氧化型低密度脂蛋白刺激小鼠足細(xì)胞后的氧化應(yīng)激效應(yīng)和機(jī)制研究
2020-10-16 06:16:28李鐸章建娜尤小寒張驥蘇震
浙江醫(yī)學(xué) 2020年18期
李鐸 章建娜 尤小寒 張驥 蘇震
足細(xì)胞損傷在許多腎小球疾病的發(fā)病機(jī)制中起重要作用,腎病綜合征和其他慢性腎臟病患者常出現(xiàn)脂蛋白的代謝異常,腎小球損傷的進(jìn)展過程中常發(fā)現(xiàn)致動脈粥樣硬化脂蛋白的沉積[1]。研究表明,氧化型低密度脂蛋白(oxidized low density lipoprotein,ox-LDL)可與轉(zhuǎn)化生長因子-β(transforming growth factor-β,TGF-β)相互作用,增加內(nèi)皮細(xì)胞和系膜細(xì)胞的損害,引起細(xì)胞外基質(zhì)的聚集,促進(jìn)腎小球硬化的發(fā)生[2-3]。而氧化應(yīng)激被認(rèn)為在糖尿病腎病的發(fā)病機(jī)制中起重要作用。筆者前期研究發(fā)現(xiàn),2型糖尿病腎病的腎小球內(nèi)有不同程度的ox-LDL沉積,且沉積程度與足細(xì)胞裂孔膜上的重要結(jié)構(gòu)成分nephrin的表達(dá)呈負(fù)相關(guān),腎小球內(nèi)超氧化物歧化酶(superoxide dismutase,SOD)的沉積與足細(xì)胞裂孔膜蛋白nephrin的表達(dá)呈正相關(guān)[4]。基于此,本研究擬通過體外實驗來探討足細(xì)胞受到ox-LDL氧化損傷刺激后的效應(yīng)和可能的機(jī)制,現(xiàn)報道如下。
1 材料和方法
1.1 試劑和儀器 LDL、無水CuSO4、足細(xì)胞培養(yǎng)的所有相關(guān)試劑以及鼠抗β-actin抗體均購自美國Sigma公司;PVDF膜(美國BioRad公司);兔抗人Cu2+-ox-LDL抗體(Chemicon International Inc,美國);結(jié)合辣根過氧化物酶的山羊抗兔抗體(美國Amersham Biosciences公司);增強(qiáng)型化學(xué)發(fā)光試劑盒(ECL kit,德國Roche公司);透析袋購自美國Spectrum Medical Industries公司;兔抗鼠nephrin抗體、小鼠抗paxillin抗體購自英國Abcam公司;小鼠抗synaptopodin抗體購自德國Progen公司;熒光標(biāo)記的phalloidin購自美國Molecular Probe公司;TUNEL檢測試劑盒購自德國Roche公司,兔抗絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)多克隆抗體、兔抗p-MAPK多克隆抗體、兔抗糖原合成酶激酶 3β(glycogen synthase kinase-3β,GSK-3β)多克隆抗體、鼠抗 p-GSK-3β(Ser9)單克隆抗體購自美國Cell Signaling公司。
1.2 方法
1.2.1 小鼠足細(xì)胞的培養(yǎng) 條件永生性小鼠腎小球足細(xì)胞來自原代分離培養(yǎng)所得[5]。在33℃、20 U/ml γ-干擾素的培養(yǎng)條件下促進(jìn)足細(xì)胞增殖以獲取足夠的細(xì)胞數(shù)。在37℃、無γ-干擾素的培養(yǎng)條件下,在完全培養(yǎng)基內(nèi)培養(yǎng)14 d至其分化成熟后用于后續(xù)實驗[5]。
1.2.2 ox-LDL的制備和鑒定 用30 ml蒸餾水將48 mg無水CuSO4溶解后用一次性無菌濾器過濾,將LDL稀釋于無菌PBS溶液,終濃度為1 mg/ml,在分子截流量為6 000~8 000道爾頓的透析袋內(nèi)將3 ml LDL置于無菌PBS溶液內(nèi)于4℃透析去除EDTA。參照J(rèn)enkins等[6]的方法,3 ml LDL溶液和3 μl CuSO4溶液混勻后置于37℃恒溫箱內(nèi),在空氣中氧化24 h,成為氧化后的LDL溶液,實驗之前新鮮配制。采用Western blot法鑒定制備的ox-LDL的特異性。
1.2.3 實驗分組 將條件永生性小鼠腎小球足細(xì)胞分別接種于面積為25 cm2(T25)的細(xì)胞培養(yǎng)瓶、包被膠原的載玻片中。在37℃、無γ-干擾素的培養(yǎng)條件下,采用完全培養(yǎng)基培養(yǎng)14 d后分化成熟。成熟的足細(xì)胞分為6組:12 h對照組、12 h LDL 刺激組(終濃度為 50 μg/ml)、12 h ox-LDL 刺激組(終濃度為 50 μg/ml)、24 h 對照組、24 h LDL刺激組、24 h ox-LDL刺激組。單純用無血清培養(yǎng)基培養(yǎng)的足細(xì)胞作為對照組。免疫熒光法和細(xì)胞凋亡檢測培養(yǎng)于載玻片上的細(xì)胞,提取培養(yǎng)瓶內(nèi)細(xì)胞的蛋白做蛋白分析。
1.2.4 nephrin和細(xì)胞骨架蛋白paxillin、F-actin表達(dá)檢測 采用免疫熒光法。將條件永生性小鼠腎小球足細(xì)胞接種于包被膠原的載玻片上,用4℃丙酮固定細(xì)胞后,Tris鹽酸緩沖液沖洗,Triton X-100進(jìn)行細(xì)胞通透后,1% FBS封閉,加入一抗4℃孵育過夜,陰性對照組以TBS代替。加入二抗于室溫下孵育1 h,二抗分別為相應(yīng)的山羊抗小鼠和山羊抗兔抗體,DAPI(1:1 000)染色細(xì)胞核。TBS沖洗后封片。在400倍的熒光顯微鏡下觀察并隨機(jī)選取10個視野,對視野內(nèi)的細(xì)胞總數(shù)和陽性細(xì)胞數(shù)分別計數(shù),計算兩者的比值即為陽性率。
1.2.5 細(xì)胞凋亡情況檢測 采用脫氧核糖核苷酸末端轉(zhuǎn)移酶介導(dǎo)的缺口末端標(biāo)記(terminal deoxynucleotidyl transferase-mediated UTP nick-end labeling,TUNEL)法。取24 h各實驗組接種于包被膠原的載玻片上的足細(xì)胞,PBS清洗后,室溫下固定細(xì)胞1 h,3% H2O2-甲醇溶液封閉10 min,在冰上于0.1% Triton X-100及0.1%檸檬酸鈉溶液中通透蓋玻片2 min,然后蓋上玻片。按照試劑盒說明書進(jìn)行操作,實驗重復(fù)3次,光鏡下每組至少觀察200個細(xì)胞,計數(shù)陽性細(xì)胞數(shù)并計算百分率。
1.2.6 細(xì)胞SOD活性檢測 采用細(xì)胞超氧基團(tuán)介導(dǎo)的煙酰胺腺嘌呤二核苷酸(nicotinaminde adenine dinucleotide,NADH)氧化率來反映SOD活性,若SOD活性高,則NADH的氧化率下降。根據(jù)上述原理,參照Paoletti等[7]的方法,提取足細(xì)胞細(xì)胞質(zhì)SOD。將12 h對照組及ox-LDL刺激組、24 h對照組及ox-LDL刺激組的細(xì)胞分別用冰的無菌PBS清洗后,加入250 μl 25 mmol/L的三乙醇胺-二乙醇胺(Tea-Dea)緩沖液后,收集細(xì)胞裂解液,4℃離心后取上清液。分光光度法檢測提取的足細(xì)胞細(xì)胞質(zhì)SOD活性。依次加入800 μl Tea-Dea緩沖液,5 μl 7.5 mmol/L NADH 溶液,25 μl EDTA/MnCl2,100 μl稀釋后的樣品至水晶比色杯后,混合均勻,空白對照組以100 μl Tea-Dea緩沖液替代樣品。以空水晶比色杯調(diào)零點,分別記錄各組和空白對照組在340 nm波長下的吸收值,以此作為基線值,同時向各組分別加入 100 μl 10 mmol/L β-巰基乙醇,觀察并記錄吸光度變化值,共觀察24 min,每8 min記錄1次。重復(fù)實驗3次,第8分鐘相對基線值的吸光度變化值記為ΔA340。抑制率為各組ΔA340與當(dāng)次實驗中空白對照組ΔA340的比率,通過抑制率來計算SOD的活性。
1.2.7 細(xì)胞 nephrin及 MAPK、phosphor-p44/42MAPK(p-MAPK)、GSK-3β、phosphor-GSK-32(p-GSK-3β)表達(dá)檢測 采用Western blot法。各實驗組細(xì)胞加入蛋白提取液,于冰上裂解5 min后收集蛋白提取物并定量。變性、電泳、轉(zhuǎn)膜封閉后,加入一抗 nephrin、GSK-3β、p-GSK-3β、MAPK、p-MAPK;使用相應(yīng)的二抗室溫孵育1 h后,用ECL試劑盒發(fā)光、曝光、沖片。膠片掃描后,采用Image J軟件對條帶進(jìn)行分析。
1.3 統(tǒng)計學(xué)處理 采用SPSS19.0統(tǒng)計軟件。計量資料以表示,多組比較采用單因素方差分析,兩兩比較采用LSD-t檢驗。P<0.05為差異有統(tǒng)計學(xué)意義。
2 結(jié)果
2.1 ox-LDL特異性的鑒定結(jié)果 Western blot結(jié)果顯示,抗Cu2+-ox-LDL多克隆抗體與陰性對照、LDL、Cu2+-ox-LDL共同孵育后,僅Cu2+-ox-LDL泳道觀察到1條160 kd左右的條帶,證實Cu2+-ox-LDL制備成功,其特性不同于LDL。兔來源的免疫球蛋白IgG與Cu2+-ox-LDL孵育后,膜上也未觀察到任何條帶,表明該抗體具有特異性(圖1),進(jìn)一步證實制備的ox-LDL具有特異性。
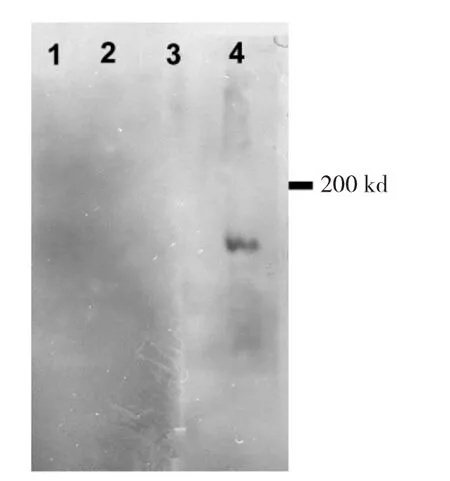
圖1 ox-LDL特異性鑒定電泳圖(1:緩沖液,一抗為兔抗Cu2+-ox-LDL;2:Cu2+-ox-LDL,一抗為兔的 IgG;3:LDL,一抗為兔抗Cu2+-ox-LDL;4:配制的 Cu2+-ox-LDL,一抗為兔抗 Cu2+-ox-LDL)
2.2 ox-LDL刺激組、LDL刺激組、對照組足細(xì)胞nephrin和paxillin、F-actin表達(dá)比較 間接免疫熒光染色后,觀察到12 h對照組、LDL刺激組和ox-LDL刺激組的nephrin表達(dá)強(qiáng)度及部位未見明顯變化(圖2,見插頁)。24 h對照組的nephrin在細(xì)胞膜、足突和細(xì)胞的足突間連接上仍有表達(dá),與12 h對照組相比nephrin強(qiáng)度未見明顯變化,LDL刺激組在足突上仍可觀察到nephrin少量表達(dá),而ox-LDL刺激組在細(xì)胞膜、足突上均未觀察到nephrin表達(dá)(圖2,見插頁)。表明ox-LDL刺激小鼠腎小球足細(xì)胞24 h后導(dǎo)致足細(xì)胞正常表達(dá)部位nephrin的喪失。
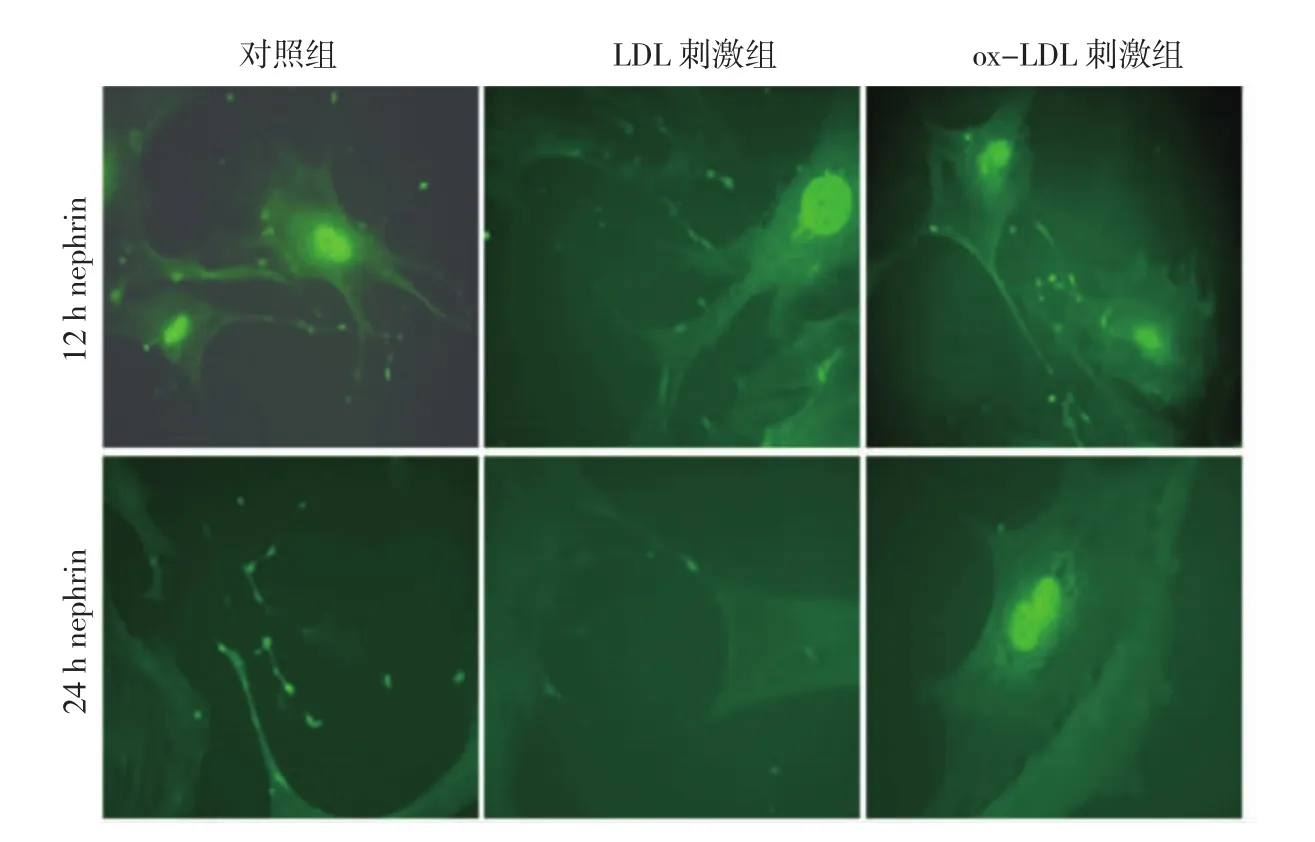
圖2 氧化型低密度脂蛋白(ox-LDL)刺激組、LDL刺激組、對照組足細(xì)胞nephrin表達(dá)比較(免疫熒光染色,×400)
足細(xì)胞骨架蛋白的觀察結(jié)果顯示,24 h對照組的paxillin在細(xì)胞質(zhì)和足突均有表達(dá),LDL刺激后足細(xì)胞的paxillin強(qiáng)度和分布均未見明顯改變,ox-LDL刺激24 h后,足突上paxillin的表達(dá)消失,paxillin僅細(xì)胞質(zhì)內(nèi)有表達(dá)(圖3,見插頁)。異硫氰熒光素標(biāo)記的phalloidin可染色F-actin。24 h對照組的F-actin沿細(xì)胞輪廓和足突有序分布;24 h LDL刺激組的染色強(qiáng)度降低,但F-actin沿細(xì)胞輪廓分布仍可見;經(jīng)ox-LDL刺激24 h后,未見足細(xì)胞中F-actin的正常分布,F(xiàn)-actin重新分布,結(jié)構(gòu)紊亂(圖3,見插頁)。提示此時足突的骨架蛋白發(fā)生重構(gòu)。
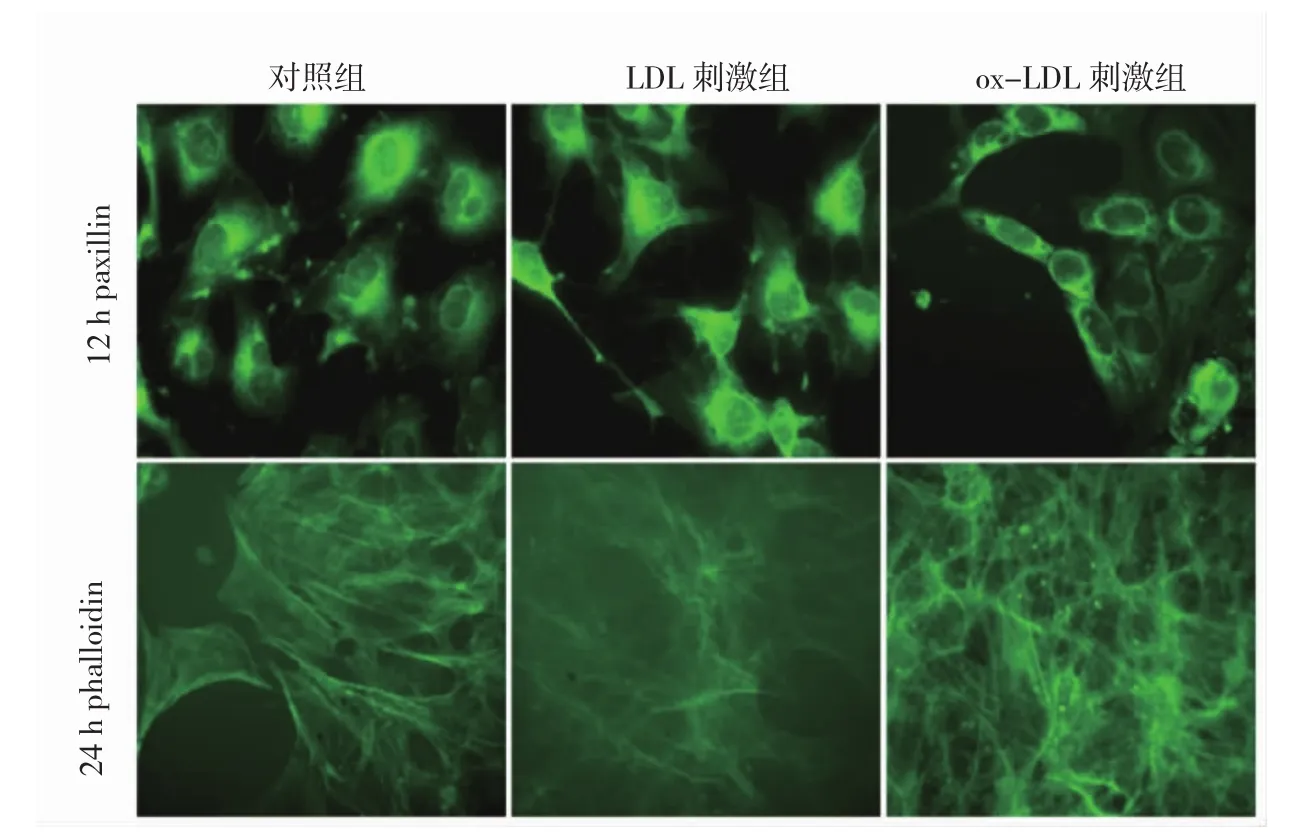
圖3 氧化型低密度脂蛋白(ox-LDL)刺激組、LDL刺激組、對照組足細(xì)胞骨架蛋白paxillin、F-actin表達(dá)比較(免疫熒光染色,×250)
2.3 24hox-LDL刺激組、LDL刺激組、對照組足細(xì)胞凋亡情況比較 TUNEL檢測顯示,無論24 h對照組,還是24 h LDL和ox-LDL刺激組,均未觀察到足細(xì)胞核著色(即發(fā)生凋亡)(圖4,見插頁),陽性對照觀察到細(xì)胞核染色陽性。說明50 μg/ml的LDL和ox-LDL分別刺激24 h,均不足以完全激活足細(xì)胞凋亡通路,只引起足細(xì)胞nephrin表達(dá)下降等損傷性改變。

圖4 24h ox-LDL刺激組、LDL刺激組、對照組足細(xì)胞凋亡情況比較(TUNEL法,×100)
2.4 ox-LDL刺激組和對照組足細(xì)胞SOD活性比較 12 h ox-LDL刺激組、12 h對照組、24 h ox-LDL刺激組、24 h對照組比較,12 h ox-LDL刺激組的SOD活性最高,NADH的氧化抑制率為(22.77±5.29)%,和其它3組相比均有統(tǒng)計學(xué)差異(均P<0.05),24 h ox-LDL刺激組的SOD活性較12 h ox-LDL刺激組下降,抑制率為(14.33±5.36)%,但差異無統(tǒng)計學(xué)意義(P >0.05)(圖5),表明足細(xì)胞受到氧化刺激時會產(chǎn)生抗氧化能力,但若氧化刺激持續(xù)存在,抗氧化能力亦會逐漸下降。
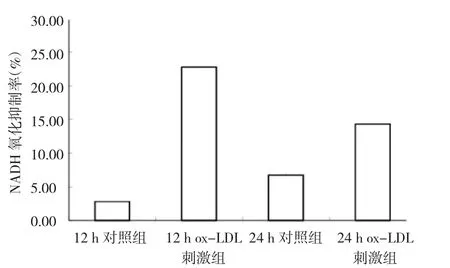
圖5 氧化型低密度脂蛋白(ox-LDL)刺激組和對照組足細(xì)胞超氧化物岐化酶(SOD)活性比較
2.5 ox-LDL刺激組、LDL刺激組、對照組足細(xì)胞nephrin及MAPK和GSK-3β信號通路情況比較 與12 h對照組相比,12 h ox-LDL刺激組、LDL刺激組足細(xì)胞nephrin表達(dá)水平無統(tǒng)計學(xué)差異(均P>0.05),提示12 h后的足細(xì)胞的nephrin表達(dá)暫未受到刺激的影響。p-MAPK/MAPK、p-GSK-3β/GSK-3β兩組信號通路中,分別進(jìn)行比值比較后均未見統(tǒng)計學(xué)差異(均P>0.05)(圖6-7),說明盡管足細(xì)胞受到不同刺激,但是12 h后的MAPK、GSK-3β的活性無明顯變化。
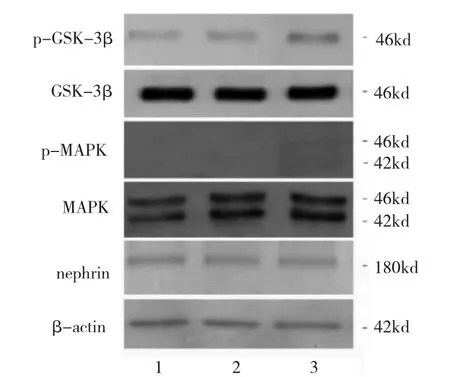
圖6 12 h氧化型低密度脂蛋白(ox-LDL)刺激組、LDL刺激組、對照組足細(xì)胞nephrin及絲裂原活化蛋白激酶(MAPK)和糖原合成酶激酶3β(GSK-3β)信號通路蛋白表達(dá)電泳圖(1:對照組;2:12 h LDL刺激組;3:12 h ox-LDL刺激組)
24 h LDL刺激組與24 h對照組相比,nephrin表達(dá)水平略有下降,而24 h ox-LDL刺激組和其它兩組相比,nephrin表達(dá)水平降低(P<0.05),表明足細(xì)胞受到ox-LDL造成的氧化刺激24 h可引起嚴(yán)重?fù)p傷,導(dǎo)致其正常的裂孔膜蛋白表達(dá)下降。同樣,與24 h對照組相比,24 h LDL刺激組的p-MAPK和MAPK比值、p-GSK-3β和GSK-3β比值均無統(tǒng)計學(xué)差異(均P>0.05);與24 h LDL刺激組和24 h對照組相比,24 h ox-LDL刺激組 p-MAPK和MAPK比值、p-GSK-3β和GSK-3β比值均明顯增高(均P<0.05)(圖8-9),MAPK和GSK-3β信號通路在ox-LDL刺激24 h后被激活。
3 討論
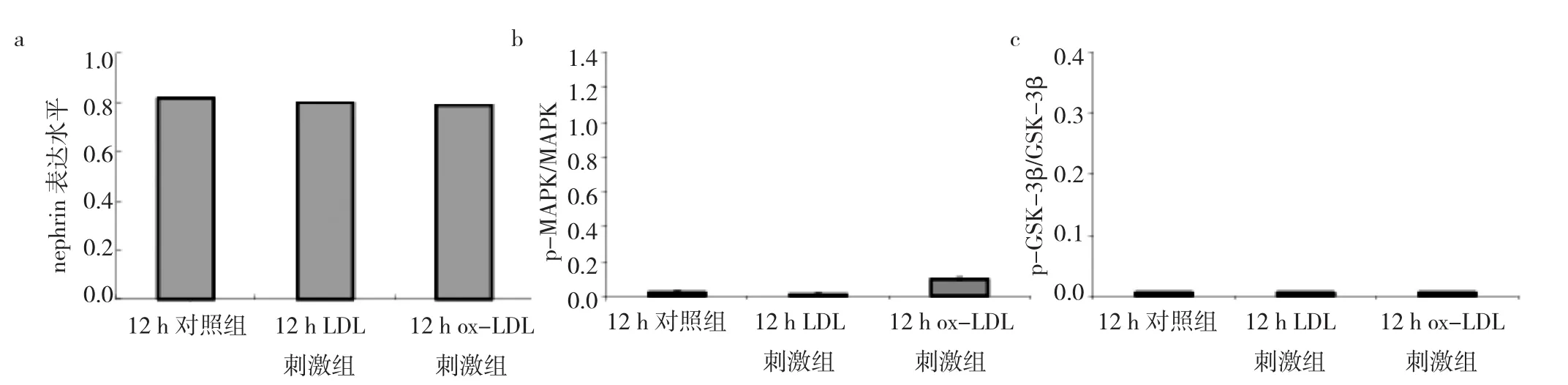
圖7 12 h氧化型低密度脂蛋白(ox-LDL)刺激組、LDL刺激組、對照組足細(xì)胞nephrin及絲裂原活化蛋白激酶(MAPK)和糖原合成激酶3β(GSK-3β)信號通路蛋白表達(dá)水平比較(a:nephrin表達(dá)水平比較;b:p-MAPK和MAPK比值;c:p-GSK-3β和GSK-3β比值)
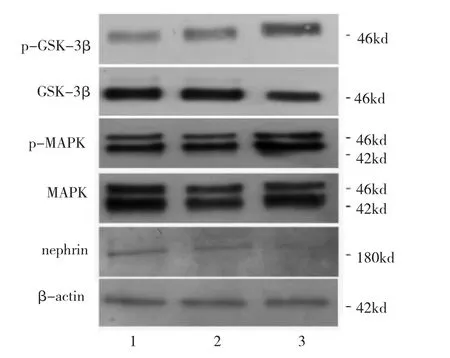
圖8 24 h氧化型低密度脂蛋白(ox-LDL)刺激組、LDL刺激組、對照組足細(xì)胞nephrin及絲裂原活化蛋白激酶(MAPK)和糖原合成酶激酶 3β(GSK-3β)信號通路蛋白表達(dá)電泳圖(1:對照組;2:24 h LDL刺激組;3:24 h ox-LDL刺激組)
在過量自由基和其他致氧因素作用下,沉積在血管壁上的血漿LDL的多價不飽和脂肪酸發(fā)生過氧化,產(chǎn)生多種活性醛類,LDL載脂蛋白B中的賴氨酸殘基與活性醛結(jié)合發(fā)生化學(xué)修飾,形成的產(chǎn)物為ox-LDL。高脂血癥可以使ox-LDL在腎臟沉積、炎癥細(xì)胞浸潤、腎臟固有細(xì)胞增生和損傷、細(xì)胞外基質(zhì)積聚及泡沫細(xì)胞形成,直接或間接地導(dǎo)致腎小球硬化。因此,ox-LDL是腎小球硬化進(jìn)展的主要危險因素之一。近年研究發(fā)現(xiàn),足細(xì)胞有攝取ox-LDL的受體[8],對足細(xì)胞的損傷可能通過RhoA激酶1介導(dǎo)的脂質(zhì)自噬障礙[9]。筆者在前期研究中觀察到2型糖尿病腎病的腎小球內(nèi)存在氧化程度較重的ox-LDL沉積,且隨著ox-LDL在腎小球內(nèi)的沉積增加,足細(xì)胞的nephrin表達(dá)呈下降趨勢。在有較多SOD-1表達(dá)的腎小球內(nèi),nephrin的表達(dá)相對增多。因此,本研究利用體外培養(yǎng)成熟的足細(xì)胞來觀察ox-LDL作用足細(xì)胞后造成的損傷及足細(xì)胞抗氧化的反應(yīng),探討可能的機(jī)制。
免疫熒光和Western blot的結(jié)果表明,與LDL刺激組和對照組相比,50 μg/ml的ox-LDL刺激足細(xì)胞12 h后,nephrin的蛋白表達(dá)強(qiáng)度、在足細(xì)胞上的分布,骨架蛋白paxillin和F-actin的活性,MAPK、GSK-3β的活性都未見顯著變化,說明在50 μg/ml濃度的LDL、ox-LDL刺激下,12 h可能不足以造成足細(xì)胞損傷。以相同濃度的ox-LDL、LDL分別刺激足細(xì)胞24 h后,LDL刺激組nephrin表達(dá)有所下調(diào),但骨架蛋白的分布未見明顯改變,p-MAPK/MAPK、p-GSK-3β/GSK-3β 均略有增加,但與對照組相比未見統(tǒng)計學(xué)差異;ox-LDL刺激組的上述指標(biāo)均發(fā)生了顯著變化,nephrin表達(dá)顯著降低,足突上骨架蛋白paxillin的表達(dá)消失,足細(xì)胞中phalloidin染色的F-actin發(fā)生了重構(gòu),同時p-MAPK/MAPK、p-GSK-3β/GSK-3β兩組比值顯著增加,提示這兩條通路的激活可能參與足細(xì)胞的氧化損傷。TUNEL檢測結(jié)果顯示24 h未見凋亡細(xì)胞,證實24 h的50 μg/ml ox-LDL刺激尚不足以引起細(xì)胞凋亡,此時足細(xì)胞的損傷作用可能主要表現(xiàn)為裂孔膜復(fù)合體分子構(gòu)成的變化和足突結(jié)構(gòu)的重排。而使用80 μg/ml的ox-LDL作用于人的足細(xì)胞則可導(dǎo)致足細(xì)胞的凋亡和細(xì)胞內(nèi)脂質(zhì)的蓄積[10]。
MAPK 家族包括 c-Jun N-terminal kinase(JNK)、細(xì)胞外信號調(diào)節(jié)激酶(extracellular signal-regulated kinase,ERK)和p38,是多種細(xì)胞外刺激(如生長因子)和周圍環(huán)境刺激改變細(xì)胞功能的通路[11]。MAPK信號通路涉及多種磷酸化級聯(lián)反應(yīng),每一種或者單獨介導(dǎo)不同的信號事件或者與其它通路相連。本實驗所使用的特異性抗體針對的是MAPK通路中ERK1/ERK2,因此觀察的MAPK通路是ERK1/ERK2通路。而Hu等[12]的研究發(fā)現(xiàn)ox-LDL通過激活FAK/p38途徑引起足細(xì)胞損傷。ERK1/p-ERK2和ERK1/ERK2的比值在LDL刺激組無明顯變化,而在ox-LDL刺激組增加明顯,表明ox-LDL具有激活ERK通路的作用。
GSK-3最早系調(diào)節(jié)糖原合成酶和磷酸化的激酶而被發(fā)現(xiàn)。有兩種類型:GSK-3α和GSK-3β,兩者結(jié)構(gòu)高度相似,GSK-3可調(diào)控糖原代謝,并可調(diào)節(jié)多種底物的磷酸化。它的活性可被GSK-3α(ser21位點)和GSK-3β(ser9位點)磷酸化所抑制。本研究發(fā)現(xiàn)足細(xì)胞受ox-LDL刺激24 h后磷酸化的GSK-3β較其它兩組增強(qiáng),GSK-3β活性受到抑制。研究發(fā)現(xiàn),GSK-3β的活性與Fyn酪氨酸激酶的活性相關(guān)。Fyn的活性對于nephrin細(xì)胞內(nèi)段酪氨酸磷酸化具有重要調(diào)節(jié)作用。筆者推測,足細(xì)胞受到50 μg/ml的ox-LDL刺激24 h后,p-GSK-3β增加,造成活性下降,并可能導(dǎo)致Fyn的活性也下降,進(jìn)一步使nephrin磷酸化減少,引起nephrin表達(dá)下調(diào)。但這一推測仍需進(jìn)一步的深入研究才可能明確。
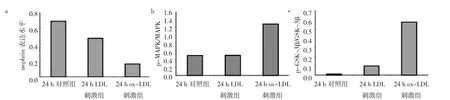
圖9 24 h氧化型低密度脂蛋白(ox-LDL)刺激組、LDL刺激組、對照組足細(xì)胞nephrin及絲裂原活化蛋白激酶(MAPK)和糖原合成酶激酶3β(GSK-3β)信號通路蛋白表達(dá)水平比較(a:nephrin表達(dá)水平比較;b:p-MAPK和MAPK比值;c:p-GSK-3β和GSK-3β比值)
細(xì)胞骨架是維持真核細(xì)胞生命活動的重要組成成分。細(xì)胞維持正常的形態(tài)、保持細(xì)胞彈性,在細(xì)胞黏著、識別和相互的“交流”中,細(xì)胞骨架的正常分布具有重要作用。ox-LDL刺激足細(xì)胞24 h后,不同細(xì)胞骨架蛋白的分布也發(fā)生異常改變,這可能是足細(xì)胞損傷后足突結(jié)構(gòu)發(fā)生改變、濾過屏障完整性受損的重要原因之一。Paxillin是分子量為68 kd的細(xì)胞骨架蛋白,在調(diào)節(jié)細(xì)胞的伸展和移行中起到重要作用。Paxillin的磷酸化依賴于GSK-3和ERK的雙重調(diào)節(jié)作用,而磷酸化對它的生物活性的調(diào)節(jié)至關(guān)重要[13]。本研究發(fā)現(xiàn)的ERK1/ERK2通路激活和GSK-3β活性下降,推測可能是造成paxillin磷酸化改變以及細(xì)胞骨架蛋白重構(gòu)的原因。
本研究還發(fā)現(xiàn),暴露于ox-LDL刺激12 h后,足細(xì)胞的SOD活性會明顯增加,高出同樣條件對照組19.94%,而當(dāng)該刺激持續(xù)24 h后,SOD活性僅比對照組高7.3%。結(jié)合12、24 h時這兩個時間點的nephrin表達(dá)變化,再次肯定了筆者前期在2型糖尿病腎病患者腎活檢中發(fā)現(xiàn)的現(xiàn)象,即SOD活性增加可保護(hù)足細(xì)胞免受氧化刺激損傷;若氧化刺激持續(xù)存在造成SOD耗竭,則會打破抗氧化和氧化之間的平衡,最終導(dǎo)致足細(xì)胞受到損傷,并且引發(fā)裂孔膜蛋白喪失、細(xì)胞骨架蛋白重構(gòu)等一系列嚴(yán)重病理事件。
總之,本研究結(jié)果顯示,體外培養(yǎng)的足細(xì)胞受到ox-LDL的氧化刺激后,早期可引起抗氧化物質(zhì)SOD活性明顯增加,足細(xì)胞并不出現(xiàn)損傷,而隨著氧化刺激的持續(xù)存在,SOD的活性僅輕度升高,足細(xì)胞出現(xiàn)損傷,MAPK、GSK-3β兩條信號通路的激活可能參與導(dǎo)致nephrin表達(dá)下調(diào)和細(xì)胞骨架蛋白重構(gòu)這些病理事件的發(fā)生。
