社群等級對金烏賊行為表型及能量代謝的影響
2020-09-17 00:01:28柏青青李江濤徐秀文張宗航李文濤張秀梅
生態學報 2020年15期
關鍵詞:優勢
柏青青,李江濤,徐秀文,張宗航,李文濤,張秀梅,2,*
1 中國海洋大學海水養殖教育部重點實驗室,青島 266003 2 青島海洋科學與技術國家實驗室,海洋漁業科學與食物產出過程功能實驗室,青島 266072
由于環境資源的有限和空間的異質性,動物社群等級在自然界中廣泛存在[1]。在社群等級形成過程中,個體之間產生競爭交互作用,形成優勢個體和劣勢個體[2]。優勢個體在不同程度上控制食物源、占據中心領域位置、對處于劣勢地位的個體發起主動攻擊行為、優先進行交配活動等;而劣勢個體處于長期壓力狀態,會減少自身的活動及對食物、領地、生殖等資源的競爭[3-4]。社群等級的差異導致了個體間能量代謝、激素、基因水平的差異[5-7]。一般來說,優勢個體爭斗能力較強,在激烈爭斗階段的能量主要由無氧呼吸提供[8-9],而當個體處于慢速游泳狀態或者懸浮狀態時,主要通過有氧呼吸提供能量[10-11]。社群等級形成過程中不同個體之間的競爭交互作用對于親體繁育及子代的生長發育具有顯著影響[12]。有研究表明在繁殖過程中,雄性個體之間常通過打斗來獲得交配權,但頻繁的爭斗行為也導致了交配成功率的下降,從而影響繁殖效率[13]。因而研究社群等級脅迫條件下,繁殖親體的行為表型及其能量代謝機制對于動物繁殖行為的人工調控及子代健康苗種培育具有重要意義。關于繁殖期間個體優勢等級的研究較少,僅見Jeffrey和Gilmour[12]等報道,處于劣勢地位的雌性斑馬魚壓力激素水平高,受下丘腦—腦垂體—腎間組織(Hypothalamic-pituitary-interrenal)軸基因調控且能夠遺傳到子代,從而影響子代的生長發育。金烏賊(Sepiaesculenta)也具有較強的領域行為[14],無論是與雌性交配還是護衛伴游,一旦有非護衛雄性闖入,護衛雄性立即做出反應,對非護衛雄性進行驅趕。處于優勢地位的大規格雄性常常主動攻擊小規格雄性[14]。然而關于社群等級對于金烏賊繁殖期行為表型以及能量代謝影響的研究尚未見報道。
金烏賊是我國近海經濟價值較高的頭足類之一,也是目前山東省重要的增殖放流物種。其世代周期短、產卵數量少的特性,使得金烏賊繁殖生物學的研究成為重點。目前已經開展了親體培育與人工養殖模式[15-16]、繁殖行為和繁殖模式[17]等的研究,但繁育技術仍不成熟。在金烏賊人工繁育過程中,如親體密度過高,個體間易產生激烈的競爭作用,形成社群等級。本文通過觀察繁殖期金烏賊雌雄個體社群等級的形成過程,分析不同優勢等級雌雄個體的行為表型以及能量代謝差異,探究行為表型與能量代謝的相關性,以期為金烏賊健康苗種培育以及規模化繁殖技術優化提供理論依據。
1 材料與方法
1.1 實驗材料
2018年7月于青島金沙灘金烏賊養殖基地開展實驗,所用金烏賊親體均采集于青島薛家島周邊海域(35°59′N,120°19′E)。將采集的雌雄親體分開暫養4 d,從中選取活力強、胴背部無損傷、色澤鮮明的雌雄親本共48只(24♀24♂)備用,測量并記錄實驗親本的胴背長和體重等形態學指標。親體暫養采用砂濾流水方式,持續充氧,池內鹽度30—32,溫度20—21.5℃,pH值7.7—7.8,水深控制在80 cm。每天定時投喂鮮活的凡納濱對蝦(Litopenaeusvannamei)。由于雄性個體間的爭斗噴墨,會導致水體能見度降低,每天9:00定時按50%比例換水1次。
1.2 實驗設計
實驗設置兩個實驗組,分別為兩雄一雌組(2♂1♀)和兩雌一雄(2♀1♂)組,每個實驗組設置8個平行。實驗開始前,在實驗水槽中(0.9 m×0.9 m×0.9 m)按比例放入雌雄個體,為避免規格差異對優勢等級形成的影響,在進行實驗組配對時選擇體長和體重差異小于5%雄性個體和雌性個體進行分組。(2♂1♀)組用于觀察雄性之間的競爭;(2♀1♂)組用于觀察雌性之間是否存在競爭關系。表1為實驗組中實驗個體的體重、胴長、性腺重、肝臟重等基本生物學參數。實驗開始前將實驗對象放置在水槽中適應12 h后進行實驗,通過錄像連續觀察行為變化。

表1 實驗組中金烏賊形態參數
1.3 行為分析方法
實驗連續進行4 d,選擇每天9:00—11:00以及13:00—15:00兩個時間段錄像進行行為觀察。錄像分析結果可將金烏賊的行為反應定義為兩大類7種不同行為,包括主動攻擊行為(攻擊、追逐、取代、威脅),防御行為(噴墨、后退、逃逸)。參照Johnsson和Bj?rnsson[18]、Filby等[19]的方法通過行為觀測,計算平均每小時主動攻擊和防御行為發生次數。主動攻擊行為記1分,防御行為記-1分。根據個體得分狀況判別雌雄個體的社群等級。
觀察發現,雄性個體嘗試交配數次后得以成功交配,通過觀察嘗試交配次數以及成功交配次數,可得出雄性金烏賊成功交配率。其中,雄性個體成功交配率=成功交配次數/嘗試交配次數。而雌性個體在繁殖過程中,間歇性靠近產卵基放卵,統計不同等級雌性個體平均每小時放卵次數。
觀察發現,實驗水槽中的金烏賊通常處于懸浮狀態,可將其分為游動懸浮、靜止懸浮、靜止伏底和爭斗狀態四種不同的狀態,統計單位時間內不同運動狀態的時間。
1.4 樣品的采集與測定方法
連續4 d行為觀察結束后,每組隨機取出2個實驗個體(共16♀16♂)用吸水紙吸取烏賊體表的水分,在解剖盤上進行活體解剖,取腕部肌肉和性腺組織進行超低溫保存。準確稱量各組織重量,按重量(g)∶體積(mL)=1∶9的比例加入9倍體積的生理鹽水,冰水浴條件下機械勻漿,2500 r/min,離心10 min,制備成10%的勻漿上清液置于-20℃冰箱中保存待測。取得的上清液需進行總蛋白測定,測定方法采用標準牛血清蛋白作標準曲線的考馬斯亮藍染色法。己糖激酶(Hexokinase, HK)、丙酮酸激酶(Pyruvate kinase, PK)、乳酸脫氫酶(Lactate dehydrogenase, LDH)、檸檬酸合酶(Citrate synthase, CS)、蘋果酸脫氫酶(Malate dehydrogenase, MDH)的活性和總蛋白、乳酸(Lactic acid, LD)濃度均采用南京建成生物研究所提供的試劑盒測定(A077-1、A076-1、A020-2、A108、A021-2、A019-2和A045-2),具體方法參照說明書。
計算每個實驗個體有氧代謝酶(CS、MDH)和無氧代謝酶(HK、PK、LDH)含量以及無氧代謝產物乳酸(LD)含量。通過錄像資料,對應行為分析結果,比較不同優勢等級個體間有氧和無氧代謝的差異,進一步分析每個實驗個體的運動狀態與有氧和無氧代謝酶的相關性。
1.5 數據處理
數據為平均值±標準差。優勢和劣勢個體之間對比采用T-檢驗;不同實驗組合之間對比采用單因素方差分析(one-way ANOVA)和Duncan兩兩比對。游動懸浮時間與有氧代謝酶含量以及爭斗時間與無氧代謝酶活性之間的關系采用曲線估計進行分析,并用 ANOVA方差分析對回歸的顯著性進行檢驗。所有統計分析均采用SPSS 19.0。
2 結果
2.1 社群等級的形成
根據觀察,放入同一實驗水槽不同組合的金烏賊,其社群等級一般在2 d之后形成。如圖1所示,社群等級形成初期個體間交互行為發生次數多,隨著時間推移個體間交互行為次數逐漸降低,并達到穩定狀態。雌性個體的防御行為次數無顯著差異(P>0.05),無論是主動攻擊還是防御行為的發生概率均低于雄性個體。

圖1 金烏賊個體間交互行為及社群等級的形成過程
金烏賊具有明顯的主動攻擊行為和防御行為。經計算得到優勢雄性和劣勢雄性個體的攻擊和防御行為得分分別為(11.8±1.98)、(-5.95±2.69)(P<0.05),優勢雌性個體和劣勢雌性個體分別為(1.48±1.22)、(-4.96±1.02)(P<0.05)(圖2)。觀察并記錄雄性個體嘗試交配次數與成功交配次數,結果發現優勢雄性和劣勢雄性個體的成功交配率分別為(28.27%±4.66%)、(11.94%±2.81%)(P<0.05)。優勢雌性個體和劣勢雌性個體放卵次數分別為(10.72±1.65)次/h、(1.85±0.68)次/h(P<0.05)。

圖2 金烏賊雌雄不同社群等級行為得分
2.2 不同等級金烏賊行為表型的差異
如圖3所示,優勢雄性個體游動懸浮時間和爭斗時間顯著高于劣勢個體(P<0.05);與之相反,優勢雄性個體靜止懸浮時間和靜止伏底時間顯著小于劣勢個體(P<0.05)。優勢雌性個體游動懸浮時間顯著小于劣勢個體(P<0.05),靜止伏底時間顯著高于劣勢個體(P<0.05)。雌性個體在繁殖期爭斗行為極少發生,爭斗時間和靜止懸浮時間顯示為優勢和劣勢個體二者之間無顯著性差異(P>0.05)(圖3)。

圖3 不同優勢等級個體四種不同運動狀態的時間分配
2.3 社群等級對金烏賊代謝酶活力的影響
通過比較分析肌肉組織和性腺組織有氧代謝通路中的蘋果酸脫氫酶(MDH)、檸檬酸合酶(CS)活性發現,(2♀1♂)實驗組優勢雌性個體肌肉中的MDH、CS活性分別為(170.982±11.157)U/gprot、(19.747±1.562)U/gprot,顯著低于劣勢個體MDH、CS活性(281.402±10.977)U/gprot、(24.125±1.164)U/gprot(P<0.05)。而性腺中MDH、CS活性顯著高于劣勢個體(P<0.05)。在(2♂1♀)實驗組中,優勢雄性個體肌肉的MDH、CS活性分別為(309.69±16.147)U/gprot、(34.927±1.479)U/gprot,顯著高于劣勢個體(212.131±6.936)U/gprot、(26.696±1.21)U/gprot(P<0.05),而在性腺中優勢和劣勢個體MDH活性無顯著差異(P>0.05)(圖4)。

圖4 不同優勢等級雌雄個體肌肉和性腺組織中有氧呼吸代謝酶活性(n=8)
如表2所示,優勢雄性個體肌肉中己糖激酶(HK)、丙酮酸激酶(PK)、乳酸脫氫酶(LDH)3種無氧代謝酶活性分別為(12.453±0.858)U/gprot、(18.269±0.583)U/gprot、(63.220±2.509)U/gprot,顯著高于劣勢雄性個體(P<0.05)。優勢雄性個體性腺中的無氧呼吸代謝酶活性與劣勢雄性個體之間差異不顯著(P>0.05)。優勢雌性個體和劣勢雌性個體肌肉和性腺之間無氧代謝酶活性差異均不顯著(P>0.05)。
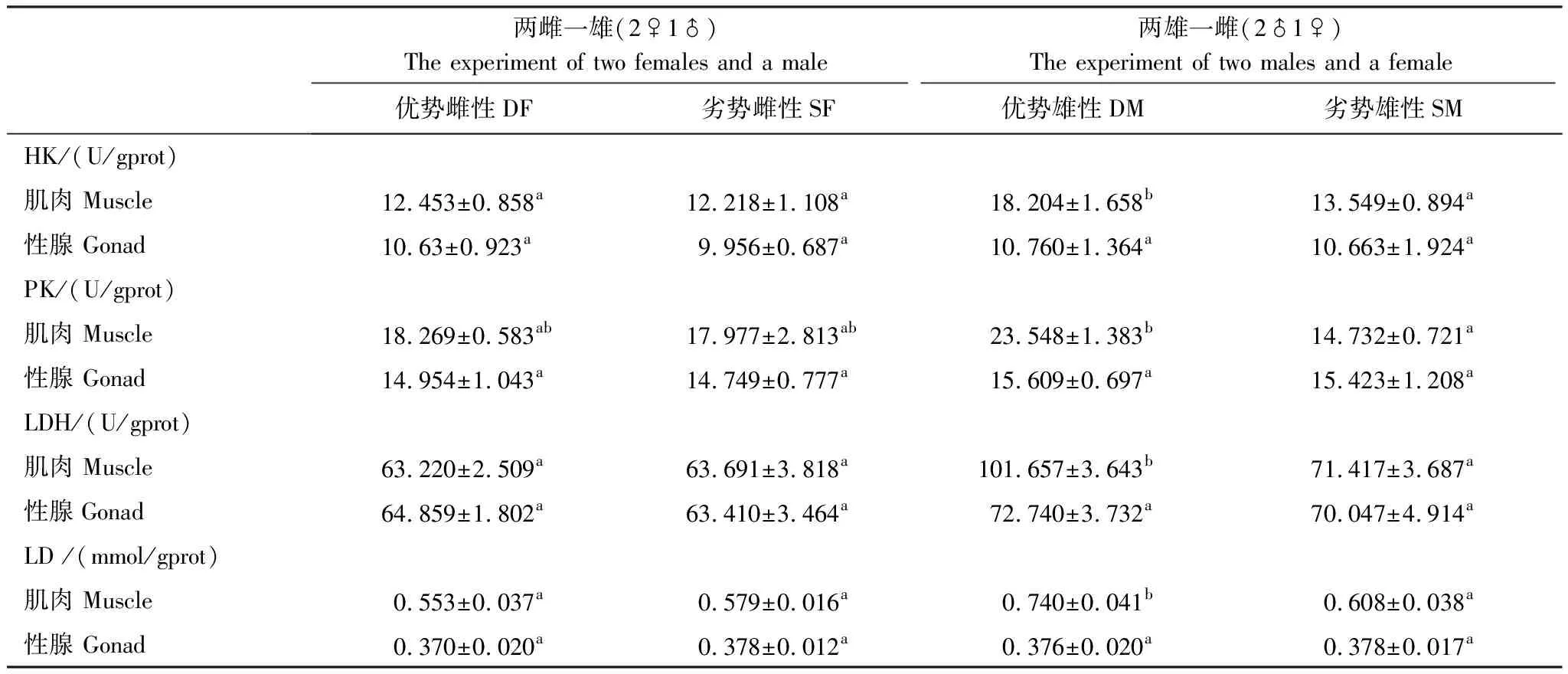
表2 不同優勢等級雌雄個體肌肉和性腺組織中無氧代謝酶活性及乳酸含量
2.4 繁殖期金烏賊行為表型與代謝酶活力的相關性
比較分析發現社群等級對于金烏賊行為表型和能量代謝具有顯著影響,不同優勢等級雌雄親體的游動懸浮時間分別與蘋果酸脫氫酶(MDH)、檸檬酸合酶(CS)兩種有氧代謝酶活性之間呈線性相關:y=5.093x+99.519(R2=0.367,F=17.419,P<0.001)、y=0.510x+11.940(R2=0.706,F=2.165,P<0.001)(圖5)。
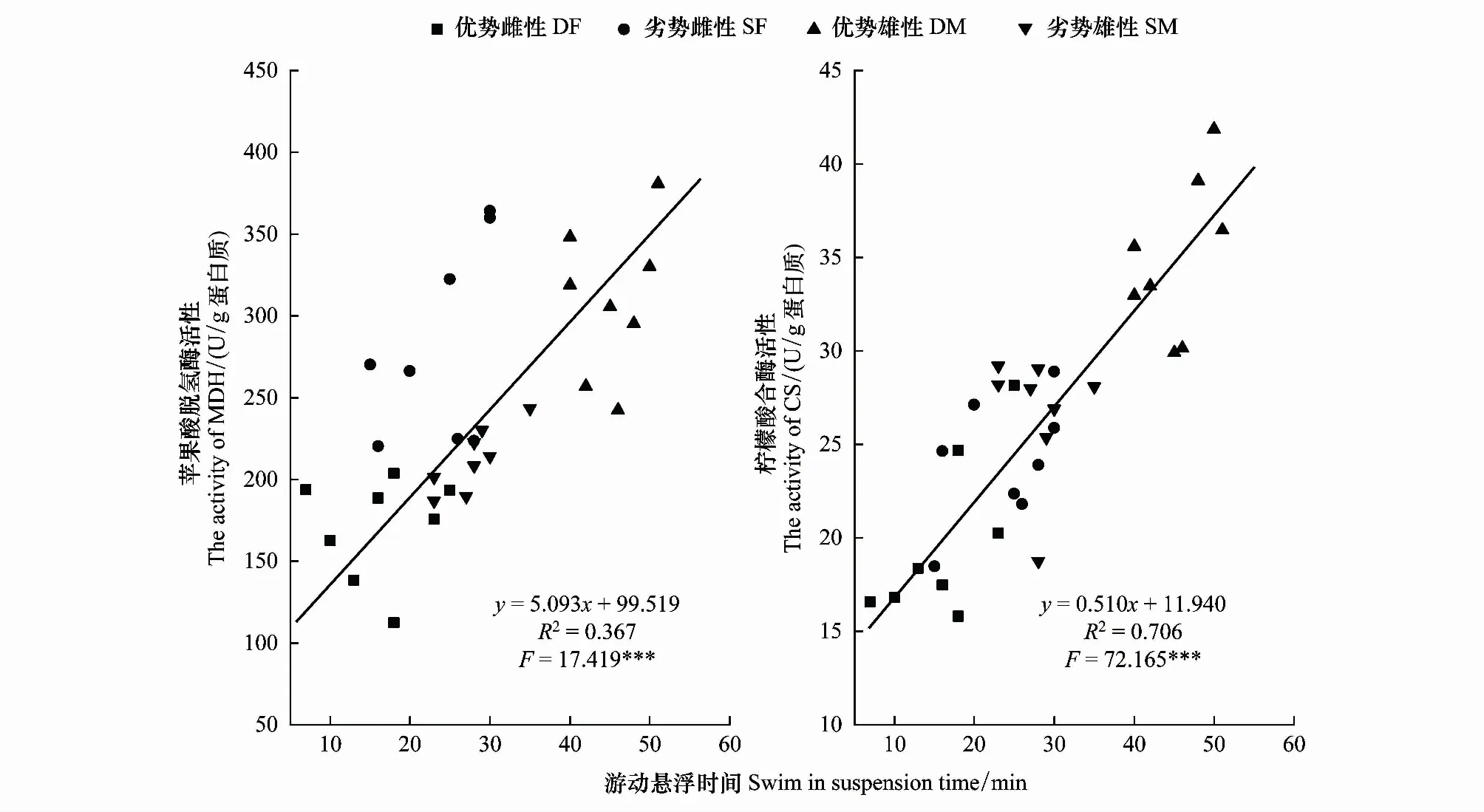
圖5 金烏賊游動懸浮時間與肌肉中有氧代謝酶活性的關系
同理分析可得,個體的爭斗時間與己糖激酶(HK)、丙酮酸激酶(PK)、乳酸脫氫酶(LDH)活性之間的關系呈顯著正相關,擬合的曲線方程分別為y=1.072x+1.559(R2=0.578,F=41.141,P<0.001)、y=0.770x+1.946(R2=0.137,F=4.762,P<0.05)、y=4.671x+63.902(R2=0.541,F=37.737,P<0.05)(圖6)。進一步分析爭斗時間與乳酸(LD)含量之間的關系發現,隨爭斗時間的增加,金烏賊腕部肌肉乳酸含量顯著增加(P<0.001)(圖6)。這表明金烏賊在發生劇烈爭斗行為時,運動時間短,主要通過無氧代謝供應能量。

圖6 金烏賊爭斗時間與肌肉中無氧代謝酶活性和乳酸含量的關系
3 結論和討論
3.1 行為交互作用與社群等級之間的關系
社群等級的形成對個體行為產生很大影響,社群等級高的個體具有較強的攻擊性,能夠占據主要食物源和領地[20]。在社群等級脅迫下,劣勢個體處于長期壓力狀態,表現為運動能力下降,并且影響其生長、代謝、免疫等功能[21]。有研究表明,劣勢魚在遭到優勢魚的“視力”威脅后,會出現躲避、減小運動范圍等行為[22]。本研究中,雄性個體之間具有較明顯的爭斗行為,社群等級現象顯著。優勢個體主動用腕迅速靠近劣勢個體并擊打劣勢個體的胴部,同時伴隨劣勢個體的躲避、噴水、噴墨等防御行為。優勢雄性個體長時間懸浮伴游在雌性個體周邊,劣勢雄性嘗試靠近后被優勢個體阻攔,靜止懸浮在水體中或者底部。而雌性個體之間未發現明顯的爭斗行為,但優勢雌性個體與雄性個體交配次數多、放卵次數多,多次放卵后會停留在水層底部,呈現靜止狀態。
3.2 社群等級與能量代謝之間的關系
Li等[23]研究發現,水生動物低強度運動時主要通過有氧代謝獲取能量,隨運動強度增加,能量代謝方式由有氧代謝轉為厭氧糖酵解,原因主要是與氧氣供應有關[10]。已有研究發現己糖激酶(HK)、丙酮酸激酶(PK)、乳酸脫氫酶(LDH)對糖酵解速率控制發揮關鍵作用,產生乳酸(LD)等代謝產物[24-26]。MDH能催化蘋果酸與草酰乙酸之間的可逆轉換,CS在三羧酸循環中可催化草酰乙酸和乙酰輔酶A之間的縮合反應,二者都是三羧酸循環的關鍵酶[27-28]。低強度運動氧氣供應充足,因此動物通過有氧代謝獲取能量[11]。本研究中,由于優勢雄性個體長時間處于游動懸浮狀態,主要依靠腕部肌肉和腹部肌肉輕微收縮運動,其肌肉有氧代謝酶(MDH、CS)活性顯著高于劣勢個體,進一步證實了金烏賊在游動懸浮時主要進行有氧代謝活動。
社群等級形成過程中,個體間通過爭奪配偶以及領域宣誓主權,易產生劇烈爭斗行為[29]。而動物在產生急速運動或劇烈爭斗時,短時間內需要提供大量能量,主要由無氧呼吸提供[30]。在凡納濱對蝦(Litopenaeusvannamei)、尼羅羅非魚(Oreochromisniloticus)中隨著游泳速度的增大,無氧呼吸酶活力顯著增加[9,26]。本研究發現,優勢雄性個體的無氧代謝酶活性(PK、HK、LDH)顯著高于劣勢雄性個體,分析認為優勢雄性個體主動發起急速攻擊次數多,時間短,腕部以及胴部肌肉劇烈收縮和拉伸,無氧代謝活動加劇。在岸蟹(Carcinusmaenas)、寄居蟹(Birguslatro)中發現由于爭斗導致體內乳酸含量的增加[31-32],本研究中優勢雄性個體肌肉中乳酸含量也顯著高于劣勢雄性個體。而雌性個體之間爭斗行為少,無激烈競爭交互作用,無氧代謝酶活性差異不顯著。無氧代謝酶活性及乳酸含量與雄性爭斗時間呈顯著正相關,也進一步證實了金烏賊在爭斗過程中主要以無氧代謝供能。
3.3 繁殖期雌雄個體的能量分配
研究表明,繁殖是一個高能量需求過程,性腺的生長發育以及卵子的形成等都需要能量供應[33-34]。由于動物攝入的食物資源有限,即能量攝入有限,用于繁殖能量的增加必然會導致用于其他生命活動的能量支出減少[33]。在“生物行為節能假說”中也提到繁殖與運動行為間存在能量競爭關系;動物通過減少自身運動時間,將能量優先用于繁殖過程[35-36]。在自然條件下,為了保證個體適應性,繁殖期的生物常通過降低生長率、減少運動能量消耗等來保證生殖成功,這一過程也稱為“生殖代價”[33,37],合理的能量投資能夠使生殖價最大化。Ru等[38-39]研究發現仿刺參(Apostichopusjaponicus)在繁殖過程中通過減少運動時間和運動距離以及降低生長率、攝食率從而減小能量分配以維持繁殖過程中的高能量需求。而在魚類中同樣有此現象發生,繁殖期的短腳床杜父魚(Myoxocephalusscorpius)和食蚊魚(Gambusiaaffinis)的最大游泳速度出現顯著下降[40-41]。研究表明,頭足類在繁殖期減少甚至停止攝食,Quetglas等研究發現尖盤愛爾斗蛸(Eledonecirrhosa)的性腺重量在非繁殖期占體重的1%,而在繁殖期達到了體重的20%—25%[42]。在減小或者停止外源性能量攝入的同時,機體儲能減少,而性腺重量的增加,加大了機體的物理負擔,導致運動能力下降。本研究中優勢雌性個體產卵后會長時間靜止伏底或靜止懸浮于水體中,自發活動明顯降低,用于運動的能量減少,肌肉的有氧代謝酶活性顯著低于劣勢個體,但性腺有氧代謝酶活性顯著高于劣勢個體,推測是優勢雌性個體大部分能量用于卵子的生成和發育所致。綜上,行為的可塑性是繁殖期能量分配的一種有效策略,雌性金烏賊通過減少運動能耗來保障繁殖的高能量需求。
繁殖期雌雄個體具有不同的能量分配方式,尼羅羅非魚(Oreochromisniloticus)雄魚比雌魚生長速度快,在繁殖期間雌雄生長差異更加顯著[43],雌性個體將更多的能量分配到生殖上,用于生長的能量減少。繁殖期中華絨螯蟹(EriocheirSinensis)雌蟹將44%能量分配至性腺和肝胰腺中,高于雄蟹的23%[44]。本研究中雄性肌肉中的MDH和CS兩種有氧代謝酶活分別是雌性個體的1.2倍和1.4倍,繁殖期雄性個體運動能耗高于雌性個體。而雌性個體性腺中的MDH和CS酶活分別是雄性個體的1.6倍和1.1倍,雌性個體將更多能量用于配子形成和性腺發育。而雄性個體間通過相互爭斗,獲取更多交配機會,將精莢粘附在雌性個體口膜處,并伴游護衛雌性個體,以保證自己的繁殖成功率,因而將更多能量用于爭斗和運動中。
4 總結
金烏賊人工繁育過程中,社群等級的形成使得不同等級雌雄個體間的行為產生差異,優勢雄性個體以交配投資為主,因而具有更強的攻擊性、爭斗和運動能力。雌性個體以產出更多的卵子作為主要生殖投資,具有較強的領域性,優勢雌性個體用于產卵繁殖的能量較多。行為活動的差異也導致了繁殖親體有氧和無氧代謝酶活性的差異。有關金烏賊社群等級的差異是否會影響親體的繁殖效率以及子代的質量等仍需要進一步研究。金烏賊為一年生頭足類,繁殖期經多次交配多次產卵,繁殖后快速死亡。在其短暫的生命周期中,生殖代價與衰老和死亡之間錯綜復雜的關系仍有待考證。
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
華人時刊(2020年13期)2020-09-25 08:21:30
趣味(語文)(2020年3期)2020-07-27 01:42:46
意林·全彩Color(2019年8期)2019-09-23 02:12:36
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
中國體育(2004年3期)2004-11-11 08:53:02
棋藝(2001年19期)2001-11-25 19:55:34
棋藝(2001年23期)2001-01-06 19:08:36
