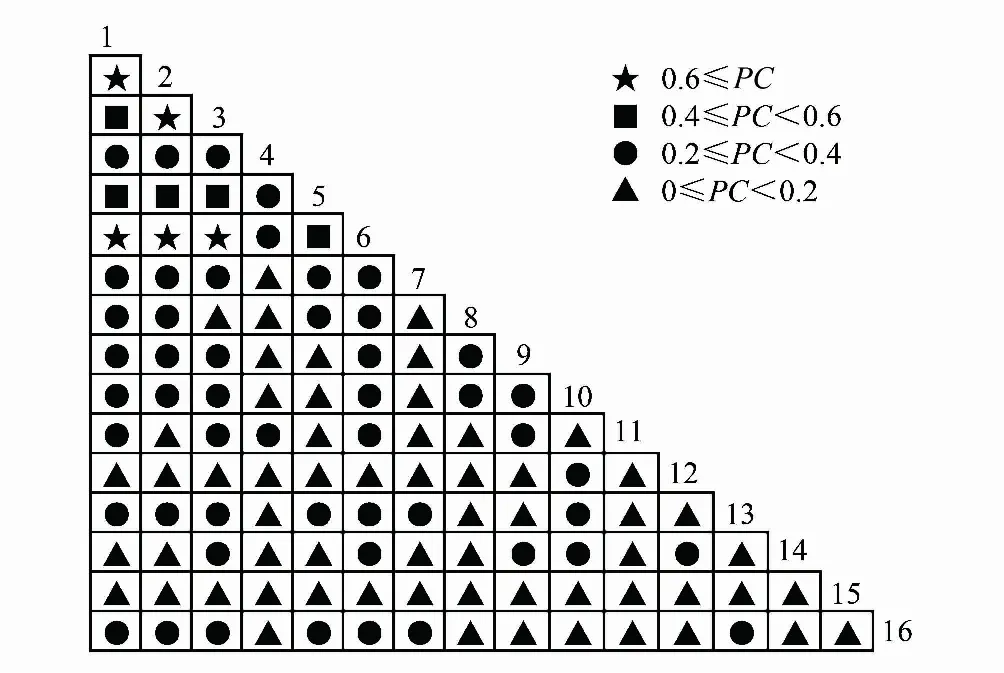
圖7 優勢種種間關聯的PC半矩陣圖
4 討論
本研究利用TWINSPAN等級分類,將浙江省內172個紅楠生境地群落劃分為8個類型,分別歸屬于浙江省四種典型的植被類型:針闊混交林、杉木林、常綠闊葉林、常綠落葉闊葉混交林。從演替角度來看,不同群落在環境因子影響下可能會向特定的方向演替發展。因此,環境狀況不同的區域可以指示不同的植被群落類型[15]。而隨著演替進展導致的群落內部環境改變(如光照)也有可能是影響紅楠分布及發展的主要原因[25]。
針闊混交林包括杉木、紅楠、木荷同為共建種的群落Ⅰ和馬尾松、木荷為共建種的群落Ⅳ,分布在坡度42°以下、海拔1000 m以下的低山區域。速生強陽性樹種杉木、馬尾松和喜光樹種木荷占據林冠層,與同層弱陽性樹種紅楠對光照資源需求互補。群落Ⅰ優勢種種間聯結緊密,紅楠、杉木、木荷、青岡、甜櫧之間均有顯著關聯,紅楠在群落中為優勢種,處于林分上層,群落組成較穩定。群落Ⅳ中紅楠與木荷、青岡、甜櫧有顯著正關聯,與馬尾松無顯著關聯,與黃山松為不顯著負關聯,群落穩定性有待提高。兩類群落分別由杉木林和馬尾松林演替而來,正處于向常綠闊葉林發展的過渡階段,群落物種組成豐富,已儲存了常綠闊葉林的大部分物種,種群結構穩定,更新良好。
群落Ⅱ屬于杉木林,樣地集中于坡度37°以下、海拔900 m以下的低山區域。這一群落以杉木為主,物種豐富度低,仍屬于演替初級階段,林分整體質量較差,通過調整杉木密度,加快下層樹種進入喬木層,進行杉木林的闊葉化改造,可以提高群落穩定性。隨演替進展,耐陰樹種紅楠優勢度會逐漸增大,此時應注意紅楠種群密度并疏伐上層高大喬木,增加林窗或林緣,增強紅楠與林下陽性植物的競爭力,提高紅楠種群的更新能力。
常綠闊葉林包括以青岡、甜櫧、木荷為建群種的群落Ⅲ和以甜櫧、紅楠、青岡為建群種的群落Ⅴ,多分布在坡度39°以下、海拔1300 m以下的中低山區域。紅楠處于林分中上層,與優勢種青岡、甜櫧、木荷均有顯著正關聯。兩類群落喬木層樹種主要集中在木荷屬、栲屬、櫟屬,僅有少量的白櫟、赤楊葉等落葉樹種間雜在林分上層。雖然林分中下層多樣性較豐富,但由于常綠闊葉林郁閉度較高,林下光照較弱,容易出現林分斷層現象,應該適當疏伐上層高大喬木,促進群落結構優化,保證紅楠幼苗的補充連續性。
闊葉混交林包括青岡、楓香為共建種的群落Ⅵ,紅楠、榆樹同為建群種的群落Ⅶ,紅楠、白櫟同為建群種的群落Ⅷ。群落Ⅵ和群落Ⅶ均分布于浙東南沿海島嶼坡度較緩、溫暖避風的低海拔區域,群落Ⅷ分布于坡度較緩的低山區域。物種組成上,由于常年受海風脅迫,相對于內陸地區海島植被物種多樣性較低,主要物種與大陸相似,以殼斗科、山茶科占優勢。這三類群落處于闊葉混交林幼齡階段,喬木層樹種資源較少,但灌木層成分較多,紅楠作為防風樹種,能夠在多樣性相對較低的海島植被中占據林分上層優勢地位,增長潛力大,但應注意控制種群密度,防止種內競爭。
從環境因子來看,紅楠生存群落的分布受到多種環境因子如海拔、坡度、降水的影響,主導因子是海拔高度。紅楠的生存群落在坡度較緩的低山區域有較明顯傾向性分布,體現了水熱條件對于紅楠在浙江省分布及發展的重要性,這與張志敏等[3]、胡紹慶[4]、江香梅和俞湘[6]、徐振東等[26]對紅楠分布環境的描述相符。在溫度及降水條件基本相同的地域,林下微環境決定了種群的更新和生長狀況[27]。腐殖質厚度反映了土壤肥力的不同,枯落物從一定程度上影響了地表溫度、含水量[28-29]以及植被幼苗的萌發,厚而致密的枯落物阻礙了種子著陸也影響了光向落葉層下的傳播[27]。因此紅楠在相對腐殖質更厚、枯落物更薄的群落Ⅴ、群落Ⅶ中的生長明顯比在群落Ⅲ、群落Ⅵ生長更占優勢,表明了土壤肥力、林下光照對于紅楠幼苗更新生長的重要性。充足的光照能夠提高紅楠幼苗的存活率,因此相比于郁閉度較大的群落Ⅳ,紅楠在林窗較多的群落Ⅰ中優勢更為明顯。
從種間關系來看,一定空間范圍內的不同植物個體之間存在相互作用。甜櫧、木荷、青岡是紅楠生存群落中出現頻率最高、優勢地位最為顯著的樹種,同時也是浙江省森林群落中十分穩定的優勢種[30-32]。紅楠與青岡、木荷、甜櫧等群落優勢種具有較緊密的聯結關系,再加上相對穩定的自然環境和低干擾性能夠增強群落結構的穩定性,因此紅楠能夠在這幾類優勢種群落中占據較為有利的優勢地位。但紅楠、杉木、木荷、白櫟對海拔、坡度、降水等環境因子的要求相近,因此對資源的搶奪也可能會導致種間競爭,影響紅楠的生長與更新。而且聯結結果顯示大多數種對種間聯結程度不顯著或者無關聯,表明紅楠群落整體結構不夠穩定。因此在群落恢復和發展過程中,選擇配置聯結性高的樹種以增強群落穩定性的同時,要注意環境資源的均衡分配。另外在環境條件基本一致的植被群落中,鄰株植物功能類型對紅楠的生長也有顯著影響,鄰株植物為落葉闊葉型物種時會促進紅楠的生長[33],這可能是紅楠能夠在以榆樹、白櫟為優勢種的群落中占據優勢地位的重要原因。
本研究還存在一定的局限性。研究中僅系統討論了紅楠生境地群落類型,下一步仍需對紅楠優勢林群落和伴生林群落進行比較分析,以探究影響紅楠發展的因素。此外由于本研究僅針對浙江省紅楠生境樣地,忽略了其他類型樣地中制約紅楠分布的生物及非生物因素。在以后的研究中,可以對浙江省不同類型樣地統一考慮,再根據結果對分布紅楠的主要類型區進行環境解析。全面分析的結果也可以和單獨分析紅楠生境地群落的結果進行比對,探究是否忽略了其他影響因素。
5 結論
(1)通過TWINSPAN將172個紅楠生境地群落劃分成8個群落類型。由于群落類型不同,隨著演替而導致的群落內部環境改變(如光照)也有可能是影響紅楠分布及發展的主要原因。在不同的演替階段,通過人工疏伐或補植等措施,能夠為紅楠創造更加有利的生長環境,提高紅楠種群的更新能力。
(2)海拔高度及坡度是決定紅楠分布及發展的主導環境因子。紅楠作為優勢種的生境地主要特征為海拔900m以下、坡度35°以下。在水熱條件基本相同的地域,郁閉度、枯落物厚度等決定的林下微環境決定了種群的更新和生長狀況,因此紅楠的分布也要考慮光照、土壤等其他環境因素的影響。
(3)青岡、木荷、甜櫧均為構成浙江省典型森林群落的穩定優勢種。種間聯結初步分析顯示,紅楠與青岡、木荷、甜櫧等群落優勢種具有顯著正關聯。由以上樹種構成的群落,加上相對穩定的自然環境和低干擾性,能夠增強結構穩定性,但應注意環境資源的均衡分配,減少種間競爭。
基于物種重要值的TWINSPAN群落分類法結合CCA排序法,對浙江省紅楠生境地群落取得了良好的分類結果,歸納出浙江省紅楠生境地的主要群落類型和環境特征,解釋了環境因子和種間關系對紅楠分布及發展的影響,同時從群落演替的角度提出促進紅楠生長及發展的對策,可為浙江省紅楠天然資源的恢復和人工林的經營提供數據支撐。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
