基于MDCK懸浮細胞灌注培養的流感病毒生產過程開發
2020-11-02 08:51:40吳熠瀟ThomasBissingerYvonneGenzel劉旭平UdoReichl譚文松
生物技術通報 2020年10期
關鍵詞:生長
吳熠瀟 Thomas Bissinger Yvonne Genzel 劉旭平 Udo Reichl 譚文松
(1. 華東理工大學 生物反應器工程國家重點實驗室,上海 200237;2. Max Planck Institute for Dynamics of Complex Technical Systems,
Germany 39106)
相較于傳統的雞胚生產法,用動物細胞來生產流感疫苗正在逐步成為主流生產方式[1]。近年來,季節性流感疫苗的市場需求正在逐步上升,且在眾多接種率較低的國家也有巨大的潛在市場開發前景。自2009年后無大流行流感爆發,卻依然對人類社會存在潛在威脅[2-3]。盡管面對日益增長的市場需求和能快速應對大流行流感病毒爆發的需求,細胞培養法生產流感病毒存在著產能低下的問題,制約了這一技術的革新與發展,因此迫切需要提高細胞培養生產流感疫苗的工作效率,主要涉及細胞株篩選、培養基優化、過程優化及強化等方面[4]。
灌注培養是實現過程強化提高過程效率的重要方法之一,相較于傳統的批培養和補料培養因其小體積大產能的優勢廣泛研究于抗體生產領域[5]。灌注培養過程通過灌注速率控制策略對培養體系進行補液,同時以相同的流速將陳舊的培養液排出,卻將細胞通過截留裝置留在反應器內。近幾年來該技術也逐漸被應用于病毒疫苗生產領域來提高病毒產量、過程效率和經濟性[6]。目前已有報道通過使用基 于AGE1.CR、CAP、PER.C6、HEK293、MDCK、PBG.PK2.1等細胞介質的灌注培養過程來生產流感病毒,病毒對于細胞介質的敏感性和病毒產量均存在差異[7-10]。MDCK細胞目前仍被認為是最適用于流感病毒擴增的細胞介質之一,但至今使用MDCK懸浮細胞的高密度培養來實現流感病毒的高效和快速擴增還鮮有報道[11-12]。
本研究以懸浮MDCK細胞株為試驗對象,通過病毒馴化獲得適于在該MDCK細胞株中擴增的工作種毒,繼而采用稀釋流加的策略評估其在低密度下的生長和產毒表現。然后引進Semi-perfusion模型來模擬MDCK細胞高密度生長和產毒的可行性并研究MOI和胰酶濃度對病毒復制的影響,最后將優化的操作參數應用于反應器體系的ATF灌注培養,實現細胞高密度生長和病毒高效率擴增。
1 材料與方法
1.1 材料
1.1.2 病毒株 原始種毒為模式甲型流感A/Puerto Rico/8/34病毒株(德國Robert Koch Institute),早期在MDCK貼壁細胞(英國ECACC)傳代,隨后在該MDCK懸浮株中適應傳代,其TCID50值為1.8×109virions/mL。
1.2 方法
1.2.1 細胞培養
1.2.1.1 搖瓶培養 種子細胞傳代:MDCK懸浮細胞以0.5×106cells/mL的密度接種于工作體積為30 mL的搖瓶中(Corning),置于37℃,5%的CO2條件下進行懸浮培養,轉速100 r/min,每隔3 d使用Xeno-CDM1傳代一次。搖瓶批式培養:取對數生長期的MDCK種子細胞以1.0×106cells/mL的密度接種于搖瓶中,工作體積為30/50 mL,培養至48 h用于病毒馴化或72 h用于稀釋流加病毒感染或48 h用于semi-perfusion培養。使用培養基為Xeno-CDM1。搖瓶高密度(Semi-perfusion)培養 取對數生長期的MDCK種子細胞以1.0×106cells/mL的密度接種于搖瓶中,工作體積為50 mL,培養至48 h用于后續Semi-perfusion培養。使用培養基為Xeno-CDM1。
1.2.1.2 生物反應器中培養 將搖瓶中擴增完畢的MDCK細胞以1.0×106cells/mL的密度接種到1 L DASGIP 反應器中,工作體積為500 mL,細胞生長至62 h時開始灌注培養。當細胞生長至40×106cells/mL的密度時以150 mL/h的灌流速度部分更換反應器內培養基,共2 h。細胞培養所用培養基為 Xeno-CDM2。細胞生長階段反應器參數設置為:pH為7.15±0.02,DO為40%空氣飽和度,溫度為37℃,攪拌轉速為80 r/min。
1.2.2 病毒感染
1.2.2.1 搖瓶中的病毒感染 病毒馴化:轉移培養至48 h的MDCK細胞培養液至離心管,全離心換液后轉移回125 mL搖瓶進行病毒感染實驗,胰蛋白酶添加量為30 U/mL,以MOI為10-5添加貼壁細胞種毒P0,置于37℃和5% CO2中繼續培養至細胞密度最高時收獲病毒液作為下一次馴化的懸浮細胞種毒P1。重復上述操作分別獲得種毒P2、P3、P4、P5和P6。P6為后續病毒感染實驗所用的工作種毒。稀釋流加感染:培養至72 h的MDCK細胞培養液,在125 mL搖瓶中采用1∶1稀釋流加的方法(15 mL細胞培養液+15 mL新鮮Xeno-CDM1培養基)進行病毒感染實驗。胰蛋白酶添加量為30 U/mL,并以MOI為10-3添加懸浮細胞工作種毒P6,置于37℃和5% CO2環境中繼續培養。高密度Semi-perfusion病毒感染:將培養至高密度的細胞培養液全離心去上清后加入含有30 U/mL或20 U/mL胰酶的培養基,轉移回125 mL的搖瓶后加入MOI為10-1或10-3的工作種毒。5 h后再次全離心換液,之后按照灌注策略進行周期性換液。參數優化實驗組設置如表1。

表1 感染階段不同操作參數實驗組設置
1.2.2.2 生物反應器中的病毒感染 灌注培養中培養基更換結束后,在反應器內加入MOI為10-3的種毒和濃度為20 U/mL的胰酶,灌注培養基更換為加入20 U/mL胰酶的Xeno-CDM2。10-20 min等病毒進入細胞后再重新開始培養基的灌流。病毒復制階段反應器參數設置為:pH為7.20±0.02,DO為40%空氣飽和度,溫度為33℃,攪拌轉速為80 r/min。反應器pH由CO2和空氣的混合氣體和7.5%的NaHCO3溶液同時控制。
根據上文所說,我們可以知道銀行作為經營風險的金融機構,想要在激烈的競爭中脫穎而出就要對風險進行有效地管理控制。而信貸業務是商業銀行中最為基礎的一個業務,只有保證了信貸風險管理的有效性才能夠更好地保證我國商業銀行的正常發展。銀行只有具備了更強更完善的信貸風險管理體系以及處理風險的能力,才能夠樹立更加可靠、穩健的市場形象,提升企業在社會中的影響地位,而且也可以更好地吸引市場中潛在的客戶群體,有利于企業更好更持久的發展,開拓了銀行的業務。
1.2.3 Semi-perfusion培養 以基于細胞特異性灌注速率(Cell-specific perfusion rate,CSPR)的灌注速率控制策略通過在取樣點間周期性的部分離心換液進行Semi-perfusion培養,以此滿足細胞的營養需求。根據反應器中的連續灌注過程,換液體積計算公式:
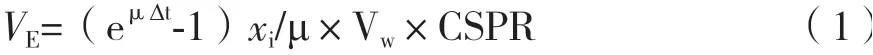
式中,?t為兩取樣點的時間間隔(h);μ為細胞比生長速率(h-1);xi為活細胞密度(cells/mL);Vw為工作體積(mL)。
離心換液自接種后48 h開始。細胞計數后根據式(1)計算換液體積,從搖瓶中取出對應體積的細胞培養液于離心管中,離心換液后重懸細胞轉移回搖瓶,維持工作體積50 mL。隨著細胞密度的上升換液體積增大,直至工作體積的60%。從此時起,固定每次的換液體積,根據公式(1)不斷縮小換液時間間隔。計算時CSPR選擇為60 pL/cell/d[7](圖1)。
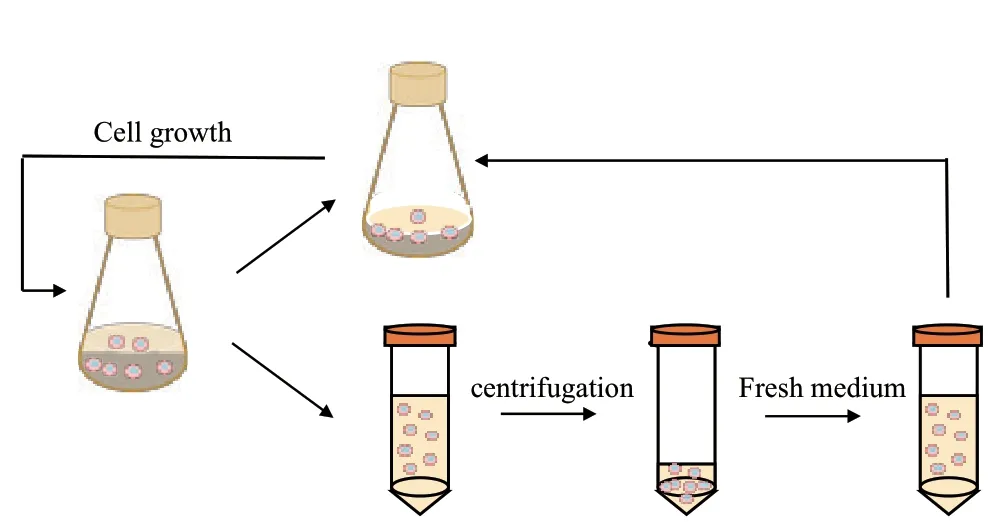
圖1 搖瓶中semi-perfusion的操作流程示意圖
1.2.4 ATF灌注培養 生物反應器與一個交替切向流(Alternating tangential flow,ATF)過濾中空纖維柱灌注系統(ATF2,德國Repligen)相連接。細胞接種62 h后開啟灌注系統和培養基灌流。部分細胞培養液通過灌注系統底部隔膜交替的膨脹和收縮在ATF柱子和連接管中往復運動,無細胞的培養液通過蠕動泵的轉動透過ATF膜流向集液瓶。ATF中空纖維膜中的液體流速控制在0.8 L/min。灌注策略中的CSPR為60 pL/cell/d(圖2)。
1.2.5 細胞計數 通過臺盼藍染色原理用Vi-CELL(美國Beckman Coulter)自動計數儀測量細胞培養過程中的細胞密度和活性。

圖2 基于ATF系統的灌注培養過程反應器裝置設置
1.2.6 病毒滴度檢測 總流感病毒顆粒通過Kalbfuss所報道的血凝素(Hemagglutinin,HA)滴度檢測方法測定[13]。HA滴度以log10表示。單細胞病毒產量(CSVY)計算如公式(2),單位為virions/cell。
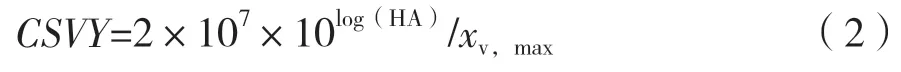
式中,xv,max為病毒復制階段的最大活細胞密度(cells/mL);HA為 血 凝 素 滴 度[log10(HAU/100 μL)]。
由于病毒復制階段的多次換液造成的部分病毒液收獲,需要計算累積HA滴度,才能對應反應器中的連續灌注過程。計算公式如式(3)。
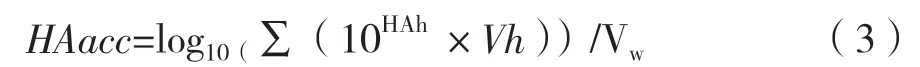
式 中,HAacc為 累 積HA滴 度(log10(HAU/100 μL);HAh為每次病毒收獲液測得的HA滴度(log10(HAU/100 μL);Vh為每次病毒收獲液的體積(mL)。
感染性病毒顆粒根據Genzel和Reichl報道的半數組織培養感染劑量(TCID50)檢測方法測定,單位為virions/mL[14]。
1.2.7 胞外代謝物檢測 葡萄糖、乳酸、谷氨酰胺和氨的濃度由Bioprofile 100 plus(美國Nova medical)通過外部標樣校準檢測得到。
2 結果
2.1 流感病毒在MDCK懸浮細胞中的馴化
將實驗室早期已適應于MDCK貼壁細胞的流感病毒以低MOI 10-5的盲傳方式在該MDCK懸浮細胞中進行馴化。經過5代馴化之后的結果如圖3所示。HA檢測法表征培養液中的總病毒顆粒數。由圖3-A可知,每一代的HA滴度都呈現先上升后平穩的趨勢(除第3代),達到最大HA值的時間第5代相較第1代縮短了24 h(第1代為60 h,第5代為36 h),表明馴化之后,病毒對細胞的感染與自我復制速率明顯加快。5次病毒傳代的最大HA滴度分別為3.52、3.49、3.33、3.60、3.59 log10(HAU/100 μL),在 整個馴化過程中最大HA保持穩定。TCID50檢測法用來定量培養液中的感染性病毒顆粒數。由圖3-B可知,每一代的TCID50滴度在達到最大值后出現了下降的現象,病毒復制過程中的最大TCID50滴度分別為1.78、1.33、1.78、3.16×109virions/mL,表明了馴化過程并沒有導致感染性病毒顆粒數的顯著減少,而是維持了穩定的水平。
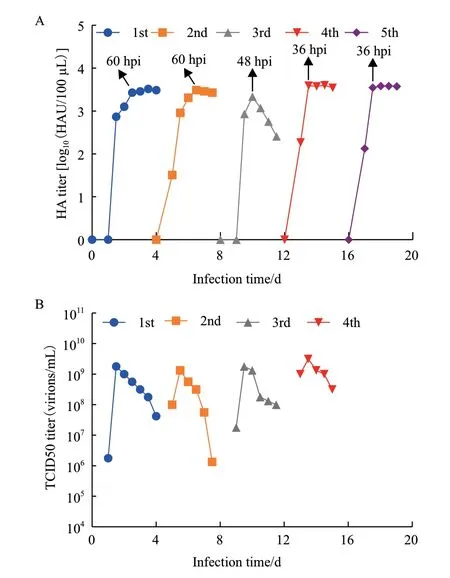
圖3 流感病毒在MDCK懸浮細胞馴化過程中的HA滴度(A)和TCID50滴度(B)變化
2.2 稀釋流加培養條件下的流感病毒生產
經過病毒馴化獲得了能在該MDCK細胞中快速增殖的種子病毒后,在搖瓶中進行了該細胞在稀釋流加策略下的生長和產毒表現,結果如圖4所示。由圖4-A可知,在細胞生長階段,當以1×106cells/mL的密度接種時,72 h后細胞密度達到(10.35±1.07)×106cells/mL,在此期間呈對數生長,平均比生長速率和倍增時間分別為0.033 h-1和21 h;細胞活性維持在(97.7±0.57)%以上。病毒感染前,由于培養基的1∶1補充細胞密度減半;在病毒復制階段細胞繼續生長一段時間后由于病毒的釋放和細胞的裂解細胞密度和活性都迅速下降。由圖4-B可知,病毒在細胞內的復制速度快且效率高,在24 hpi時便幾乎完成了增殖和釋放過程,最大HA滴度為(3.57±0.17)log10(HAU/100 μL)。通過公式(2)計算可得到搖瓶批培養中的病毒感染過程的CSVY為(11 225±1 832)virions/cell。
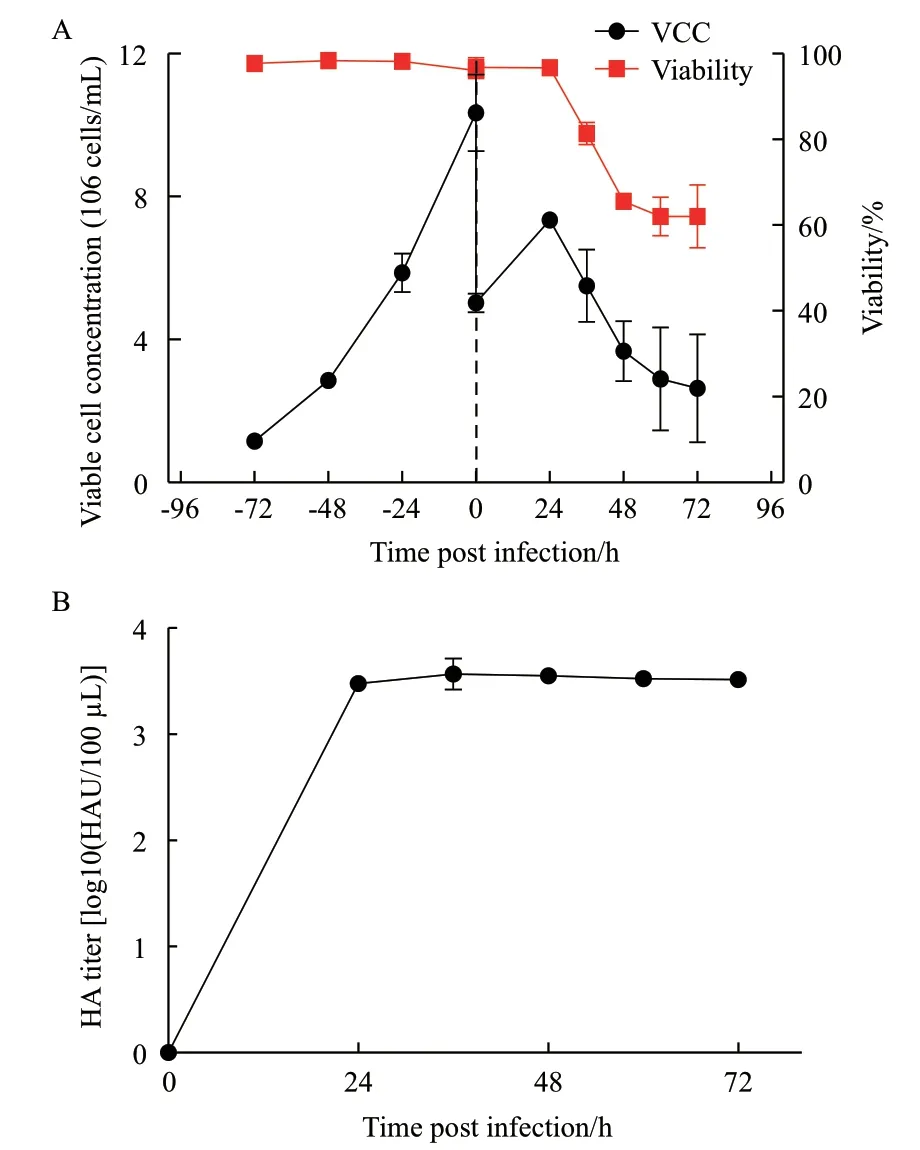
圖4 搖瓶中稀釋流加培養條件下的細胞密度和活性(A)以及HA滴度(B)
2.3 Semi-perfusion中MDCK細胞的高密度生長與流感病毒生產可行性
分析了該MDCK懸浮細胞低密度生長和產毒過程的特征后,展開了以搖瓶為縮小模型去模擬灌注過程中細胞高密度培養條件下(>15×106cells/mL)生產流感病毒的可行性分析,考察病毒感染復制階段不同操作參數胰酶濃度和MOI在灌注模式下對于流感病毒擴增的影響,并同時關注流感病毒在高細胞密度條件下擴增時是否出現CSVY下降的現象,即“高細胞密度效應”,結果如圖5所示。在預實驗中,通過優化灌注策略中的CSPR,確定CSPR為60 pL/cell/d以延長具有高比生長速率的指數生長期,滿足細胞高密度生長的營養需求。如圖5-A所示,在細胞生長階段,以1×106cells/mL的密度接種,約130 h后HCD1、HCD2、HCD3三個實驗組的細胞可分別生長至43.2、44.0、46.8×106cells/mL,且隨著細胞密度的上升,比生長速率下降,最低時僅為0.015/h。在病毒復制階段,僅HCD3實驗組的細胞在病毒感染后繼續生長,且密度維持了一段時間至34 hpi后由于病毒的累積才開始下降,說明在高密度情況下高MOI的病毒感染與復制要快于低MOI。通過比較HCD1和HCD2組,可發現高胰酶濃度組在病毒感染后細胞密度出現明顯下降,而低胰酶濃度組密度基本維持穩定,說明30 U/mL的胰酶濃度對細胞的傷害會比20 U/mL的胰酶濃度明顯。由圖5-B可知,有更高MOI的實驗組HCD1和HCD2病毒釋放較早,在13 hpi時已檢測到高滴度3.83、3.81 log10(HAU/100 μL),而較低MOI組HCD3在24 hpi才開始檢測到滴度3.72 log10(HAU/100 μL)。3個實驗組之后由于細胞的死亡和換液滴度都出現HA滴度下降的趨勢。比較HCD1和HCD2組發現了相似的HA滴度生產動力學,因此所選擇的兩個胰酶濃度對于HA生產的影響作用并不大。通過公式(3)計算得到的不同實驗組累積HA滴度沒有明顯差異,分別為4.05、4.19、4.12 log10(HAU/100 μL),而HCD2實驗組的CSVY最高,為7 016 virions/cell,但是仍低于批培養中的CSVY,即出現了“高密度效應。”綜合考量,選用MOI 0.001和胰酶濃度 20 U/mL作為后續反應器驗證的感染階段參數。
2.4 基于ATF的生物反應器中的灌注培養過程驗證
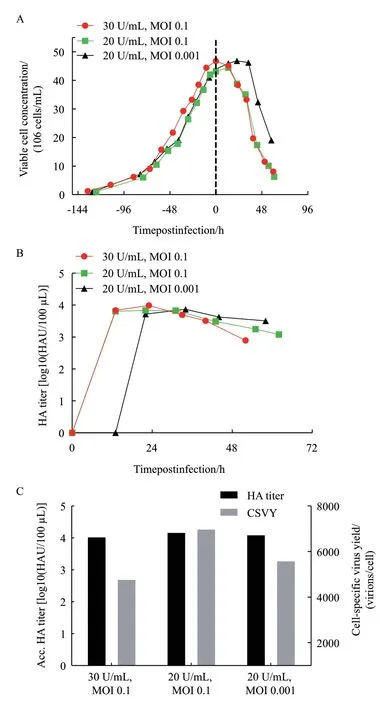
圖5 Semi-perfusion中不同胰酶濃度和MOI對MDCK細胞生長(A)、流感病毒HA滴度(B)和累積HA滴度以及單細胞病毒產量(C)的影響
完成了搖瓶中的細胞高密度生長與流感病毒生產的可行性分析后,進行了ATF生物反應器灌注培養的可行性驗證。在Semi-perfusion的結果基礎上,調整了培養基的緩沖溶液體系提升了反應器的pH控制,同時也降低了病毒復制階段的溫度。由圖6-A和6-B可知,在細胞生長階段,與Semi-perfusion相似,細胞在反應器中經過136.5 h達到41.2×106cells/mL的密度,也同樣出現了比生長速率下降的現象。另外也發現隨著培養液滲透壓的下降,細胞的直徑也相應變小。接毒后,由于降溫病毒累積自24 hpi開始,至36 hpi時達到4.23 log10(HAU/100 μL)。此外,在ATF柱子的透過液中也檢測到了HA滴度,表明有部分流感病毒通過了ATF膜,未截留在反應器內。通過計算得到累積HA滴度為4.37 log10(HAU/100 μL),相 應 的CSVY為10 083 virions/cell,說 明 反應器灌注培養過程通過降溫策略可以緩解甚至克服“高密度效應”。從圖6-C可知,灌注速率隨著細胞密度的變化而變化,在細胞密度最高時達到了3.12 RV/d,由于每個時間段內灌注速率的值根據該時間段結束時的預期細胞密度計算得到,所以整個培養過程灌注速率呈現一個階梯式的變化。根據每個時間段所消耗的培養基的體積計算過程中實際的CSPR發現實際CSPR在62-84 pL/cell/d之間(除病毒感染前的2 h換液階段),說明基于CSPR的灌注速率控制策略效果尚佳。從圖6-D可知,葡萄糖和谷氨酰胺在灌注啟動前被消耗,在灌注啟動后由于新鮮培養基的補給兩者濃度處于穩定的狀態,分別約為18 mmol/L和3.5 mmoL/L;相應地,代謝副產物乳酸和氨在灌注啟動前被生成,之后由于灌注過程中培養基的補入也處于穩定的狀態,約為25 mmol/L和3.5 mmol/L。病毒感染初期階段(<24 hpi),盡管由于培養基的補入,葡萄糖仍被大量地消耗,造成了乳酸的大量生成,高達35.1 mmol/L。
3 討論
目前用動物細胞來生產流感疫苗存在的產能低下問題制約了該工藝的發展,因此提高過程的生產效率成為了解決這個問題的關鍵[15-16]。通過灌注培養模式來實現過程強化是提高病毒生產效率的有效途徑之一[17]。本研究以一株MDCK懸浮細胞株為研究對象,通過病毒馴化,批培養驗證以及灌注培養過程開發,成功地建立了一個高效的流感病毒生產平臺,成果討論歸納于表2。
3.1 病毒馴化是疫苗生產過程的必要步驟
流感病毒在細胞內的馴化通常采用低MOI,因為高MOI的情況下會產生更多的無感染性且不完整的缺陷型病毒顆粒(DP),其中的缺陷型干擾顆粒(DIP)會進一步干擾完整病毒顆粒的復制,導致感染性病毒顆粒的減少[18]。而低MOI情況下生產的種毒感染性顆粒比例更高,因此在作為工作種毒時使用體積更小,且在細胞內復制時受到DIP的干擾也更小。通過病毒馴化加快病毒復制過程的現象也見于Vero和HEK293細胞中[19-20]。根據病毒種類的不同,病毒復制在馴化之后的改善程度也不同,本文中的H1N1株在馴化之后維持了馴化前的病毒滴度,而之前報道的部分黃病毒如MVA和Zika病毒在馴化之后感染性滴度得到了1個log的提升[21]。
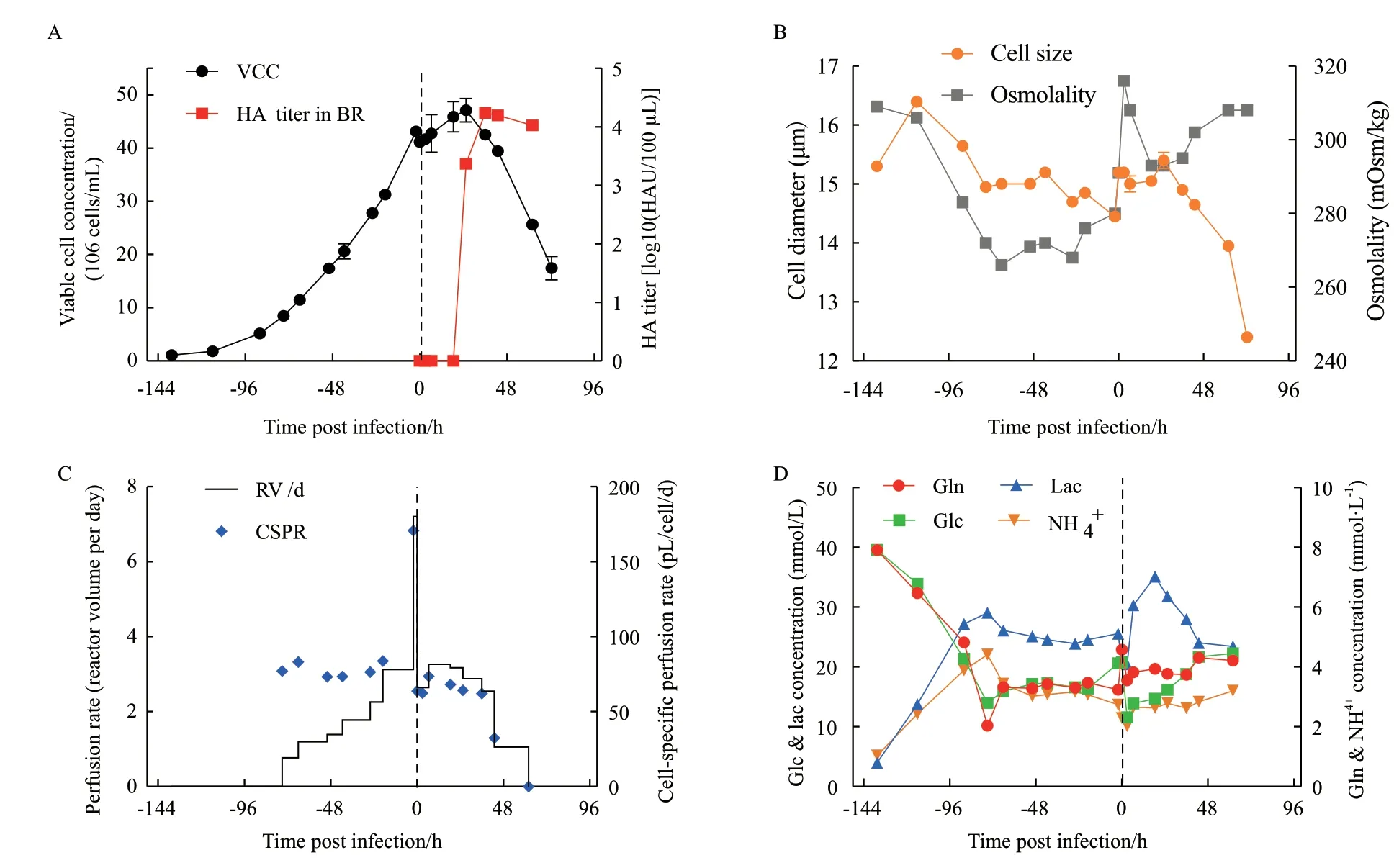
圖6 ATF灌注培養過程中的細胞密度和病毒HA滴度(A)、細胞直徑和培養液滲透壓(B)、灌注速率和CSPR(C)以及胞外谷氨酰胺、葡萄糖、乳酸和氨的濃度(D)

表2 MDCK細胞生產流感病毒技術成果表
3.2 MDCK細胞作為流感病毒生產基質的成功運用
提高流感病毒產量的關鍵在于提高細胞密度和單細胞產量。細胞株篩選、培養基開發和反應器過程控制均可影響細胞密度和單細胞產量。本研究中的MDCK懸浮細胞批培養中生長密度可至千萬級別,高于目前已報道的大部分MDCK細胞[20,22-23],且它在該培養基中的CSVY高于11 000 virions/cell,這兩者的結合保證了最終的流感病毒高產量。本研究中獲得的病毒HA高于目前已有文獻報道使用不同的細胞介質如PER.C6、HEK293、PBG.PK2.1生產流感的其他批培養或稀釋流加培養過程,再一次驗證了MDCK細胞為最佳的候選株之一用于流感病毒的生產[11]。
3.3 過程強化實現了病毒產量的進一步提升
灌注培養在病毒疫苗生產領域近幾年發展迅速,基于該株MDCK細胞在低密度培養條件下的優良產毒表現,通過灌注培養實現過程強化,提高病毒產量和產毒效率。以搖瓶或搖管作為Scale-down模型來模擬實際反應器中的灌注過程并進行高密度培養的過程的開發與優化既可實驗高通量篩選也降低了操作復雜程度和成本[24]。經過前期的灌注速率優化,CSPR為60 pL/cell/d可支持細胞達到40×106cells/mL的密度。通過此模型可獲得更高的細胞密度,但是也會由于頻繁的離心與換液造成過程的不穩定,影響細胞的生長。由于選擇的灌注策略的影響,細胞密度達到一定水平時換液時間間隔會較小(< 4 h),由此會引起過程中pH、溫度、滲透壓的不穩定,從而影響細胞生理狀態。
MOI對于病毒復制過程有重要影響,在此體系中選用高MOI可緩解由于不斷的換液操作帶走體系中的病毒顆粒而延緩復制過程的問題,選用低MOI則可以減少大規模生產時的種毒接種量,也可以減少DIP的產生。本研究通過比較MOI 0.1和0.001發現在高密度條件下MOI對于細胞生長和病毒復制的影響與批培養條件有相似性[25]。30 U/mL的胰酶濃度對細胞的傷害明顯高于20 U/mL的胰酶濃度,但是并沒有發現對病毒產量有明顯的抑制作用。通過Semi-perfusion得到的>4 log10(HAU/100 μL)的滴度與已有報道中的另一株MDCK懸浮細胞株病毒產量相似,但是報道缺乏反應器的灌注過程驗證[7]。
本研究最后進行了灌注培養的反應器驗證及過程表征,反應器中相似的細胞生長曲線驗證了Semiperfusion模型模擬反應器培養的可行性,且病毒復制階段的降溫措施提高了CSVY,克服了“細胞密度效應”。根據已有報道,盡管流感病毒的直徑(80 nm)小于ATF膜的孔徑(0.2 μm),卻由于膜對培養液中的細胞碎片、某些蛋白、DNA和化合物的非特異性吸附幾乎沒有病毒顆粒可以穿過ATF膜進入到透過液中[9]。本研究中卻發現了在透過液中約有25%的病毒顆粒,原因可能是由于降溫措施減少了培養液中的宿主細胞蛋白和DNA水平,仍需進一步檢測培養液蛋白和DNA濃度進行驗證。最終獲得的累積HA滴度為現有應用反應器灌注培養過程生產流感病毒的報道中最高,而采用MDCK細胞更是首次[8]。目前過程中的灌注速率控制是為半連續,需要人工階段性地根據細胞密度調整灌注速率,后期的研究可以根據PAT的理論引入在線活細胞密度檢測體系去實現灌注速率自動化控制。此外,代謝物檢測發現病毒感染階段乳酸產量較高(約35 mmol/L),其對病毒產量的潛在影響尚不能確定。培養基的“深度”(Depth in medium)優化可改善病毒復制階段的副產物積累問題,同時也有希望降低整個過程中的CSPR,從而降低培養基消耗量和提高過程經濟性[5]。
4 結論
低MOI的病毒馴化可明顯加快病毒在細胞內的感染與復制進程,且不影響總病毒滴度和感染性病毒滴度。搖瓶中的稀釋流加培養可使細胞密度達千萬級別,病毒滴度達(3.57±0.17)log10(HAU/100 μL)。通過Semi-perfusion模型驗證了MDCK細胞高密度生長和產毒的可行性,且發現不同MOI和胰酶濃度對病毒感染復制有影響作用。最后通過反應器中的灌注培養使細胞密度高達40×106cells/mL,病 毒產 量 高 達4.37 log10(HAU/100 μL),CSVY為10 083 virions/cell,與批培養過程相當。本工作嘗試的結果提升了流感病毒的生產效率,為基于細胞培養的流感疫苗產業化生產提供了新選擇。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
