利用CRISPR-Cas9技術(shù)和Cre/loxP系統(tǒng)構(gòu)建豬偽狂犬病病毒gE基因缺失疫苗株
2020-09-27 04:06:52史志斌馬寧寧劉占王永生陳陸
畜牧與獸醫(yī) 2020年10期
關(guān)鍵詞:小鼠
史志斌,馬寧寧,劉占,王永生,陳陸
(河南農(nóng)業(yè)大學(xué)牧醫(yī)工程學(xué)院動(dòng)物傳染病教研室,河南 鄭州 450046)
偽狂犬病病毒(pseudorabies virus,PRV)屬皰疹病毒科α皰疹病毒亞科[1],引起的豬偽狂犬病(porcine pseudorabies)嚴(yán)重危害全球養(yǎng)豬業(yè)[2]。豬急性感染PRV后引起妊娠母豬流產(chǎn)、產(chǎn)死胎及木乃伊胎、種豬不育、新生仔豬腦脊髓炎等[3-4]。急性感染耐過(guò)豬和隱性感染豬能建立終生潛伏感染,潛伏的病毒存在于三叉神經(jīng)節(jié)、嗅球和扁桃體等組織,在應(yīng)激狀態(tài)下活化并向外排毒,因此,潛伏感染豬是偽狂犬病潛在傳染源[5-7]。PRV潛伏感染特性給豬偽狂犬病的防控帶來(lái)巨大困難。目前,防控本病主要依靠疫苗免疫,生產(chǎn)中常用的偽狂犬病疫苗主要是PRV毒力基因(如TK、gE、gI等)或糖蛋白基因(如gC、gG等)缺失的基因工程苗[8],尤其以TK和gE基因的缺失最為重要。gE基因缺失可降低PRV神經(jīng)嗜性,也可作為診斷標(biāo)志,通過(guò)對(duì)gE糖蛋白誘導(dǎo)抗體的血清學(xué)檢測(cè)鑒別疫苗免疫豬和野毒感染帶毒豬,為偽狂犬病的凈化提供技術(shù)支持[9-11]。2012年始,偽狂犬病在我國(guó)傳統(tǒng)疫苗免疫場(chǎng)再次出現(xiàn)暴發(fā)和流行,新出現(xiàn)的流行株表現(xiàn)毒力增強(qiáng)。為了研發(fā)針對(duì)當(dāng)前流行株的疫苗,本文首先從實(shí)驗(yàn)室分離的4株P(guān)RV流行株篩選高免疫原性親本株,然后利用CRISPR-Cas9技術(shù)和Cre/loxP系統(tǒng)對(duì)親本株gE基因進(jìn)行缺失,并對(duì)構(gòu)建的gE基因缺失株制備的滅活疫苗的免疫效力進(jìn)行評(píng)價(jià),為偽狂犬病標(biāo)志疫苗的研制提供參考。
1 材料與方法
1.1 主要試劑
EcoRⅠ、SpeⅠ、HindⅢ、BbsⅠ和T4DNA連接酶購(gòu)自TaKaRa公司;低熔點(diǎn)瓊脂糖購(gòu)自北京索萊寶科技有限公司;2×DMEM高糖培養(yǎng)基購(gòu)自杭州吉諾生物醫(yī)藥技術(shù)有限公司。
1.2 病毒和細(xì)胞
PRVHNQYY2012株、HNQXX2012株、HNQXY2012株、HNQBA2012株,Vero和293T細(xì)胞由本實(shí)驗(yàn)室保存。
1.3 質(zhì)粒
pMD18-T載體購(gòu)自TaKaRa公司;pX330和DH5α由本實(shí)驗(yàn)室保存;pcGlobin2-Cre質(zhì)粒購(gòu)自上海權(quán)陽(yáng)生物科技有限公司;pEGFP質(zhì)粒由本實(shí)驗(yàn)室合成保存。
1.4 實(shí)驗(yàn)動(dòng)物
6周齡昆明小鼠購(gòu)自河南省實(shí)驗(yàn)動(dòng)物中心。
1.5 方法
1.5.1 引物設(shè)計(jì)
根據(jù)GenBank公布的PRV基因序列(登錄號(hào):KP722022.1),使用軟件Primer 5.0設(shè)計(jì)gE基因上下游同源臂的擴(kuò)增引物。利用http://crispr.mit.Edu設(shè)計(jì)靶向PRV gE基因的單鏈引導(dǎo)RNA(single guide RNA,sgRNA)。序列信息見(jiàn)表1。

表1 引物及sgRNA序列信息
1.5.2 PRV親本株篩選
將HNQYY2012株、HNQXX2012株、HNQXY2012株、HNQBA2012株稀釋至104TCID50/0.1 mL后滅活,病毒液和Montanide ISA 201 VG佐劑按體積比1∶1比例混合乳化,制成豬偽狂犬病滅活疫苗。25只昆明小鼠隨機(jī)均分5組,1~4組分別皮下注射不同毒株制備的滅活疫苗,200 μL/只(104TCID50/只),最后1組注射DMEM培養(yǎng)基200 μL/只作對(duì)照。一免2周后進(jìn)行二次免疫,一免2周、二免2周和二免4周時(shí)分別采血并測(cè)定中和抗體效價(jià),選擇抗體水平高、免疫原性好的毒株用于構(gòu)建gE基因缺失株。
1.5.3 病毒基因組提取
HNQYY2012株接毒于Vero細(xì)胞,待細(xì)胞病變80%時(shí)收毒。酚氯仿法抽提病毒DNA后溶于TE,-20 ℃保存。
1.5.4 轉(zhuǎn)移載體的構(gòu)建
以HNQYY2012株基因組為模板,利用引物gEL-f/gEL-r和gER-f/gER-r分別擴(kuò)增gE基因上、下游同源臂gEL和gER。PCR反應(yīng)條件為95 ℃ 5 min;95 ℃ 30 s,60 ℃ 30 s,72 ℃ 80 s,30個(gè)循環(huán);72 ℃ 10 min。gEL和gER分別用EcoRⅠ/SpeⅠ和SpeⅠ/HindⅢ雙酶切,純化后克隆到EcoRⅠ/HindⅢ雙酶切的pMD18-T載體,獲得重組質(zhì)粒pTgE。pEGFP質(zhì)粒經(jīng)NheⅠ/SpeⅠ雙酶切后,純化獲得EGFP片段并克隆到pTgE質(zhì)粒上,得到重組載體pTgE-EGFP。
靶向PRV gE基因的sgRNA退火成雙鏈,將pX330質(zhì)粒用BbsⅠ酶切,形成黏性末端,經(jīng)T4 DNA連接酶連接后,雙鏈sgRNA與pX330質(zhì)粒連接構(gòu)成質(zhì)粒pX330-sgRNA。
1.5.5 HNQYY2012ΔgE/EGFP+的獲得和純化
利用Lipofectamine?2000 Reagent轉(zhuǎn)染。將HNQYY2012基因組、pX330-sgRNA和pTgE-EGFP按質(zhì)量比1∶1∶2混于250 μL Opti-MEM,獲得溶液A;250 μL Opti-MEM加入10 μL Lipofectamine?2000混勻,獲得溶液B;溶液A和B輕緩混勻,室溫靜置20 min后緩慢滴加到293T細(xì)胞,36 h后-80 ℃凍融2次備用。處理后的細(xì)胞培養(yǎng)液(含細(xì)胞碎片)8 000 r/min離心5 min,收集上清液,經(jīng)0.22 μm濾器過(guò)濾除菌并接種Vero細(xì)胞,PRV的基因組感染特性導(dǎo)致Vero細(xì)胞產(chǎn)生病變,達(dá)到70%時(shí)收獲病毒,蝕斑純化法挑取帶綠色熒光空斑得到純化病毒,命名為HNQYY2012ΔgE/EGFP+。
1.5.6 HNQYY2012ΔgE株的獲得與純化
共轉(zhuǎn)染HNQYY2012ΔgE/EGFP+基因組和pcGlobin2-Cre質(zhì)粒到293T細(xì)胞內(nèi),pcGlobin2-Cre質(zhì)粒表達(dá)產(chǎn)生的Cre酶識(shí)別HNQYY2012ΔgE/EGFP+基因組EGFP基因兩端同向的Loxp序列實(shí)現(xiàn)EGFP基因的敲除。參照1.5.5方法獲得含HNQYY2012ΔgE基因組的上清液并接種Vero細(xì)胞。蝕斑純化直至所有細(xì)胞病變均不帶熒光時(shí)獲得HNQYY2012ΔgE株。
1.5.7 HNQYY2012ΔgE株的PCR鑒定
酚氯仿法抽提HNQYY2012ΔgE病毒基因組,用引物gEL-f和gER-r進(jìn)行PCR鑒定,并對(duì)PCR產(chǎn)物進(jìn)行測(cè)序。
1.5.8 HNQYY2012株親本株與ΔgE株一步生長(zhǎng)曲線(xiàn)
HNQYY2012親本株和HNQYY2012ΔgE以MOI=0.5接種Vero細(xì)胞,接毒后12、24、36、48、60、72 h收獲病毒,測(cè)定TCID50并繪制一步生長(zhǎng)曲線(xiàn)。
1.5.9 HNQYY2012ΔgE株LD50測(cè)定
35只昆明鼠隨機(jī)均分成7組,1~3組分別皮下注射102、103和104的TCID50的PRV親本毒,4~6組分別皮下注射102、103和104的TCID50的HNQYY2012ΔgE,第7組為DMEM的對(duì)照組。觀(guān)察1周,Karber法計(jì)算LD50。
1.5.10 豬偽狂犬病滅活疫苗(HNQYY2012ΔgE株)的制備和免疫效力試驗(yàn)
HNQYY2012ΔgE株用Vero細(xì)胞增殖并定量為2×106TCID50/0.1 mL。0.3%的甲醛滅活后與Montanide ISA 201 VG佐劑按體積比1∶1乳化,制備成滅活疫苗。30只昆明鼠隨機(jī)均分成6組,1~3組免疫滅活疫苗0.1 mL/只,4~6組為不做免疫的對(duì)照組。1~3組二免二周后和對(duì)照4~6組分別以5LD50、10LD50和20LD503種劑量攻毒,統(tǒng)計(jì)小鼠死亡情況。
1.6 統(tǒng)計(jì)與分析
利用SPSS 20.0對(duì)所得統(tǒng)計(jì)數(shù)據(jù)進(jìn)行顯著性分析,采用單因素方差分析進(jìn)行顯著性檢驗(yàn),多重比較采用新復(fù)極差法(SSR),設(shè)置顯著性水平為0.05。最后用作圖軟件GraphPad Prism 6進(jìn)行制圖,數(shù)據(jù)表示的形式為“平均值±標(biāo)準(zhǔn)差”。
2 結(jié)果與分析
2.1 PRV親本株篩選
如圖1所示,在一免2周時(shí)4種毒株中和抗體水平均顯著高于DMEM培養(yǎng)基對(duì)照組(P<0.05),但4種毒株之間差異不顯著(P>0.05);在二免2周和二免4周時(shí)HNQYY2012和HNQXY2012株在抗體水平上相比其他毒株明顯升高(P<0.05)。HNQYY2012株優(yōu)于HNQXY2012株,因此選擇HNQYY2012株為親本株構(gòu)建PRV gE基因缺失株。
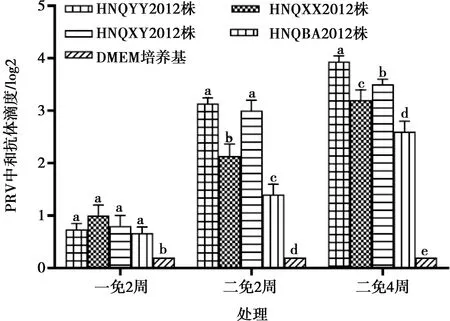
同一處理內(nèi)不同小寫(xiě)字母表示差異顯著(P<0.05),相同字母表示差異不顯著(P>0.05)
2.2 轉(zhuǎn)移載體的構(gòu)建
以pTgE為對(duì)照,用引物gEL-f/gER-r進(jìn)行PCR擴(kuò)增,pTgE-EGFP擴(kuò)增產(chǎn)物長(zhǎng)度大于對(duì)照組(圖2),PCR產(chǎn)物測(cè)序結(jié)果表明擴(kuò)增的條帶包括gE上游同源臂、EGFP序列和gE下游同源臂,證明成功構(gòu)建轉(zhuǎn)移載體pTgE-EGFP。pX330-sgRNA進(jìn)行雙酶切鑒定,酶切后呈單一條帶,未連接的PX330空載體酶切后出現(xiàn)2個(gè)條帶(圖3),證明成功構(gòu)建了pX330-sgRNA質(zhì)粒。
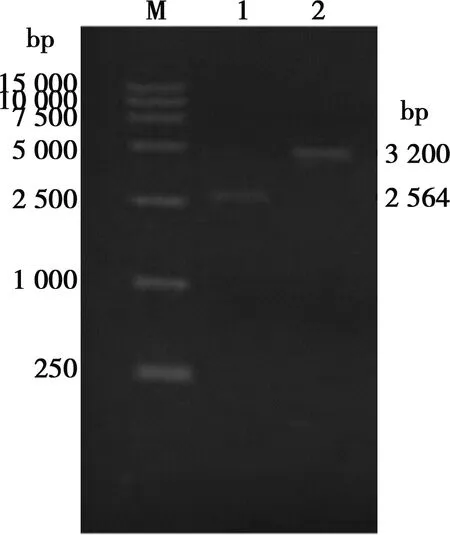
M.15 000 bp DNA Marker;1.pTgE;2.pTgE-EGFP
M.15 000 bp DNA Marker;1.pX330-sgRNA的EcoRⅠ和BbsⅠ雙酶切;2.pX330的EcoRⅠ和BbsⅠ雙酶切
2.3 重組病毒HNQYY2012ΔgE的獲得與純化
pX330-sgRNA作用下,HNQYY2012基因組和pTgE在293T細(xì)胞內(nèi)發(fā)生重組,收獲樣品后接種Vero細(xì)胞,72 h后出現(xiàn)明顯帶綠色熒光的細(xì)胞病變,說(shuō)明有重組病毒產(chǎn)生。經(jīng)過(guò)7輪噬斑純化,得到重組病毒HNQYY2012ΔgE/EGFP+(圖4)。在Vero細(xì)胞上擴(kuò)增HNQYY2012 ΔgE/EGFP+并提取基因組,以相同方法將HNQYY2012ΔgE/EGFP+基因組和pcGlobin2-Cre質(zhì)粒共轉(zhuǎn)染293T細(xì)胞,獲得重組病毒HNQYY2012ΔgE基因組后再次接種Vero細(xì)胞,72 h后出現(xiàn)不帶綠色熒光的細(xì)胞病變。相同方法純化后獲得純度較高的重組病毒HNQYY2012 ΔgE。
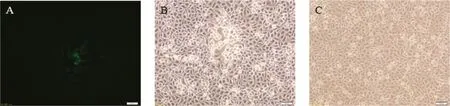
A.HNQYY2012ΔgE/EGFP+重組病毒接種Vero細(xì)胞;B.HNQYY2012ΔgE/EGFP+重組病毒接種Vero細(xì)胞;C.正常Vero細(xì)胞對(duì)照
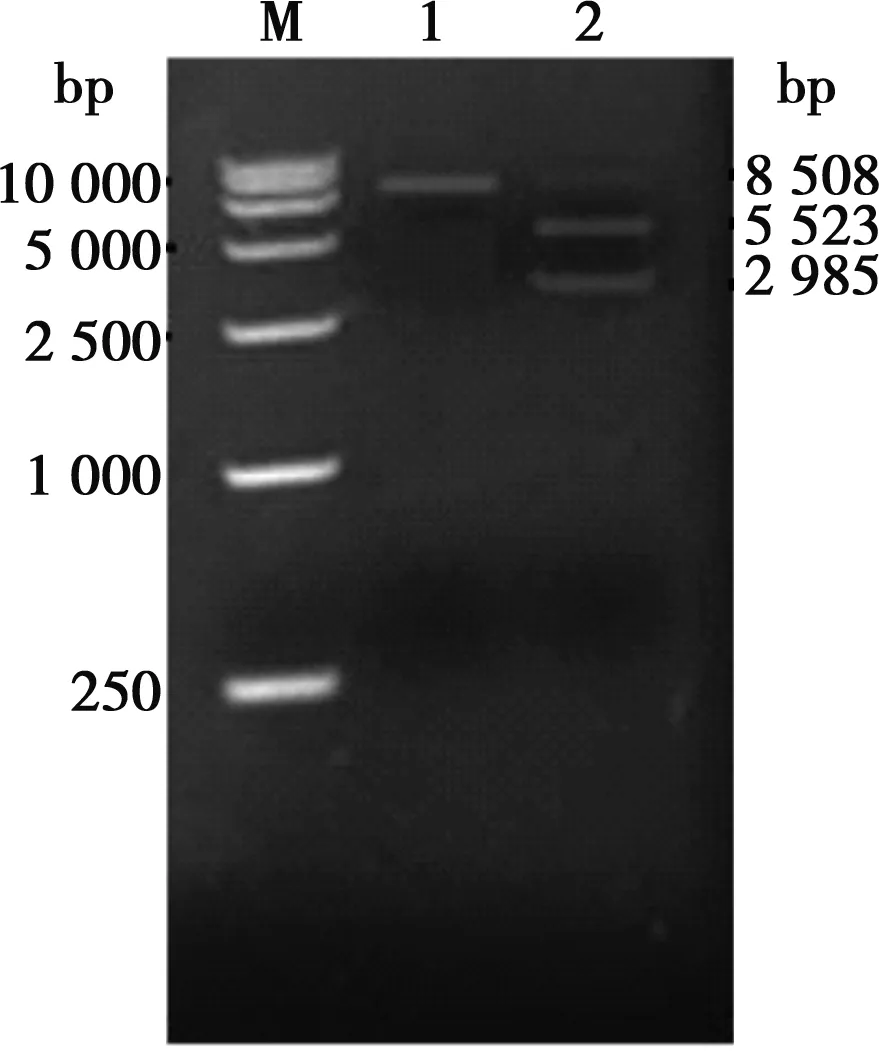
以重組病毒HNQYY2012ΔgE和HNQYY2012ΔgE/EGFP+基因組為模板,用引物gEL-f/gER-r擴(kuò)增gE基因。重組病毒HNQYY2012ΔgE/EGFP+擴(kuò)增之后在4 400 bp的位置出現(xiàn)條帶,而重組病毒HNQYY2012ΔgE在2 600 bp的位置出現(xiàn)條帶(圖5)。對(duì)重組病毒HNQYY2012ΔgE的PCR產(chǎn)物進(jìn)行測(cè)序,結(jié)果表明HNQYY2012ΔgE重組病毒gE基因完全缺失。
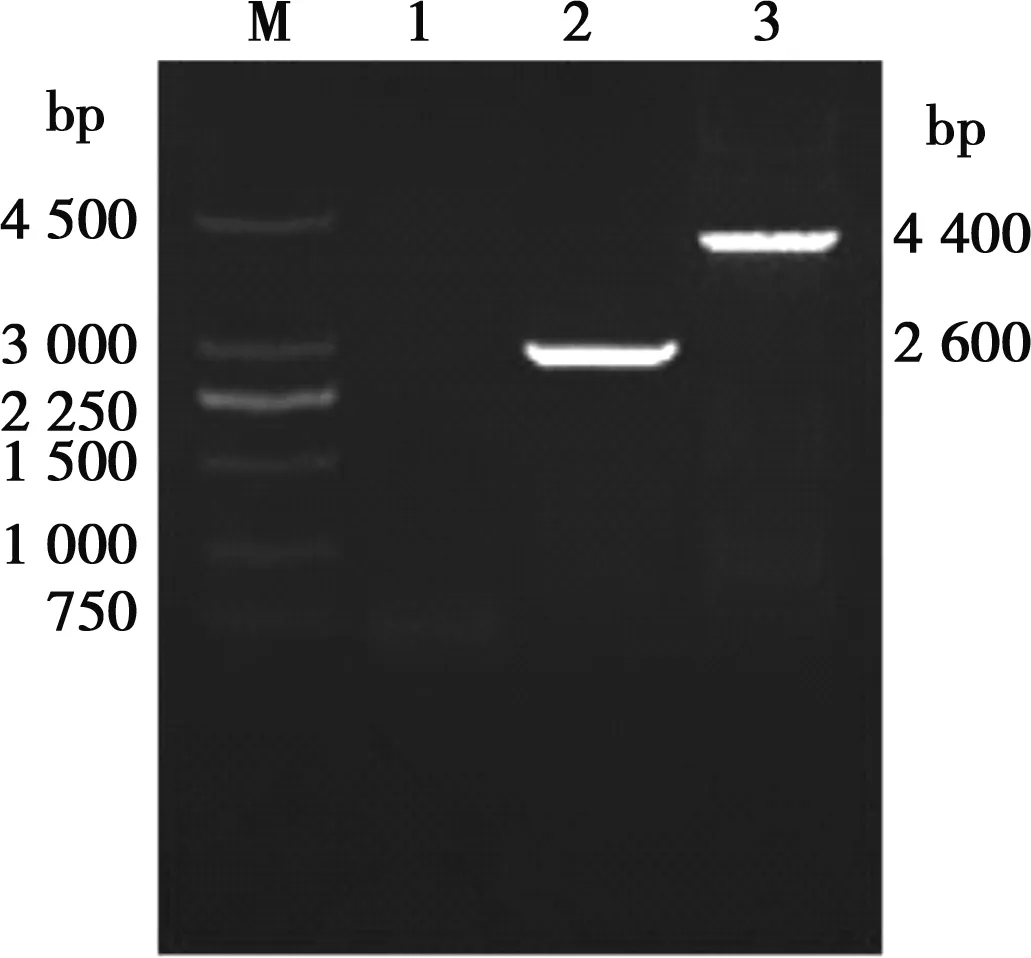
M.250 bp DNA Marker;1.空白對(duì)照組;2.gEL-f/gER-r對(duì)HNQYY2012ΔgE的PCR擴(kuò)增;3.gEL-f/gER-r對(duì)HNQYY2012ΔgE/EGFP+的PCR擴(kuò)增
2.4 HNQYY2012ΔgE和親本毒一步生長(zhǎng)曲線(xiàn)的測(cè)定
由圖6可知HNQYY2012ΔgE和親本毒的最高滴度分別為106.8TCID50/0.1 mL和107.0TCID50/0.1 mL,差異不顯著(P>0.05),HNQYY2012ΔgE的病毒滴度和親本毒差異不大。在24、36、60 h時(shí)間點(diǎn),親本毒滴度極顯著高于HNQYY2012ΔgE(P<0.01或P<0.001),說(shuō)明gE基因缺失對(duì)PRV的滴度和增殖速度影響較小。
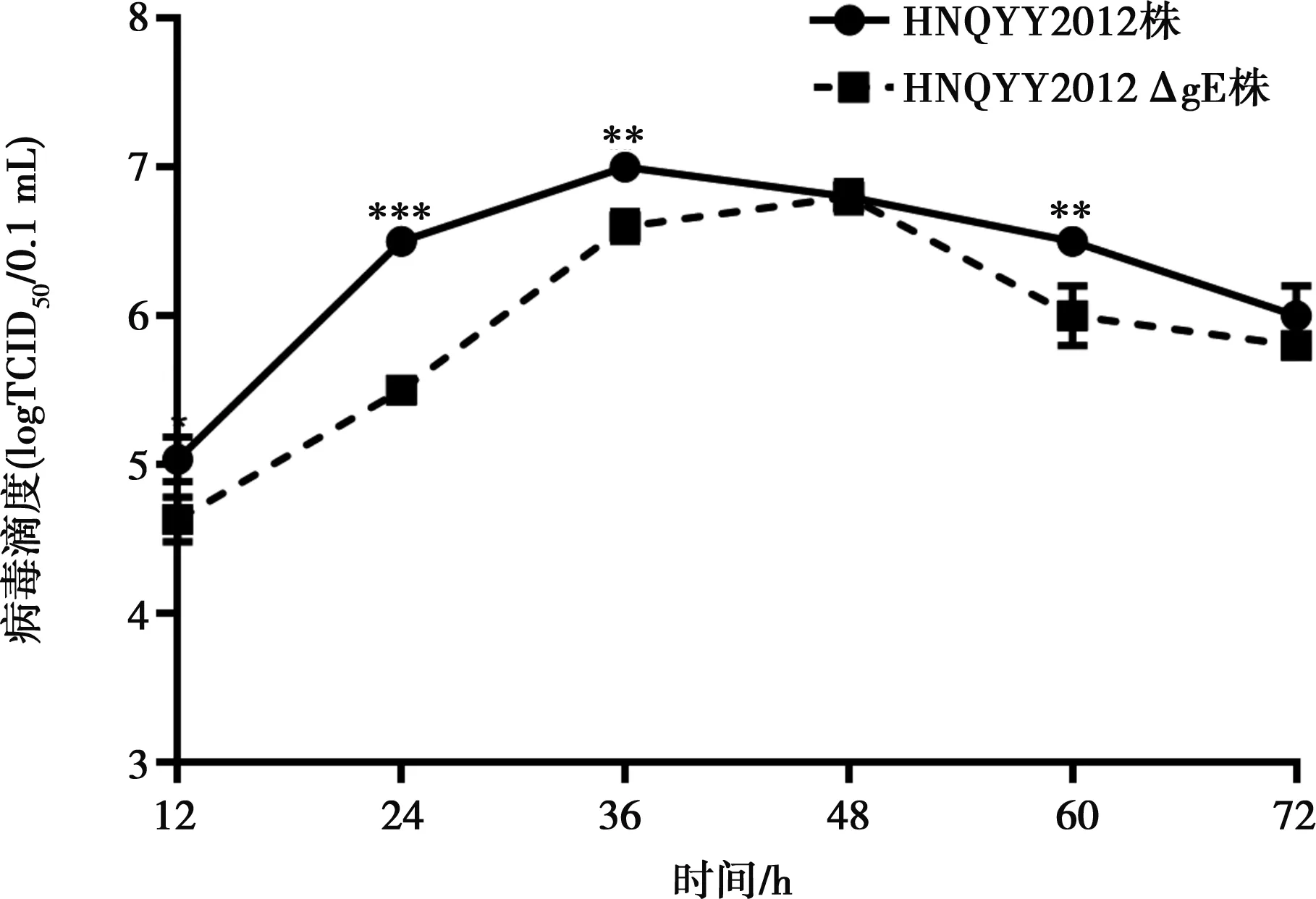
**P<0.01,***P<0.001
2.5 HNQYY2012ΔgE和親本毒LD50測(cè)定
測(cè)定HNQYY2012ΔgE和親本毒LD50分別為103.8TCID50和102.5TCID50,差異顯著(P<0.05),說(shuō)明gE基因缺失后PRV的毒力有一定程度下降。
2.6 滅活疫苗(HNQYY201ΔgE株)的免疫效力試驗(yàn)
滅活疫苗(HNQYY2012ΔgE株)免疫小鼠對(duì)5LD50、10LD50劑量攻毒保護(hù)率均為100%,20LD50的攻毒劑量組有1只小鼠存活,保護(hù)率為20%,3組對(duì)照組保護(hù)率均為0。結(jié)果表明豬偽狂犬病HNQYY2012 gE基因缺失株滅活疫苗具有良好免疫效力。小鼠生存曲線(xiàn)如圖7所示。
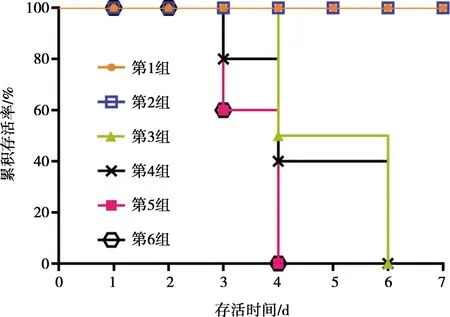
圖7 HNQYY2012ΔgE滅活疫苗免疫小鼠攻毒后生存曲線(xiàn)
3 討論
CRISPR-Cas9基因編輯技術(shù)是從細(xì)菌和古細(xì)菌中發(fā)展演變而來(lái),可以對(duì)靶基因進(jìn)行敲除和突變,其技術(shù)成熟,成本較低,在活體動(dòng)物、細(xì)胞和病毒中都有廣泛的應(yīng)用[12-14]。徐廣軍等[15]利用CRISPR-Cas9技術(shù)建立了GTP蛋白酶M(IRGNs)基因缺失的MDCK細(xì)胞系。IRGNs是由干擾素誘導(dǎo)細(xì)胞產(chǎn)生的,在抵御病原微生物中發(fā)揮著核心的作用。通過(guò)敲除MDCK細(xì)胞的IRGMs,可以進(jìn)一步研究IRGMs基因在細(xì)胞內(nèi)抵御病原微生物感染中的作用。吳濤等[16]利用CRISPR-Cas9基因編輯技術(shù)對(duì)腸道病毒71型(EV71)的VP4區(qū)域進(jìn)行了剪切,從而抑制了EV71病毒的復(fù)制,為治療EV71提供了新的途徑。Whitworth等[17]利用CRISPR-Cas9技術(shù)分別敲除了豬的CD163和CD1D基因,這2個(gè)基因被認(rèn)為是豬繁殖與呼吸綜合征病毒(PRRSV)的受體基因和抗原遞呈因子,通過(guò)敲除這2個(gè)基因后,豬體對(duì)PRRSV具有良好的抗病能力,證明CRISPR-Cas9可以成功地應(yīng)用到動(dòng)物疫病防控策略中。Cre-loxp系統(tǒng)也常被用于進(jìn)行病毒基因組重組的系統(tǒng)。梁苑燕等[18]首先構(gòu)建出了帶有l(wèi)oxp位點(diǎn)和EGFP的PRV gE-/EGFP+株,之后通過(guò)Cre酶處理后,得到了不帶EGFP的PRV gE-株,之后又利用同樣的方法得到了PRV gE-/TK-株。Xu等[19]利用CRISPR-Cas9和Cre-loxp系統(tǒng)同時(shí)對(duì)PRV的gE/TK基因進(jìn)行缺失,成功構(gòu)建了PRV gE-/TK-株。本試驗(yàn)利用CRISPR-Cas9技術(shù)和Cre-loxp系統(tǒng)對(duì)PRV gE基因進(jìn)行缺失,經(jīng)過(guò)PCR和測(cè)序鑒定,證明gE基因得到完全缺失。試驗(yàn)結(jié)果表明,利用CRISPR-Cas9技術(shù)和Cre-loxp系統(tǒng)進(jìn)行病毒的基因缺失是可行且高效的,同時(shí)以2012年后新出現(xiàn)的高免疫原性PRV流行株為親本株構(gòu)建的gE基因缺失株并制備滅活疫苗更符合市場(chǎng)需求。
HNQYY2012ΔgE株和親本株的一步生長(zhǎng)曲線(xiàn)結(jié)果比較發(fā)現(xiàn),兩者的增殖最高滴度差異不顯著,但gE基因的缺失導(dǎo)致PRV在細(xì)胞上的增殖速度減緩。Wang等[11]研究發(fā)現(xiàn)HN1201株感染PK-15細(xì)胞后病毒增殖速度減緩卻可以達(dá)到相近的病毒滴度,病毒蝕斑變小,與本試驗(yàn)結(jié)果一致。HNQYY2012ΔgE毒力和親本毒相比差異顯著,半數(shù)致死量高于親本毒表明gE基因缺失后PRV毒力下降。gE基因是PRV的主要毒力基因,在病毒的復(fù)制及病毒在細(xì)胞間的傳播過(guò)程中起到關(guān)鍵的作用。研究證明gE基因缺失后,PRV的毒力明顯的降低[20]。將HNQYY2012ΔgE株制備滅活疫苗,并在昆明鼠上進(jìn)行了免疫效力試驗(yàn),結(jié)果表明,免疫滅活疫苗后,小鼠可以抵御105TCID50劑量的親本毒的攻擊,而對(duì)106TCID50劑量攻毒后的保護(hù)率明顯降低。結(jié)果證明所制備的滅活疫苗可以對(duì)小鼠產(chǎn)生有效的保護(hù)。HNQYY2012ΔgE株滅活疫苗免疫動(dòng)物,取得的血清中不含gE抗體,可用于鑒別疫苗免疫動(dòng)物和野毒感染動(dòng)物,故本試驗(yàn)獲得的毒株可作為PRV的候選疫苗株使用。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫(huà)刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂(lè)園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
