miR-122-5p靶向PPP2R2A在嬰兒膽道閉鎖中的作用機制研究①
2020-09-29 07:24:48趙成基李斌德王文赟馬仲福曾永娟胡繼科
中國免疫學雜志 2020年16期
趙成基 李斌德 李 剛 王文赟 馬仲福 曾永娟 胡繼科
(蘭州大學第二醫院小兒外科,蘭州 730000)
膽道閉鎖(biliary atresia,BA)是一種僅見于嬰兒的膽管閉塞性疾病,其特征是膽道進行性梗阻、肝纖維化和炎癥,若BA患兒前期接受積極治療則可以存活至成年[1]。統計顯示,BA的發病有明顯的地區與種族差異,相較于北美和西歐,其在亞洲的發病率明顯較高[2,3]。在臨床上,肝移植仍然是提高BA患兒存活率的主要治療手段,但至今為止BA的發病機制尚不明確,增加了其治療的難度[4]。MicroRNA是內源性的小分子非編碼RNA,可以通過與靶基因的3′非翻譯區(3′-UTR)結合調節靶基因的翻譯和表達,從而參與多種生物學過程[5-7]。Peng等[8]在2016年的報道中表明,miR-122-5p在BA患兒血清中的表達顯著上調,且Gao等[9]的研究顯示miR-122-5p參與了肝癌細胞的增殖、遷移和侵襲。因此,本研究通過構建細胞模型,在體外探究miR-122-5p在BA中的作用機制,為后續的體內實驗及臨床治療提供理論基礎。
1 資料與方法
1.1資料
1.1.1臨床資料 收集2018年1月至2018年12月間到本院就診并診斷為的BA的10例患兒血清,選取同期在本院診斷為非BA的9例患兒血清,基本信息見表1。所有參與本研究的患兒均征得監護人同意且通過我院倫理委員會的批準。
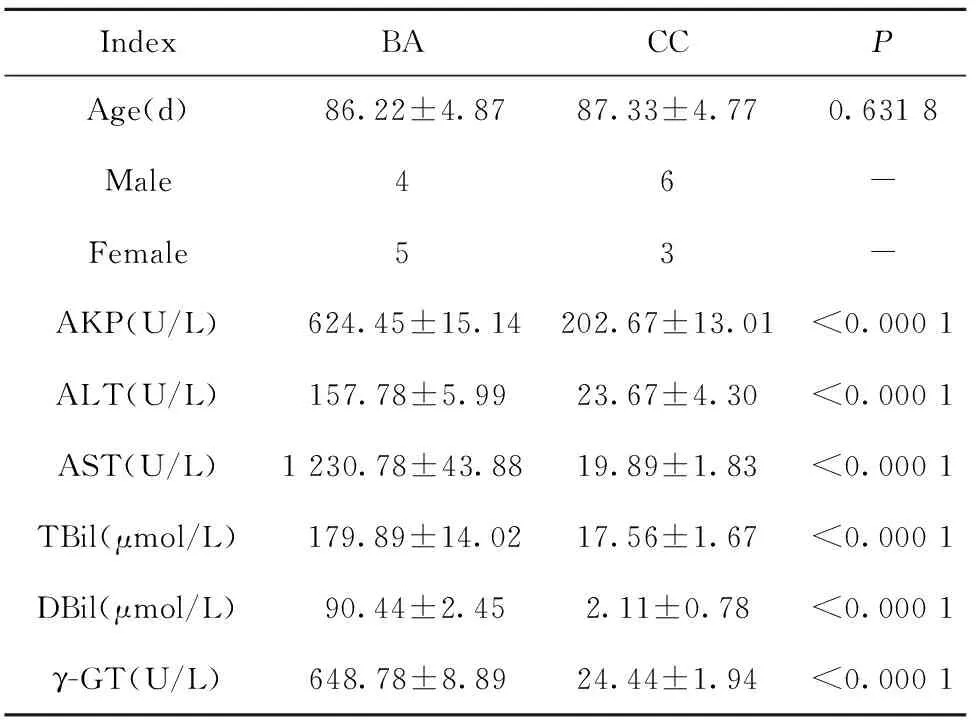
表1 BA及CC患兒臨床信息Tab.1 Clinical information in infants with BA and CC
1.1.2細胞與試劑 人源肝星形細胞LX-2購自武漢益普生物科技有限公司,貨號YCL-0560。胎牛血清和LipofectamineTM2000 Reagent購自Invitrogen公司;TRIzol和RNA提取試劑盒購自北京天健生物技術有限公司;SYBR Premix Ex TaqTMⅡ試劑盒、PrimescripTMRT reagent Kit、miRNA PrimescripTMRT reagent Kit購自TaKaRa公司;引物委托Sangon Biotech設計并合成;青霉素和鏈霉素購自上海源葉生物科技有限公司;RPMI1640培養基、CCK-8和DMSO購自Sigma-Aldrich公司;蛋白提取試劑盒購自南京凱基生物公司;Annexin V-PE/7-AAD Apoptosis Kit購自上海鈺博生物科技有限公司。
1.2方法
1.2.1細胞培養與轉染 將LX-2細胞培養于含10%胎牛血清、100 U/L青霉素及100 mg/L鏈霉素的DMEM培養基中,培養箱培養條件為37℃、5% CO2。待細胞長勢穩定后將miR-122-5p mimics、Anti-miR-122-5p及pc-PPP2R2A按實驗分組轉染入細胞中。用LipofectamineTM2000試劑盒按操作指導完成轉染。48 h后收集轉染細胞進行后續實驗。
1.2.2RT-qPCR 使用TRIzol和RNA提取試劑盒提取總RNA。NanoDrop檢測RNA質量和濃度,統一稀釋為500 ng/μl。使用PrimescripTMRT試劑盒和miRNA PrimescripTMRT試劑盒將PPP2R2A mRNA及miR-122-5p的RNA反轉錄為cDNA,然后使用SYBR Premix Ex TaqTMⅡ試劑盒作為qPCR擴增的模板。在LightCycle 96熒光定量PCR儀上進行RT-qPCR:95℃預變性30 s,95℃變性5 s,共40個循環,退火,60℃延伸30 s。GAPDH用作mRNA內參,U6用作miRNAs內參,并使用2-ΔΔCt法計算相對表達量,重復3次。引物序列見表2。
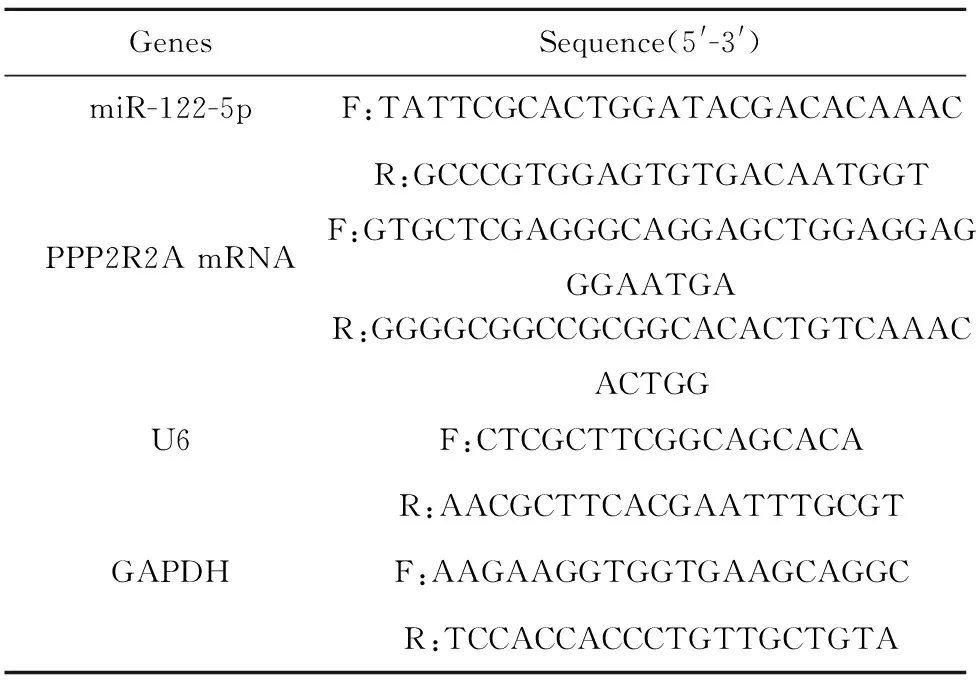
表2 引物序列Tab.2 Primer sequences
1.2.3Western blot 使用RIPA裂解液分離細胞中的蛋白質。將相同濃度的蛋白質混合物在95℃下煮沸10 min。然后將20 μl的混合物(包含30~50 μg樣品)加入到含有10%聚丙烯酰胺凝膠的平板中,電泳分離蛋白質。將蛋白質從凝膠轉移到PVDF膜上,封閉,然后在4℃與一抗(1∶1 000)孵育過夜。樣品用TBST洗滌,然后與二抗(1∶2 000)在室溫下孵育1 h。β-actin或GAPDH用作內參,用Image J測定條帶灰度,重復3次。
1.2.4細胞凋亡檢測 先在不同分組下處理48 h,600 g離心5 min,棄上清并用PBS洗滌細胞1次,各組細胞以1×105個/孔的密度接種到6孔板中培養12 h,然后根據試劑盒的操作流程,使用Annexin V-PE/7-AAD Apoptosis Kit和流式細胞儀檢測不同處理下細胞凋亡情況,重復3次。
1.2.5CCK-8 將細胞以5×103個/孔的密度接種于96孔板,孵育過夜。在經過不同組處理后繼續培養72 h,向各孔中加入CCK-8試劑10 μl,繼續在37℃、5%CO2條件下孵育30 min,使用酶標儀測量各孔在450 nm處的吸光度(OD450),使用CV=(試驗OD450/對照OD450)×100% 計算細胞增殖活性。
1.3統計學處理 采用GraphPad Prism8軟件繪制相關統計圖和進行統計分析。組間數據差異采用t檢驗和單因素方差分析(one-way ANOVA)進行統計分析,P<0.05表示差異具有統計學意義。
2 結果
2.1miR-122-5p在BA患者及CC患者血清中的表達 miR-122-5p在BA患兒血清中的表達量顯著高于CC患兒血清中的表達量(P<0.000 1),提示miR-122-5p可能是嬰兒BA潛在治療靶點。見圖1。

圖1 miR-122-5p在CC及BA患兒血清中的表達量Fig.1 Expressions of miR-122-5p in serum of children with CC and BA
2.2miR-122-5p對LX-2細胞的影響 轉染Anti-miR-122-5p或miR-122-5p mimics入LX-2細胞內miR-122-5p的表達量均有顯著變化(P<0.05)。相較于對照組LX-2細胞,轉染Anti-miR-122-5p顯著抑制LX-2細胞的增殖活性(P<0.000 1),并促進LX-2細胞的凋亡(P<0.000 1),轉染miR-122-5p mimics顯著促進LX-2細胞的增殖活性并抑制LX-2細胞的凋亡(P<0.000 1)。Western blot實驗結果顯示,轉染Anti-miR-122-5p會顯著抑制p-AKT蛋白和Ki67蛋白的表達(P<0.001);轉染miR-122-5p mimics則得到相反的結果。見圖2。
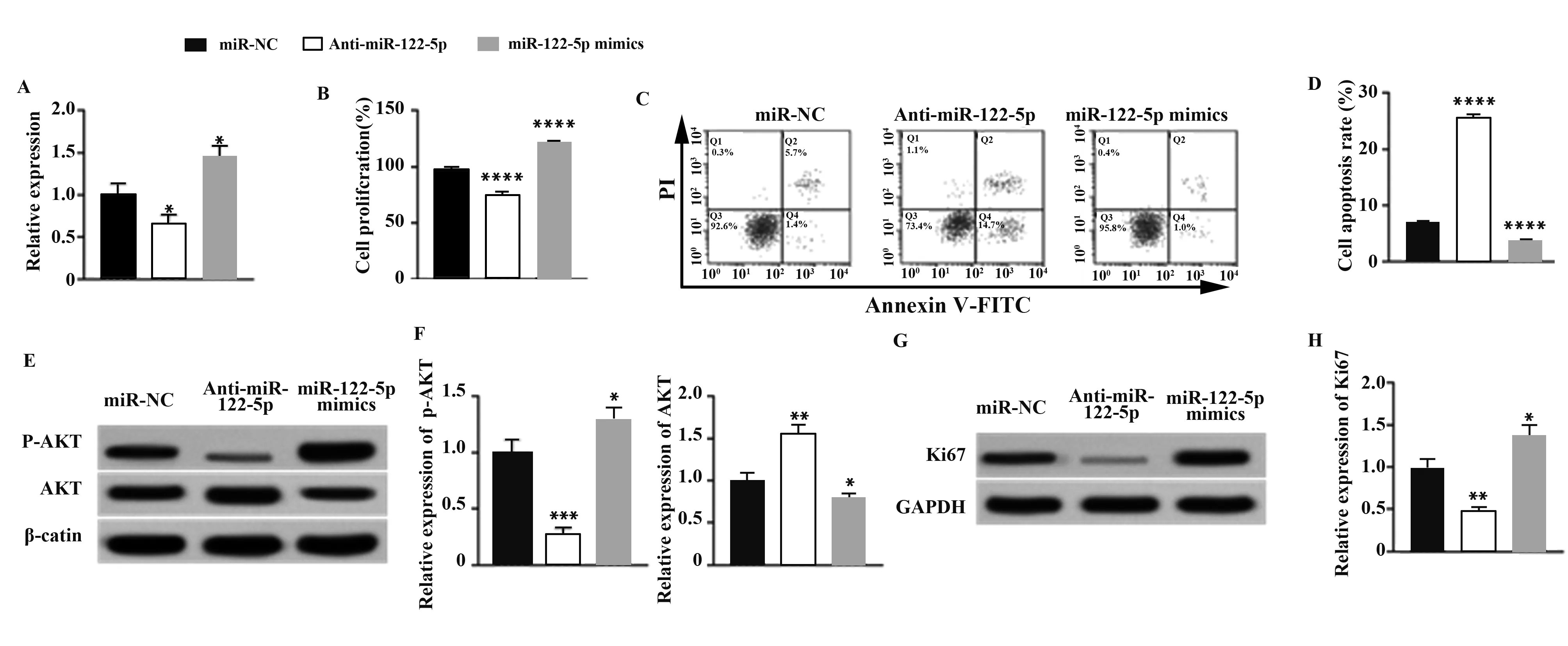
圖2 miR-122-5p對LX-2細胞的影響Fig.2 Effect of miR-122-5p on LX-2 cells
2.3miR-122-5p通過3′-UTR直接靶向PPP2R2A 如圖3A所示,PPP2R2A在其3′-UTR端與miR-122-5p有部分靶向結合位點。雙熒光素酶報告基因實驗結果顯示,過表達miR-122-5P降低了野生型PPP2R2A的熒光素酶活性(P<0.01,圖3B),但對突變型PPP2R2A的熒光素酶活性影響無統計學意義。Western blot實驗結果顯示,過表達miR-122-5p顯著下調LX-2細胞中PPP2R2A蛋白的表達(P<0.01,見圖3C、D)。

圖3 PPP2R2A是miR-122-5p的靶基因Fig.3 PPP2R2A is target gene of miR-122-5p
2.4miR-122-5p靶向PPP2R2A影響LX-2細胞生物學行為 如圖4所示,通過RT-qPCR實驗檢測miR-122-5p與PPP2R2A mRNA的轉染效率。轉染Anti-miR-122-5p或pc-PPP2R2A均顯著上調PPP2R2A蛋白在LX-2細胞中的表達量(P<0.01),同時抑制LX-2細胞的增殖(P<0.001)并促進LX-2細胞的凋亡(P<0.000 1)。共轉染miR-122-5p mimics和pc-PPP2R2A對PPP2R2A蛋白的表達量、LX-2細胞的增殖和凋亡無顯著影響。Western blot實驗結果顯示,轉染Anti-miR-122-5p或pc-PPP2R2A均會顯著抑制p-AKT和Ki67蛋白在LX-2細胞內的表達(P<0.01),并上調AKT蛋白在LX-2細胞內的表達(P<0.01),同時過表達miR-122-5p和PPP2R2A對上述蛋白的表達影響差異無統計學意義。

圖4 miR-122-5p/PPP2R2A對LX-2細胞的影響Fig.4 Effect of miR-122-5p/PPP2R2A on LX-2 cells
3 討論
眾所周知,肝纖維化是BA的顯著特征,然而肝纖維化的形成機制仍然是未知的[10]。研究顯示,LX-2細胞不僅保留了細胞因子的表達且還具有有效轉染等能力,因此非常適用于人類肝纖維化的體外研究[11]。本研究采用LX-2細胞作為體外研究模型,探究miR-122-5p在BA中的作用機制。
研究顯示,miRNA有助于BA的診斷[12,13]。在本研究中通過對在我院就診的BA及CC患兒血清miR-122-5p的表達進行檢測發現,miR-122-5p在BA患兒血清中的表達顯著上調,且抑制miR-122-5p在LX-2細胞中的表達,從而顯著降低LX-2細胞的增殖活性并抑制Ki67蛋白的表達,這說明miR-122-5p在LX-2細胞的增殖中發揮重要作用。PI3-K/AKT信號通路是參與細胞功能的重要途徑之一,如細胞增殖、分化、遷移和細胞內轉運等,且在肝纖維化的過程中也常被激活[14-19]。本研究發現,抑制miR-122-5p在LX-2細胞中的表達可顯著降低AKT蛋白的磷酸化水平,表明miR-122-5p可能參與了AKT蛋白的活化,影響肝纖維化的進程。生物信息學網站預測結果顯示,PPP2R2A是miR-122-5p的下游作用靶點,且雙熒光素酶報告基因實驗證實了這一預測。
PPP2R2A是PPP2A磷酸酶的調控亞基之一,參與了多種細胞功能,其中就包括細胞增殖[20-22]。PPP2A是公認的AKT活性調控因子[23]。有研究報道稱PPP2R2A的異位表達能有效削弱AKT的Ser-473和Thr-308殘基的磷酸化[24]。本研究發現,miR-122-5p靶向并抑制了PPP2R2A蛋白的翻譯,且過表達PPP2R2A在LX-2細胞中能有效抑制AKT蛋白的磷酸化,這表明PPP2R2A可能是miR-122-5p與AKT產生作用的中間信號蛋白。
總之,本研究結果顯示miR-122-5p在BA患兒血清中的表達顯著增加,下調miR-122-5p的表達對LX-2細胞的增殖產生抑制作用并促進LX-2細胞的凋亡,下調miR-122-5p對AKT蛋白的活性也有顯著的抑制作用。綜上,本研究證明miR-122-5p在LX-2細胞內通過調控PPP2R2A的表達影響LX-2細胞的生物學行為,可能成為BA肝纖維化治療的一種新策略。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中老年保健(2021年3期)2021-08-22 06:50:04
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
現代臨床醫學(2021年1期)2021-01-26 00:56:02
科技傳播(2019年22期)2020-01-14 03:06:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
發明與創新(2016年38期)2016-08-22 03:02:52
