急性砷暴露對小鼠腎臟免疫功能的影響
2020-10-22 02:16:24許何麗徐國偉毛亞萍竇文靜王主段曉旭
沈陽醫學院學報 2020年5期
許何麗, 徐國偉, 毛亞萍, 竇文靜, 王主, 段曉旭*
(1. 沈陽醫學院公共衛生學院預防醫學專業2016 級, 遼寧 沈陽110034; 2. 中國醫科大學公共衛生學院環境與慢病研究中心; 3. 沈陽醫學院公共衛生學院毒理學教研室; 4. 沈陽醫學院公共衛生實驗教學中心)
砷是地殼中普遍存在的元素之一, 長期砷暴露嚴重威脅人類健康, 可導致皮膚癌、 膀胱癌、肺癌等多器官損害。 除了致癌性, 流行病學和實驗研究還發現砷具有一定的免疫調節效應[1-2], 這正成為砷研究領域一個新的熱點問題。 無機砷可以誘導巨噬細胞分化和吞噬能力減弱、 朗格漢斯細胞和樹突狀細胞的數目明顯減少以及Th17 細胞特異性轉錄因子維甲酸相關孤核受體γt (retinoidrelated orphan receptor gammat, RoRγt) 及T 細胞增殖能力下降[3]。 除脾、 胸腺這些免疫器官外,越來越多的研究提示機體內一些非免疫器官中也含有巨噬細胞、 樹突狀細胞、 淋巴細胞等免疫細胞, 并且這些免疫細胞可以參與機體的免疫調節,從而參與一些免疫和非免疫性疾病的發生發展。如腎臟中含有存在由樹突狀細胞和巨噬細胞構成的腎單核吞噬細胞系統及部分CD4+/CD8+組織常駐記憶T 細胞(tissue-resident memory T cell, Trm細胞) 和Treg 細胞等, 這些免疫細胞組成的免疫微環境可以調節機體的免疫應答及腎臟的防御功能[4-5]。 腎臟是砷排泄、 蓄積和發揮毒作用的主要靶器官, 砷暴露增加大鼠腎臟中TNF-α、 NF-κB和IL-1β 等的表達[6]。 這些結果提示砷引起腎損傷的過程中或多或少的會捕捉到免疫反應的“蹤跡”, 但目前關于砷對腎臟的免疫學相關研究尚不多見。 本研究采用整體動物學模型, 探討急性砷暴露對小鼠腎臟內抗原吞噬、 趨化黏附和抗原提呈功能的影響, 為砷所致腎損傷提供新的思路和靶點。
1 材料與方法
1.1 實驗動物 SPF 級C57BL/6 雌性小鼠40 只,體重18~22 g, 由北京維通利華公司提供, 實驗動物許可證號11400700063049, 飼養于沈陽醫學院實驗動物中心。 本實驗遵循沈陽醫學院有關實驗動物管理和使用以及國家《關于動物倫理與福利的作者指南共識》 的規定。
1.2 儀器與試劑 超高速低溫離心機(美國Heal Force 公司), Infinite M200 型微孔板發光檢測儀(瑞士Tecan 公司), Nano Drop ND-2000 分光光度計(美國Thermo Fisher Scientific 公司), ABI 7500實時定量PCR 儀 (美國ABI 公司), 亞砷酸鈉(NaAsO2) (分析純, 美國Sigma 公司); Trizol 試劑盒和SYBR Premix Ex Taq II Real-time PCR 試劑盒(日本TaKaRa 公司), 尿素氮(BUN)、 肌酐(Cr)、 酸性磷酸酶(ACP) 測定試劑盒(南京建成生物工程研究所)。
1.3 方法
1.3.1 砷溶液配置 將2.5 g NaAsO2粉末, 溶于100 ml 純凈水中, 配置成濃度為25 g/L 的砷儲備液, 逐步稀釋為0.125、 0.25 和0.5 g/L 的砷溶液用于實驗。
1.3.2 模型建立 將小鼠隨機分為4 組, 分別為對照組、 2.5、 5 和10 mg/kg NaAsO2暴露組, 各10 只。 一次性灌胃方式染毒, 染毒時間為24 h,對照組給予生理鹽水。 記錄小鼠染毒前后體重變化。 染毒終點眼球取血用于生化指標測定, 處死解剖, 摘取完整的腎臟, 記錄小鼠腎臟重量。
1.3.3 生化檢測 取各組小鼠外周血并分離出血清, 按照試劑盒說明書檢測血清中BUN、 Cr 和ACP 水平, 檢測腎組織勻漿中ACP 水平。
1.3.4 Real-time PCR 法檢測腎臟免疫功能 將收集的腎組織充分勻漿, 加入Trizol 溶液中, 提取組織中的總RNA, 反轉錄合成cDNA 后, 測定腎臟趨化黏附功能分子(Ccr5、Ccr7、Icam1) 和抗原提 呈 功 能 相 關 分 子 (MhcⅡ、Cd80、Cd86、Cd40) 的表達, 嚴格按說明書配制反應體系并在相應條件下進行擴增。 反應結束后, 根據溶解曲線判斷待擴增基因的特異性, 采用2-△△Ct值表示各目的基因的mRNA 相對表達量, 各引物序列見表1。

表1 引物序列

續表1
1.4 統計學方法 采用SPSS 24.0 軟件對數據進行統計學分析, 計量資料以均數±標準差表示, 組間比較采用單因素方差分析, 兩兩比較采用LSD-t法(滿足方差齊性時) 或Dunnett’ T3 法(不滿足方差齊性時)。P<0.05 為差異有統計學意義。
2 結果
2.1 一般情況比較 實驗期間, 各組小鼠生長良好, 活動靈敏, 大便成型, 毛發順齊, 均存活且未見明顯的系統毒性。 各組小鼠染毒前后體重無明顯變化, 與對照組比較差異無統計學意義(P>0.05), 砷暴露組小鼠的腎臟重量及臟器系數均無明顯變化(P>0.05), 見圖1。
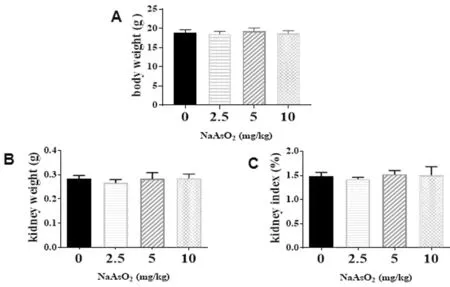
圖1 各組小鼠一般情況比較
2.2 血清BUN 和Cr 水平比較 與對照組比較,砷暴露組小鼠血清BUN 水平明顯增加(P<0.05),見圖2A; 而4 組小鼠血清Cr 水平比較差異無統計學意義(P>0.05), 見圖2B。
2.3 砷暴露對小鼠腎臟抗原吞噬功能的影響 與對照組比較, 血清ACP 水平降低(P<0.05), 見圖3A; 腎組織中ACP 水平升高(P<0.05), 見圖3B。
2.4 砷暴露對小鼠腎臟趨化黏附功能的影響 砷暴露組小鼠腎組織中Ccr5、Ccr7的mRNA 水平隨著染毒劑量的增加逐漸下降, 10 mg/kg 砷暴露組與其它組比較差異均有統計學意義(P<0.05),見圖4A、 B; 與對照組比較, 砷暴露組Icam1的mRNA 水平均出現不同程度的下降, 5 mg/kg 砷暴露組與2.5 mg/kg 和10 mg/kg 砷暴露組比較差異均有統計學意義(P<0.05), 見圖4C。

圖2 對照組和砷暴露組小鼠血清BUN 和Cr 水平比較

圖3 對照組和砷暴露組小鼠血清和腎組織ACP 水平比較
2.5 砷暴露對小鼠腎臟抗原提呈功能的影響 與對照組、 2.5 mg/kg 砷暴露組比較, 5 和10 mg/kg砷暴露組腎組織中MhcⅡ的mRNA 水平隨著染毒劑量的增加逐漸下降, 5 和10 mg/kg 砷暴露組比較差異有統計學意義(P<0.05), 見圖5A; 與對照組比較, 砷暴露組Cd80與Cd86的mRNA 表達水平明顯下降(P<0.05), 與2.5 mg/kg 砷暴露組比較, 5 和10 mg/kg 砷暴露組Cd80的mRNA 表達水平明顯升高, 見圖5B、 C; 而Cd40的mRNA 水平與對照組相比無明顯變化(P>0.05), 見圖5D。

圖4 對照組和砷暴露組小鼠腎臟組織中Ccr5、Ccr7 和Icam1 的mRNA 表達水平比較
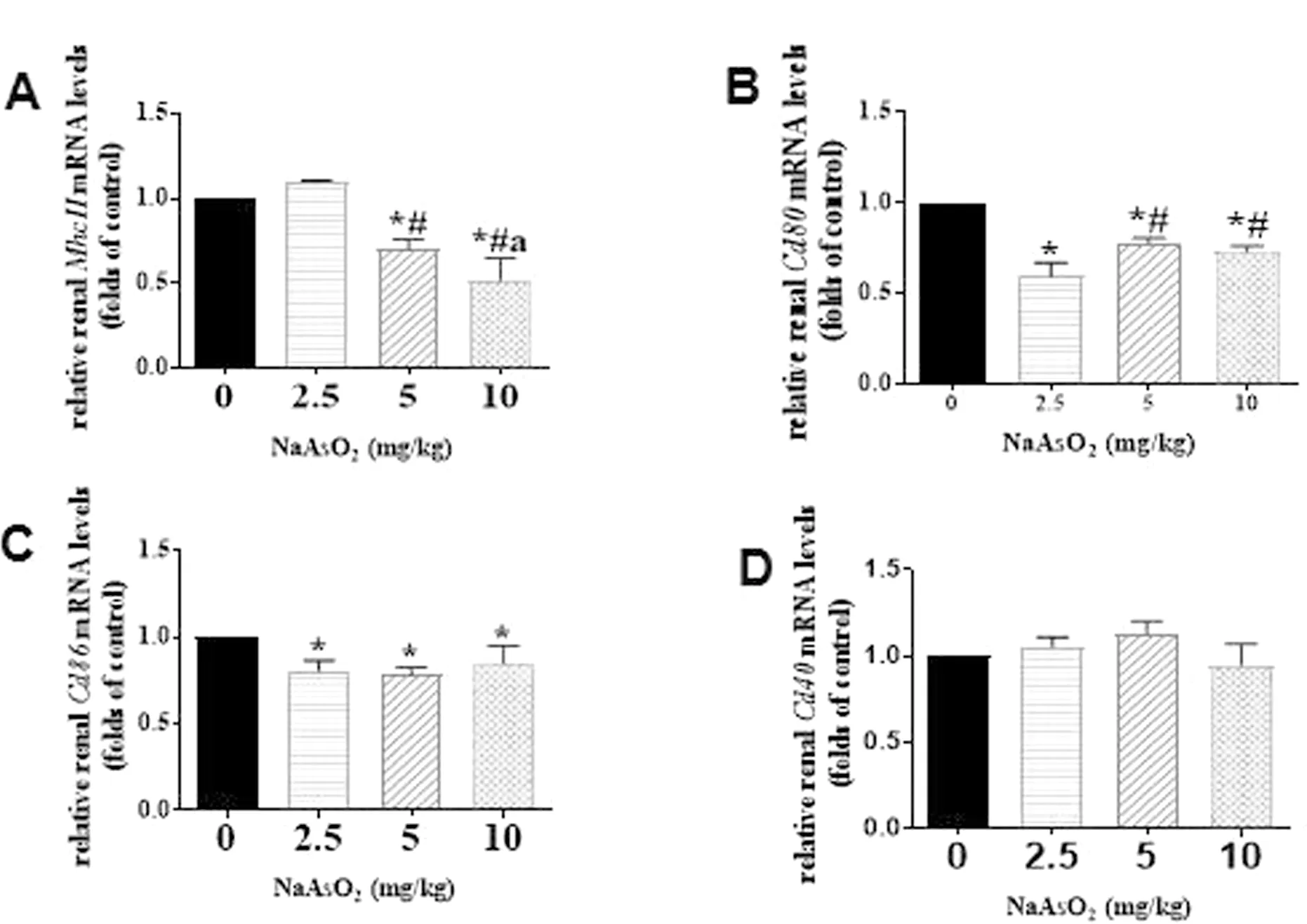
圖5 對照組和砷暴露組小鼠腎組織中MhcⅡ、 Cd80、Cd86 和Cd40 的mRNA 表達水平比較
3 討論
眾所周知, 腎臟是砷毒性及其代謝的主要靶器官之一, 長期砷暴露可以影響腎臟的生理功能,引起腎損傷, 甚至導致腎癌等。 隨著免疫學研究的進展, 研究發現非免疫器官腎臟中也含有樹突狀細胞、 巨噬細胞和淋巴細胞等免疫細胞組成的免疫微環境, 也發揮著一定的免疫調節效應[4-5]。
BUN 是蛋白質代謝的主要終產物, 而Cr 是肌肉在體內代謝的產物, 二者主要通過腎小球濾過排出體外, 故血清BUN 和Cr 是評價腎臟功能的重要指標[7]。 本研究發現與對照組比較, 砷暴露組小鼠血清BUN 的水平明顯增加, 差異有統計學意義, 而血清Cr 的水平無明顯差異。 Mahajan 等[8]研究發現Wistar 大鼠50 ~150 μg/L 砷暴露28 d,其血清BUN 和Cr 明顯增加。 徐昉等[9]的研究結果顯示長期低劑量砷攝入時, 隨著時間的延長, 血清BUN 和Cr 才呈明顯升高, 這提示血清Cr 的水平變化不明顯可能與BUN 和Cr 的代謝部位不同和染毒時間過短有關。 人群流行病學研究發現飲水型砷中毒病區居民血清BUN 平均水平高于非高砷區居民[10], 這些結果均提示砷暴露能夠引起腎功能的損害。
ACP 是溶菌體酶的重要組分, 存在于前列腺、肝和腎等多種組織和體液中, 可以直接參與蛋白質、 糖類和脂類的吞噬作用, 具有防御和免疫調節等生理功能[11]。 本研究結果發現, 與對照組比較, 砷暴露小鼠血清中ACP 的活性明顯下降, 腎臟中ACP 活性升高, 差異有統計學意義。 Rana等[12]研究發現Wistar 大鼠20 mg/L 砷染毒3 個月,血清中ACP 水平明顯高于對照組。 田耕晨[11]等研究指出, 三角帆蚌在受到免疫激活時, 血清中ACP 呈現明顯先升后降趨勢。 本研究結果在一定程度上可以推斷是與血清和腎臟組織對外源性物質刺激時反應快慢不同有關。 同時, 也可能與染毒的時間和動物的種屬不同有關。
Ccr5 和Ccr7 是樹突狀細胞表面的趨化因子受體; Icam 既是細胞間黏附分子又是抗原提呈細胞上的協同刺激分子, 這些分子均調節機體的免疫反應[13]。 本研究發現與對照組比較, 10 mg/kg 砷暴露明顯降低小鼠腎臟組織中Ccr5、Ccr7的mRNA的表達, 砷暴露明顯降低小鼠腎臟組織中Icam1的mRNA 的表達, 差異均有統計學意義。 Zhao 等[14]研究發現C57BL/6 小鼠飲用含砷(0.1、 0.1、 1、10 mg/L NaAsO2) 水12 個月后, 脾和肺組織中Ccr5和Ccr7的mRNA 水平明顯高于對照組。 另一項研究發現100 μg/L 砷暴露明顯增加小鼠腎臟Icam1 的蛋白水平[15]。 這些結果的不一致可能與砷的劑量、 持續時間及作用的靶器官的不同有關,還需要進一步研究和探討。 本研究結果提示砷引起的腎臟組織中趨化黏附因子的表達水平的下降可能參與砷的免疫調節效應。
多種免疫細胞表面分布有Cd40、 Cd80 和Cd86 等多種分化抗原, 具有多種生物學功能。Cd40 與Cd40 的受體Cd40L 結合后可上調其表面的Cd80、 Cd86 及MhcⅡ的表達, 增強抗原提呈的作用[16]。 本研究發現與對照組比較, 砷暴露小鼠腎組織中MhcⅡ、Cd80和Cd86分子的mRNA 水平明顯下降, 差異均有統計學意義。 Bahari 等[17]研究發現樹突狀細胞經1μg/L 砷處理12 h 和24 h后, Cd40 和MhcⅡ分子的表達下降, 影響樹突狀細胞的分化和成熟。 Macoch 等[18]的研究也發現,1~2 μmol/L 砷會抑制樹突狀細胞表面共刺激分子Cd80 和Cd86 的表達。 這些結果均提示砷暴露可以抑制機體的抗原提呈功能。
綜上所述, 本研究發現急性砷暴露破壞小鼠腎臟內抗原吞噬功能分子ACP 的活性, 抑制趨化黏附功能相關分子(Ccr5、 Ccr7 和Icam1) 以及抗原提呈功能相關分子(MhcⅡ、 Cd80 和Cd86) 表達, 從而破壞腎臟正常的免疫功能, 進而導致腎臟損傷, 為砷導致腎臟免疫損傷提供了理論依據。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
現代臨床醫學(2021年1期)2021-01-26 00:56:02
學苑創造·A版(2020年9期)2020-10-13 09:41:02
人大建設(2019年12期)2019-05-21 02:55:32
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
