蛋氨酸調(diào)控畜禽氧化應(yīng)激的研究進(jìn)展
2020-11-02 08:51:30劉融崔凱白福恒刁其玉
生物技術(shù)通報 2020年10期
關(guān)鍵詞:氧化應(yīng)激研究
劉融 崔凱 白福恒 刁其玉
(1. 蘭州大學(xué)草地農(nóng)業(yè)生態(tài)系統(tǒng)國家重點(diǎn)實(shí)驗(yàn)室 蘭州大學(xué)草地農(nóng)業(yè)科技學(xué)院,蘭州 730020;2. 中國農(nóng)業(yè)科學(xué)院飼料研究所 農(nóng)業(yè)農(nóng)村部
飼料生物技術(shù)重點(diǎn)實(shí)驗(yàn)室,北京 100081;3. 德州市陵城區(qū)白富恒養(yǎng)殖場,德州 250523)
氧化應(yīng)激(Oxidative stress)是指機(jī)體受到外界刺激產(chǎn)生并積累過多自由基所導(dǎo)致的體內(nèi)氧化還原失衡狀態(tài)。在生產(chǎn)中,氧化應(yīng)激引發(fā)腹瀉等動物性疾病、導(dǎo)致幼畜成活率下降,并對畜禽生產(chǎn)水平和畜產(chǎn)品品質(zhì)造成嚴(yán)重影響[1]。畜禽產(chǎn)生氧化應(yīng)激的主要原因包括機(jī)體的生理狀況、日糧營養(yǎng)水平和環(huán)境因素等。
營養(yǎng)物質(zhì)是動物體內(nèi)自由基產(chǎn)生的物質(zhì)基礎(chǔ),也是動物體內(nèi)清除自由基的物質(zhì)來源。近年來,關(guān)于營養(yǎng)水平與機(jī)體健康的研究成為生命科學(xué)領(lǐng)域的焦點(diǎn)。2019年,Cell雜志同時發(fā)表了3篇文章詳細(xì)報道了禁食和嚴(yán)重卡路里限制對不同免疫細(xì)胞水平和定位及其機(jī)制的影響[2-4]。Nature報道密西根大學(xué)Ursula Jakob教授團(tuán)隊關(guān)于生命早期的氧化應(yīng)激會讓線蟲活得更久的最新發(fā)現(xiàn)[5]。模式動物日糧中能量和蛋白質(zhì)攝入限制引起的細(xì)胞老化速率降低等現(xiàn)象可能是由蛋氨酸限制引起的ROS產(chǎn)生及氧化損傷的減少導(dǎo)致的[6]。
限制飲食中蛋氨酸可以迅速而有效地改變機(jī)體的代謝狀態(tài),尤其是以蛋氨酸為中心的代謝路徑。多項(xiàng)研究證實(shí),蛋氨酸代謝調(diào)控炎癥性巨噬細(xì)胞活化的免疫代謝,蛋氨酸限制飲食可影響腫瘤的發(fā)生和治療。蛋氨酸對于緩解機(jī)體氧化應(yīng)激,提高機(jī)體抗氧化能力具有重要影響[7-8]。MetR可以通過減少線粒體活性氧產(chǎn)生、調(diào)控Nrf2、ATF4、AP-1等轉(zhuǎn)錄因子介導(dǎo)的關(guān)鍵信號通路以及改變miRNAs表達(dá)等方式參與氧化應(yīng)激的調(diào)控,改善氧化狀態(tài)并在一定程度上促進(jìn)機(jī)體健康。本文綜述了氧化應(yīng)激的產(chǎn)生、蛋氨酸調(diào)控氧化應(yīng)激的機(jī)制及信號通路,以期為畜牧生產(chǎn)提供理論指導(dǎo)。
1 氧化應(yīng)激的產(chǎn)生及影響
1.1 氧化應(yīng)激的產(chǎn)生機(jī)理
體內(nèi)自由基產(chǎn)生過多,超出機(jī)體抗氧化系統(tǒng)的清除能力,機(jī)體氧化系統(tǒng)和抗氧化系統(tǒng)失衡,造成細(xì)胞與機(jī)體氧化損傷,即產(chǎn)生了氧化應(yīng)激。機(jī)體自由基主要包括活性氧族(Reactiveoxygenspecies,ROS)和活性氮族(Reactivenitrogenspecies,RNS)。機(jī)體內(nèi)抗氧化系統(tǒng)包括酶類和非酶類,酶類抗氧化系統(tǒng)主要是超氧化物歧化酶(Superoxidedismutase,SOD)、輔 酶Q、谷 胱 甘 肽 過 氧 化 物 酶(Glutathioneperoxidase,GSH-Px)、谷胱甘肽還原酶(GSSG-R)、谷胱甘肽轉(zhuǎn)移酶(Glutathionetransferase,GST)、過氧化氫酶(Catalase,CAT)等;非酶類抗氧化系統(tǒng)主要有谷胱甘肽、維生素A、維生素C、維生素E、β-胡蘿卜素、茶多酚等;另外,體內(nèi)高濃度的游離氨基酸也可作為活性氧的清除劑[9]。以上物質(zhì)能夠直接清除自由基或通過抵抗自由基發(fā)揮作用,進(jìn)而保護(hù)機(jī)體免受自由基的傷害。氧化還原狀態(tài)的判定是必要的,目前通過檢測機(jī)體本身的氧化還原狀態(tài)以及組織或血液中的氧化應(yīng)激代謝產(chǎn)物、抗氧化物質(zhì)(抗氧化酶和非酶抗氧化劑)可以對機(jī)體氧化應(yīng)激狀態(tài)進(jìn)行綜合判定[10]。
1.2 氧化應(yīng)激對機(jī)體以及畜禽生產(chǎn)的影響
氧化應(yīng)激會引發(fā)細(xì)胞內(nèi)蛋白質(zhì)、脂類和DNA等細(xì)胞大分子的氧化損傷,造成遺傳信息的丟失或改變,也可使細(xì)胞膜上不飽和脂肪酸過氧化,改變細(xì)胞膜的通透性,破壞細(xì)胞穩(wěn)態(tài)[9]。在人類病理學(xué)研究中,氧化還原失衡被認(rèn)為與胰島素抵抗、動脈粥樣硬化、癌癥和神經(jīng)退行性疾病的病因和進(jìn)展有關(guān)[11]。在畜禽生產(chǎn)中,氧化應(yīng)激導(dǎo)致幼齡畜禽腹瀉率上升、抗病力下降,成活率降低;降低成年畜禽的生產(chǎn)性能并對畜禽產(chǎn)品品質(zhì)造成不利影響[1]。斷奶應(yīng)激過程存在嚴(yán)重的氧化應(yīng)激并伴隨腸屏障功能受損、疾病易感性增加和內(nèi)分泌失調(diào)等問題,Zhu等[12]的研究表明仔豬斷奶應(yīng)激刺激自由基的產(chǎn)生并抑制抗氧化能力。畜禽飼養(yǎng)中的營養(yǎng)及理化環(huán)境等因素均是氧化應(yīng)激的誘因,溫雅俐[13]的研究表明,熱應(yīng)激可降低奶牛采食量和營養(yǎng)物質(zhì)的消化率,降低產(chǎn)奶量。馬燕芬等[14]的研究表明,熱應(yīng)激破壞奶山羊瘤胃上皮組織結(jié)構(gòu),導(dǎo)致瘤胃黏膜屏障通透性增加,降低其生理機(jī)能和生產(chǎn)性能。
2 蛋氨酸對氧化應(yīng)激的影響
蛋氨酸是一種含硫必需氨基酸,是琥珀酰輔酶A、同型半胱氨酸、半胱氨酸、肌酸和肉堿的前體。蛋氨酸具有重要的生理功能,研究表明蛋氨酸除了作為蛋白質(zhì)合成過程中的起始氨基酸參與機(jī)體蛋白質(zhì)的合成外,還可以影響哺乳動物的先天免疫系統(tǒng)和消化功能,并干預(yù)脂質(zhì)代謝、內(nèi)源性抗氧化酶(如甲硫氨酸亞砜還原酶A)的激活以及谷胱甘肽的生物合成以對抗氧化應(yīng)激[15]。
適量添加蛋氨酸可改善畜禽氧化應(yīng)激狀態(tài),仔豬飼喂高水平含硫氨基酸(SAA)(0.85%),總抗氧化能力,超氧化物歧化酶、過氧化氫酶活性增加[16]。李亮[7]的研究表明,蛋氨酸能夠顯著提高雉雞血清及肝臟抗氧化能力。蛋氨酸過量添加導(dǎo)致蛋氨酸中毒,危害動物健康導(dǎo)致生產(chǎn)性能的降低[17]。模式動物研究結(jié)果多反映出MetR對氧化應(yīng)激的正面效應(yīng),MetR后的高脂日糧小鼠肝臟與血漿中T-AOC和GSH-Px活力顯著增加,丙二醛和ROS含量極顯著降低,血脂和肝臟脂肪積累也顯著降低[8]。雖然MetR可產(chǎn)生如減少內(nèi)臟脂肪、增加胰島素敏感性和改善脂質(zhì)代謝等多種有益的生理效應(yīng)[18],但MetR日糧會降低體重增長速率,所以很明顯,MetR對大型家畜的生產(chǎn)意義不大。王杰[19]的研究表明,相對于蛋氨酸正常添加組,蛋氨酸限制組末重顯著降低,料重比顯著增加。MetR 顯著降低羔羊血清SOD活性,下調(diào)羔羊肝臟組織α型谷胱甘肽轉(zhuǎn)移酶(GSTA)、μ谷胱甘肽轉(zhuǎn)移酶(GSTM)、谷胱甘肽過氧化物酶(GSH-Px)等抗氧化相關(guān)基因的表達(dá)水平。Castellano等[20]的研究表明,日糧蛋氨酸缺乏時生長豬肝臟GSH含量的降低和脂肪組織抗氧化酶活性的增加共同改變了細(xì)胞氧化還原狀態(tài),該實(shí)驗(yàn)中,不同組織的抗氧化酶活性變化趨勢并不一致。目前,蛋氨酸對機(jī)體氧化應(yīng)激的影響的研究并未取得一致結(jié)果,處理效果與添加劑量以及動物種類密切相關(guān),仍需進(jìn)一步研究。
3 MetR調(diào)控氧化應(yīng)激的分子機(jī)制
MetR對細(xì)胞氧化應(yīng)激的調(diào)控是多種機(jī)制共同作用的結(jié)果,其中主要機(jī)制有兩種:MetR可減少線粒體中ROS(mtROS)的產(chǎn)生從而降低細(xì)胞氧化應(yīng)激狀態(tài);MetR作用于信號通路調(diào)控肝臟組織抗氧化基因的表達(dá),從而調(diào)控細(xì)胞氧化還原狀態(tài)。
3.1 MetR對線粒體ROS產(chǎn)生的影響
線粒體電子傳遞鏈由復(fù)合體 I、II、III、IV四個主要的氧化還原復(fù)合體酶系組成,其功能是進(jìn)行生物氧化,并與磷酸化相偶聯(lián),共同產(chǎn)生ATP。電子在傳遞給末端氧化酶之前漏出呼吸鏈與氧反應(yīng)生成超氧自由基的過程是線粒體生成活性氧的源頭。Chance[21]的報道指出,正常生理狀態(tài)線粒體活性氧的產(chǎn)生量相當(dāng)于其耗氧的1%-2%,活性氧會導(dǎo)致脂質(zhì)、蛋白質(zhì)及線粒體DNA(mtDNA)等細(xì)胞大分子損傷。研究表明,活性氧在線粒體復(fù)合物 Ⅰ 和復(fù)合物 Ⅲ 中產(chǎn)生[22],位于親水性復(fù)合物 Ⅰ 結(jié)構(gòu)域中的硫鐵簇等結(jié)構(gòu),或位于復(fù)合物I的內(nèi)膜臂中的泛醌是導(dǎo)致復(fù)合物I ROS生成的主要電子輸運(yùn)組分;復(fù)合物 Ⅲ 的Q0位點(diǎn)的半醌自由基(UQH·)可能是O2生成O2
-·的單電子供體[23]。
大量研究表明,MetR性日糧可以降低線粒體活性氧的產(chǎn)生率,從而降低線粒體氧化損傷[6,24-27]。飼喂80% MetR日糧的大鼠肝臟、心臟線粒體的活性氧生成減少,特別是復(fù)合物I產(chǎn)生的活性氧,mtDNA氧化損傷標(biāo)記物(8-oxodg)濃度降低,線粒體糖氧化損傷標(biāo)記物(CEL和CML)、脂質(zhì)氧化損傷標(biāo)記物(MDAL和CML)、蛋白質(zhì)氧化損傷標(biāo)記物(GSA和AASA)含量降低[6,24]。飼喂40% MetR日糧7周后大鼠的心臟、肝臟、腦和腎臟組織以及30%MetR下仔豬的肝臟線粒體也獲得了類似的結(jié)果[24-26](表1)。在上述研究中,活性氧生成的減少并不是氧通量降低引起的,因?yàn)榛钚匝跎傻臏p少時氧氣消耗量往往是不變或者增加的[6,24-26]。線粒體復(fù)合物濃度和活力的下降以及線粒體自由基泄露百分比的減少是MetR降低線粒體ROS的產(chǎn)生從而降低氧化損傷的主要原因。另外,部分研究顯示線粒體凋亡誘導(dǎo)因子(Apoptosis-inducing factor,AIF)的蛋白質(zhì)豐度降低[26],但它的變化存在組織特異性[25]。
另外,MetR下模式動物壽命的延長支持線粒體自由基老化理論,許多研究結(jié)果認(rèn)為,長壽物種中存在較低的mtROS產(chǎn)生,對內(nèi)源性抗氧化劑或蛋白質(zhì)和DNA修復(fù)系統(tǒng)的需求也較小,這樣有效降低了能量消耗[22]。研究表明MetR日糧下,線粒體中MnSOD含量不變[22-23]。對此研究者認(rèn)為,線粒體氧化應(yīng)激的減少是mtROS產(chǎn)生減少而不是抗氧化酶的增加。MetR作用于線粒體的機(jī)制有待于進(jìn)一步的研究。
3.2 MetR調(diào)控氧化應(yīng)激相關(guān)的信號通路
MetR導(dǎo)致蛋氨酸代謝途徑發(fā)生改變,其中轉(zhuǎn)硫途徑的改變導(dǎo)致肝臟GSH含量下降[28],GSH濃度的降低可能作為信號分子調(diào)控基因表達(dá)。在氧化應(yīng)激通路中Nrf2、ATF4和AP-1/c-Jun是重要的氧化還原反應(yīng)轉(zhuǎn)錄因子[29]。
3.2.1 核因子NF-E2相關(guān)因子(Nrf2) Nrf2-ARE通路是最廣泛的抗氧化應(yīng)激通路之一,是體內(nèi)最大的抗氧化應(yīng)激體系。Nrf2是細(xì)胞抗氧化反應(yīng)重要的調(diào)控因子,細(xì)胞在氧化應(yīng)激和親電子物的過量攻擊時能夠引起Nrf2/Keap1復(fù)合物解聚,游離的Nrf2迅速進(jìn)入細(xì)胞核,與小Maf等堿性亮氨酸拉鏈(bZIP)蛋白形成異二聚體與靶基因中抗氧化應(yīng)答原件(Antioxidant response element,ARE)序列結(jié)合,ARE位于半胱氨酸連接酶催化亞基(Glutamate cysteineligasecatalyticsubunit,GCLC)、血紅素氧合酶1(HO-1)和NAD(P)H醌氧化還原酶1(NQO1)以及α和π型谷胱甘肽轉(zhuǎn)移酶(α-GST;GST-π)等許多解毒和抗氧化酶基因的啟動子區(qū)域[11]。Nrf2與ARE結(jié)合可增強(qiáng)抗氧化酶系統(tǒng)相關(guān)基因的表達(dá),發(fā)揮抗氧化作用,重建細(xì)胞氧化還原穩(wěn)態(tài)。研究表明,MAPKs、PI3K/Akt和蛋白激酶C均可在氧化應(yīng)激下被激活[30]。通過檢測MAPK家族中胞外信號調(diào)節(jié)激酶(ERK)、絲裂原激活蛋白激酶(p38)和c-Jun氨基端激酶(JNK)等轉(zhuǎn)錄調(diào)節(jié)因子的磷酸化水平以及RNA干擾等手段,證實(shí)在小鼠肝臟原代細(xì)胞中,MetR通過激活ERK參與GST的表達(dá)調(diào)控[31]。

表1 MetR對動物線粒體氧化應(yīng)激相關(guān)參數(shù)變化
二相解毒酶GST(包括A3/5、M1和P1)在肝臟組織中的誘導(dǎo)作用在喂食蛋白質(zhì)缺乏日糧或蛋氨酸缺乏日糧的大鼠中得到報道[11]。Tsai等[32]的研究表明,GSTP基因在大鼠原代肝細(xì)胞中的表達(dá)由含硫氨基酸限制特異性誘導(dǎo)上調(diào),而不是通過亮氨酸、異亮氨酸、苯丙氨酸或賴氨酸的限制。Lin等[11]的研究發(fā)現(xiàn),MetR通過ERK-Nrf2-GPEI信號通路上調(diào)GSTP基因轉(zhuǎn)錄。小鼠對MetR日糧肝轉(zhuǎn)錄組學(xué)反應(yīng)的分析表明,蛋氨酸限制性日糧對肝臟轉(zhuǎn)錄組的影響主要集中在對肝臟促炎信號轉(zhuǎn)錄的下調(diào),以及抗氧化反應(yīng)上調(diào)。肝臟中促炎通路基因表達(dá)的減少可能是NFE2l2基因的激活驅(qū)動抗氧化反應(yīng)的結(jié)果[33]。
3.2.2 活 化 轉(zhuǎn) 錄 因 子4(ATF-4) ATF-4是 一 種bZIP轉(zhuǎn)錄因子,由缺氧、內(nèi)質(zhì)網(wǎng)應(yīng)激、氨基酸剝奪和氧化應(yīng)激等應(yīng)激信號誘導(dǎo)的,ATF4 與 CCAAT增強(qiáng)子結(jié)合蛋白(CCAAT /enhancer-binding protein,C/EBP)形成二聚體,與 C/EBP 激活轉(zhuǎn)錄因子應(yīng)答元件(C/EBPactivating transcription factor response elements,CARE)結(jié)合,參與調(diào)控氧化應(yīng)激、氨基酸合代謝、脂質(zhì)代謝、自噬和血管生成等相關(guān)基因的表達(dá)[34]。
Kilberg等[35]綜述了調(diào)控ATF4表達(dá)的幾種途徑,其中MetR可通過PERK/eIF2α/ATF4和GCN2/eIF2α/ATF4通路誘導(dǎo) ATF4的表達(dá)。一般性調(diào)控阻遏蛋白 激 酶2(General control nonderepressible 2kinase,GCN2)是氨基酸缺乏的感受器,在維持機(jī)體氨基酸平衡相關(guān)的信號通路的調(diào)節(jié)過程中發(fā)揮著重要作用,在限制必需氨基酸的細(xì)胞外供應(yīng)或阻斷其它非必需氨基酸的合成所誘導(dǎo)的氨基酸反應(yīng)(AAR)中被激活。幾乎每種氨基酸(包括蛋氨酸)氨基酸限制時,細(xì)胞內(nèi)空載tRNA增多,激活GCN2,從而引起 eIF2α 的磷酸化,上調(diào)ATF-4的表達(dá)[36]。除GCN2外PERK也參與eIF2α 的磷酸化和翻譯控制過程[37]。由理化刺激引發(fā)的內(nèi)質(zhì)網(wǎng)應(yīng)激(Endoplasmic reticulum stress,ERS)會觸發(fā)未折疊蛋白反應(yīng)(Unfolded protein response,UPR)的多條通路,其中包括激活eIF2α的激酶PERK,eIF2α的磷酸化可上調(diào)ATF4的合成和下游轉(zhuǎn)錄程序。GCN2或PERK的營養(yǎng)傳感參與蛋氨酸限制期間對蛋白質(zhì)平衡長期控制機(jī)制,但也有研究表明它們對MetR期間的代謝適應(yīng)是不必要的[18]。
eIF2α的激酶PERK通常在內(nèi)質(zhì)網(wǎng)應(yīng)激條件被激活,但也有研究指出,在沒有內(nèi)質(zhì)網(wǎng)應(yīng)激的情況下,MetR也 可 激 活PERK并 通 過PERK向eIF2α/ATF4和Nrf2傳遞信號。MetR的結(jié)果是通過eIF2α/ATF4激活綜合應(yīng)激反應(yīng)(ISR)并激活Nrf2敏感抗氧化反應(yīng)程序[37]。上述機(jī)制對MetR誘導(dǎo)的整體應(yīng)激反應(yīng)(ISR)具有重要意義,它有助于調(diào)節(jié)細(xì)胞內(nèi)穩(wěn)態(tài)和生理反應(yīng)實(shí)現(xiàn)MetR期間的代謝適應(yīng)(圖1)。
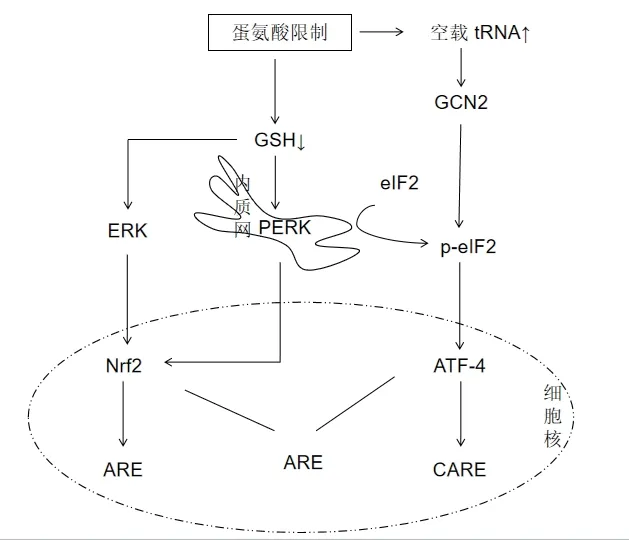
圖1 MetR調(diào)控Nrf2、ATF-4信號通路
3.2.3 激活蛋白1(AP-1)及其他轉(zhuǎn)錄因子 AP-1/c-Jun作為許多細(xì)胞內(nèi)信號級聯(lián)的靶點(diǎn),這些級聯(lián)被細(xì)胞外信號分子(如細(xì)胞因子、生長因子和激素)或各種物理化學(xué)應(yīng)激源激活。AP-1與細(xì)胞增殖、程序性死亡的調(diào)節(jié)和應(yīng)激反應(yīng)有關(guān)[38]。研究發(fā)現(xiàn),L-半胱氨酸和/或L-蛋氨酸供應(yīng)不足降低細(xì)胞GSH含量,并激活氧化還原敏感轉(zhuǎn)錄因子,包括Nrf2、AP-1和NF-kb[39]。
Tsai等[32]研究表明,限制L-蛋氨酸可能通過GSH耗竭從而激活ERK-AP-1信號通路,AP-1與GSTP增強(qiáng)子I(GPE I)的5'上游區(qū)域的TPA反應(yīng)元件(TPAResponse Element,TRE)結(jié)合,上調(diào)GSTP基因的表達(dá)。
AP-1/c-Jun和ATF4除了單獨(dú)參與細(xì)胞應(yīng)激下的反應(yīng),還可與Nrf2共同作用于氧化還原反應(yīng)。Nrf2與其他bZIP成員形成二聚體,參與ARE的激活,除了與小Maf蛋白結(jié)合外,bZIP蛋白c-Jun(AP-1轉(zhuǎn)錄因子的主要成分)和ATF4也可能與Nrf2二聚[29]。Venugopal 和Jaiswal[40-41]的研究表明,c-Jun是Nrf2激活A(yù)RE依賴性轉(zhuǎn)錄的結(jié)合伙伴,而Fos家族的成員是抑制因子。He等[42]的研究表明,ATF4可協(xié)同 Nrf2 調(diào)控ARE依賴性的Ⅱ相解毒酶和抗氧化酶基因的表達(dá)。
3.3 MetR改變小RNA(miRNAs)表達(dá)
蛋氨酸水平除了影響DNA 甲基化這種非常普遍的基因表達(dá)調(diào)控方式,還可能通過調(diào)節(jié)miRNAs實(shí)現(xiàn)營養(yǎng)表觀遺傳學(xué)調(diào)控。miRNAs屬于短鏈非編碼RNA,在動物體內(nèi)通常參與轉(zhuǎn)錄后基因沉默的調(diào)節(jié)作用。氧化應(yīng)激可以改變許多miRNAs的表達(dá)水平,但是這些變化如何與氧化應(yīng)激反應(yīng)相關(guān)卻知之甚少。
Engedal等[43]研究發(fā)現(xiàn)13種miRNA在人類細(xì)胞中受到氧化應(yīng)激的調(diào)節(jié):let-7f、miR-9、miR-16、miR-21、miR-22、miR-29b、miR-99a、miR-125b、miR-128、miR-143、miR-144、miR-155和miR-200c。利用數(shù)據(jù)庫及軟件預(yù)測了以上miRNA的共同基因靶點(diǎn)和25條共同的細(xì)胞通路,并利用功能分析得出miRNA的靶基因主要作用于(蛋白)泛素化途徑。
研究表明,miRNAs受蛋氨酸可用性的調(diào)節(jié),并可能在代謝中發(fā)揮作用[44-45]。攝入MetR日糧(0.12%)的小鼠骨髓中的miR-133a,miR-335-5p和miR-204表達(dá)增加,血漿和肝臟中的miR-31表達(dá)增加[46]。虹鱒魚攝入MetR日糧(0.775%)4周,骨骼肌中的miR-133a水平較低[47]。研究者認(rèn)為,miRNAs在細(xì)胞應(yīng)激反應(yīng)中起著不可或缺的作用,通過調(diào)節(jié)miRNAs的數(shù)量、miRNAs靶點(diǎn)的數(shù)量或miRNAs蛋白復(fù)合物的活性等方式介導(dǎo)應(yīng)激反應(yīng),恢復(fù)或合成新的體內(nèi)平衡[48]。
4 結(jié)語
蛋氨酸作為必需氨基酸,是動物生長發(fā)育不可以缺少的營養(yǎng)素。大量研究證實(shí),蛋氨酸的供給水平與機(jī)體氧化應(yīng)激狀態(tài)和健康水平息息相關(guān)。線粒體結(jié)構(gòu)和功能的變化、Nrf2、ATF4、AP-1等轉(zhuǎn)錄因子介導(dǎo)的信號通路的激活以及miRNAs的改變在MetR下氧化還原狀態(tài)的調(diào)控中起關(guān)鍵作用。MetR下線粒體以及信號通路的變化機(jī)制的探索對氨基酸營養(yǎng)調(diào)控具有重要的理論價值,同時對畜禽健康養(yǎng)殖有重要實(shí)際意義。
猜你喜歡
體育科技文獻(xiàn)通報(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
世界科學(xué)技術(shù)-中醫(yī)藥現(xiàn)代化(2020年2期)2020-07-25 02:05:56
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
西南軍醫(yī)(2016年6期)2016-01-23 02:21:19
新疆醫(yī)科大學(xué)學(xué)報(2015年10期)2015-12-26 12:33:30
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
