蛹蟲草多糖促進CETP-tg小鼠膽固醇逆轉運*
2020-11-04 06:40:24王效琰翟曉天劉伯言司艷紅秦樹存
中國病理生理雜志 2020年10期
邵 波, 李 英,, 王效琰, 翟曉天, 田 華 ,劉伯言, 張 穎 , 司艷紅△, 秦樹存,△
(山東第一醫科大學 1基礎醫學院,2動脈粥樣硬化研究所,3第二臨床學院,山東泰安271000)
動脈粥樣硬化(atherosclerosis,AS)性心腦血管疾病是臨床常見病和多發病,可引起心肌梗塞、腦卒中等并發癥,對人類造成了嚴重危害[1]。膽固醇逆向轉運(reverse cholesterol transport,RCT)是機體抗AS 的重要機制之一。盡管他汀類等藥物的應用降低了血脂,緩解了AS 的發展,卻不能逆轉動脈壁已形成的斑塊。然而,細胞膽固醇外流和RCT 可將外周組織(包括AS 斑塊)中過多的膽固醇轉運至肝臟進行再循環或以膽汁酸的形式隨糞便排出體外,是機體逆轉AS 的重要途徑[2]。因此,尋找并確證促RCT的活性成分具有重要的臨床意義。
蛹蟲草與冬蟲夏草相似,具有多種保健作用。蛹 蟲 草 多 糖(Cordyceps militarispolysaccharide,CMPS)作為蛹蟲草的主要活性成分,其功效逐漸被國內外學者重視與研究。已有報道顯示:CMPS粗品可明顯降低C57BL/6 小鼠血漿非高密度脂蛋白膽固醇(non-high-density lipoprotein cholesterol,non-HDLC)濃度,并促進外周組織膽固醇經由糞便排出,但其具體機制尚未闡明[3]。本實驗選用與人類血脂譜極為相似的膽固醇酯轉運蛋白轉基因(cholesterol ester transporter transgene,CETP-tg)小鼠作為動物模型,選用巨噬細胞源性泡沫細胞作為AS 細胞模型,評價了CMPS純品對體內外RCT的影響。
材料和方法
1 材料和儀器
RAW264.7 細胞購自中國科學院上海細胞研究所。蛹蟲草子實體購自泰安岱珍生物科技有限公司;CETP-tg 雄性小鼠由山東第一醫科大學動脈粥樣硬化研究所提供;高脂飼料購自小黍有泰(北京)生物科技有限公司;3H-膽固醇購自中國同位素公司;胎牛血清購自Gibco;DMEM 培養基和胰酶購自HyClone;BCA 蛋白濃度測定試劑盒和RIPA 裂解液購自北京索萊寶科技有限公司;總膽固醇(total cholesterol,TC)、甘油三酯(triglyceride,TG)、高密度脂蛋 白 膽 固 醇(high-density lipoprotein cholesterol,HDL-C)和低密度脂蛋白膽固醇(low-density lipoprotein cholesterol,LDL-C)檢測試劑盒購自南京建成生物工程研究所;油紅O 粉末購自Sigma;清道夫受體B1(scavenger receptor B1,SR-B1)、LDL 受體(LDL receptor,LDLR)、膽固醇7α-羥化酶A1(cholesterol 7α-hydroxylase A1,CYP7A1)、載脂蛋白A1(apolipoprotein A1,ApoA1)、三磷酸腺苷結合盒轉運體A1(ATP-binding cassette transporter A1, ABCA1)、ABCG1、肝 X 受體 α(liver X receptor α,LXRα)和過氧化物酶體增殖物激活受體γ(peroxisome proliferator-activated receptor γ,PPARγ)抗體購自Abcam;Ⅱ抗購自北京中杉金橋生物技術有限公司;氧化低密度脂蛋白(oxidized low-density lipoprotein,ox-LDL)和高密度脂蛋白3(high-density lipoprotein 3,HDL3)購自廣州奕元生物技術有限公司。
旋轉蒸發儀(Heidolph);FD8508 真空冷凍干燥機(Ilshin);液閃計數儀(Perkin Elmer);Spectra Max iD3 多功能酶標儀(Molecular Devices);CO2培養箱(Binder)。
2 方法
2.1 多糖的分離與純化 蛹蟲草子實體粉末先用95%乙醇回流脫脂2 次,沉淀晾干后用5 倍體積蒸餾水95℃提取3 次,離心并濃縮獲取蛹蟲草水提液,而后用4 倍體積無水乙醇沉淀多糖24 h,離心,沉淀物依次用無水乙醇、丙酮和乙醚各洗滌2 次,晾干,獲取粗多糖。采用Sevage 試劑去除蛋白,采用AB-8 大孔吸附樹脂去除色素,采用苯酚-硫酸法測定多糖濃度為62%,采用高效液相凝膠滲透色譜法測定多糖分子量分布并接取分子量(1 200~3 900)kD 的純化多糖用于后續實驗。
2.2 動物處理 8 周齡CETP-tg 小鼠28 只隨機分為模型組和CMPS組,每組14只,實驗前兩組小鼠體重無明顯差異。2 組小鼠每日均喂飼高脂飼料(含15.8%脂肪+1.25%膽固醇),自由飲水。CMPS 組每日灌胃予與CMPS 純品50 mg/kg,模型組給予等容積純凈水灌胃,連續給藥4 周后,每組中隨機取7 只進行同位素示蹤實驗,剩余7 只小鼠常規取材進行機制探討。
2.3 體內RCT 效率的檢測 制備3H-膽固醇培養液:ox-LDL 與3H-膽固醇37℃共孵育30 min 后加入無血漿DMEM 培養液中,其中ox-LDL 終濃度為100 mg/L,3H-膽固醇終濃度5 mCi/L。
制備3H-膽固醇標記的泡沫細胞:RAW264.7 細胞貼壁后換上述培養液培養24 h,收獲3H-膽固醇泡沫細胞,胰酶消化離心后加入預冷PBS 洗2 遍,將細胞懸浮于PBS 中,調整細胞數為1.2×1010/L。取100 μL檢測細胞內放射活性,其余置于冰上備用。
動物RCT 效率檢測:小鼠腹腔注射標記的泡沫細胞懸液,每只0.5 mL,單獨籠養48 h 后處死小鼠,測定血漿、肝、膽汁、腸壁和糞便中的放射活度。血漿放射活性檢測:于注入細胞懸液后6、24 和48 h 分別內眥靜脈取血,離心后取55 μL 血漿,加入閃爍液10 mL,液體閃爍計數(liquid scintillation count,LSC)檢測。肝臟、膽汁、腸道中的放射活性檢測:48 h 后處死小鼠,取肝臟、膽汁、腸道,組織勻漿后,用正己烷/異丙醇(3/2)萃取脂質,重復3次,收集脂質層,真空冷凍干燥,進行LSC。糞便中的放射活性檢測:分別撿取0~24 h 及24~48 h 內糞便稱重,4℃純凈水中浸泡過夜(1 g 糞便加入10 mL 純凈水),第2 天加入等量無水乙醇,樣品勻漿后取200 μL,進行LSC。計算各部位放射活性占注入體內的百分比。
2.4 細胞膽固醇外排的檢測 鼠源RAW264.7 巨噬細胞正常生長于含10%胎牛血清的DMEM 高糖培養基中。細胞實驗時,取處于對數生長期細胞,調整細胞密度至1×108/L 種于6 孔板,貼壁后改用無血清培養基。用100 mg/L ox-LDL 作用24 h 誘導細胞泡沫化,倒掉培養液,PBS 洗3 次,不同濃度CMPS(0、5、10和20 mg/L)干預6 h后,加入50 mg/L HDL3繼續培養6 h,使細胞內膽固醇流出,檢測細胞內脂質、TC水平及蛋白表達變化。
2.5 油紅O 染色 將細胞培養于放有無菌蓋玻片的6 孔培養板內,細胞經處理后,4%多聚甲醛溶液固定,PBS 潤洗后以油紅O 染色液染色15 min,蘇木素復染,水性封片劑封固,顯微鏡觀察,每張爬片隨機選取5個區域拍照。
2.6 細胞內總膽固醇的測定 RAW264.7 細胞按上述膽固醇外排處理后,每孔加入300 μL RIPA裂解細胞,離心取上清,部分上清液用于測定細胞內TC水平,部分用于提取蛋白。分別按照組織總膽固醇試劑盒說明書和BCA蛋白濃度測定試劑盒說明書檢測總膽固醇和總蛋白濃度,結果以每mg 蛋白中膽固醇的相對水平進行表示。
2.7 Western blot 分析 取小鼠血漿(檢測ApoA1)進行蛋白定量及變性,取肝組織(檢測SR-B1、LDLR和CYP7A1)和培養的巨噬細胞(檢測ABCA1、ABCG1、LXRα和PPARγ)用 RIPA 提取蛋白后進行蛋白定量及變性,按每孔50 μg加入6%~10%分離膠中,電泳轉膜,用含5%脫脂奶粉的TBS-T 封閉2 h,I抗4℃孵育過夜,TBS-T 洗膜后以相應辣根過氧化物酶標記的Ⅱ抗孵育1 h,ECL 化學發光系統顯影,采用ImageJ 軟件分析蛋白條帶積分吸光度,最終以目的蛋白積分吸光度/β-actin 積分吸光度反映蛋白表達的相對水平,以每mg 蛋白的ApoA1 蛋白條帶積分吸光度反映血漿ApoA1水平。
3 統計學處理
采用SPSS 19.0 軟件進行統計學分析。數據以均數±標準差(mean±SD)表示。兩組間比較采用獨立樣本t檢驗;多組間比較采用單因素方差分析及LSD法。以P<0.05為差異有統計學意義。
結 果
1 動物實驗發現蛹蟲草多糖純品降低了高脂飲食CETP-tg 小鼠血漿膽固醇水平,提高了體內膽固醇逆轉運效率
1.1 蛹蟲草多糖純品降低了高脂飲食CETP-tg小鼠血漿 TC、LDL-C 及 HDL-C 濃度 高脂喂飼 4 周后,CETP-tg 小鼠血漿 TC、HDL-C和LDL-C 水平分別為137.41、102.86 和49.61 mg/dL;每天灌胃給予CMPS純品后,小鼠血漿TC、HDL-C 和LDL-C 水平分別降低了24%、23%和22%;但未發現CMPS對CETP-tg小鼠血漿TG水平有顯著影響,見圖1A。
1.2 蛹蟲草多糖純品促進了高脂飲食CETP-tg小鼠體內膽固醇逆轉運過程 在CMPS 干預4 周后,小鼠腹腔注射3H-膽固醇標記的巨噬細胞源性泡沫細胞,采用同位素示蹤技術檢測了3H-膽固醇排泄的整個過程。閃爍計數儀檢測結果顯示:與模型組相比,50 mg/kg CMPS 干預后轉運至血漿中的3H-膽固醇在6 h及24 h 分別增加46%和45%;48 h 后兩組肝臟中同位素水平沒有統計學顯著性(數據未顯示);48 h 后模型組膽汁中的3H-膽固醇占注入體內總量的0.37%,CMPS 干預組則為0.52%,增加了39%;48 h后模型組腸道中的3H-膽固醇占注入總量的2.55%,CMPS 干預組則為3.89%,增加了53%;最終24 h 內對照組糞便中的3H-膽固醇含量為7.17%,CMPS 干預組為9.41%,24 h 內糞便外排增加了31%,見圖1B。而24~48 h 內2 組糞便外排量的差異沒有統計學顯著性(數據未顯示)。
1.3 蛹蟲草多糖純品上調了高脂飲食CETP-tg小鼠膽固醇逆轉運過程肝臟相關蛋白SR-B1、LDLR 及CYP7A1 的表達 Western blot 實驗結果顯示,模型組和CMPS 組血漿ApoA1 水平的差異沒有統計學顯著性;然而,與模型組相比,肝臟中SR-B1、LDLR 和CYP7A1 的蛋白表達明顯升高,分別增加了105%、71%和58%,見圖1C。
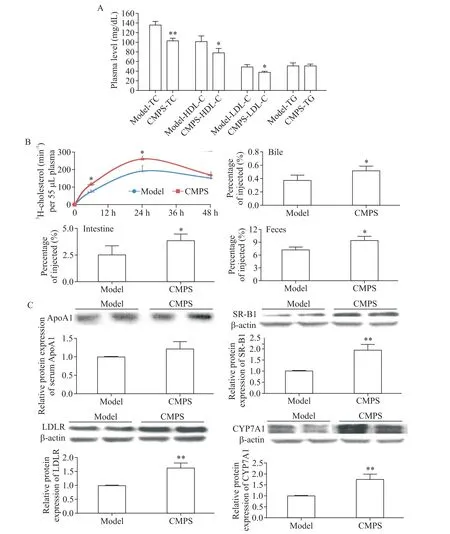
Figure 1.CMPS reduced plasma cholesterol level and promoted RCT in CETP-tg mice fed with high-fat diet.A:the effects of CMPS on plasma levels of TC,LDL-C,HDL-C and TG in CETP-tg mice.B:3H-cholesterol-labeled RAW264.7 cells were intraperitoneally injected into CETP-tg mice.At 12,24,36 and 48 h after injection,blood samples from each group were collected from retro-orbital sinus for dynamic monitoring 3H-cholesterol transport efficiency of plasma.The mice were killed after 48 h.The levels of 3H-labeled cholesterol in bile,intestine and feces from each group were also determined by liquid scintillation counting.C:the protein expression of ApoA1 in plasma and the expressions of SR-B1,LDLR and CYP7A1 in liver were detected by Western blot.Mean±SD. n=3.*P<0.05,**P<0.01 vs model group.圖1 蛹蟲草多糖純品降低了高脂飲食CETP-tg小鼠血漿膽固醇水平,提高了體內膽固醇逆轉運效率
2 細胞實驗驗證蛹蟲草多糖純品促進了巨噬細胞源性泡沫細胞膽固醇外排
2.1 蛹蟲草多糖純品降低了巨噬細胞源性泡沫細胞內脂質和TC 的水平 本實驗中,我們觀察了CMPS 純品對巨噬細胞源性泡沫細胞膽固醇流出的影響。RAW264.7 細胞吞噬ox-LDL 泡沫化后,給予50 mg/L HDL3介導膽固醇外排,在外排過程中,采用不同濃度CMPS(0、5、10和20 mg/L)進行干預。油紅O 染色及細胞內TC 檢測顯示CMPS 以劑量依賴的方式促進了膽固醇外流,見圖2。
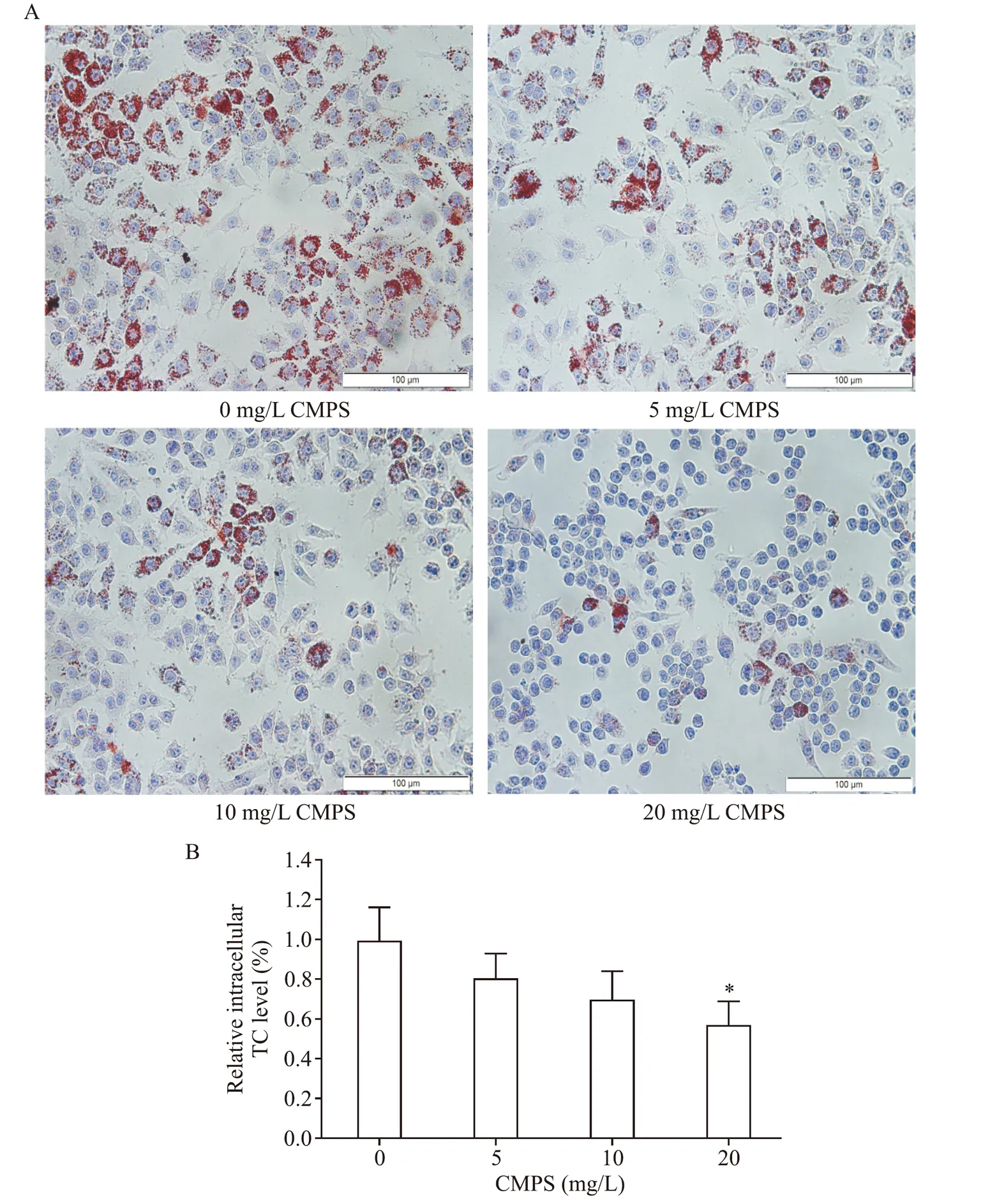
Figure 2.CMPS decreased lipid and total cholesterol(TC)levels in macrophage-derived foam cells.A:lipid droplet content was assessed using oil red O staining;B:intracellular TC content was assessed by the corresponding testing kit.Mean±SD. n=3.*P<0.05 vs 0 mg/L CMPS group.圖2 蛹蟲草多糖純品降低了巨噬細胞源性泡沫細胞內脂質和TC水平
2.2 蛹蟲草多糖純品上調了巨噬細胞中PPARγ-LXRα-ABCA1/ABCG1 信號通路相關蛋白的表達CMPS 與 RAW264.7 泡沫細胞共孵育 6 h 后,Western blot 結果表明,CMPS 處理明顯升高細胞內ABCA1、ABCG1、LXRα及PPARγ蛋白的表達,見圖3。
討 論
巨噬細胞源性泡沫細胞在動脈壁積聚是脂紋和粥樣斑塊形成的關鍵環節,然而,RCT 可以逆轉此過程。RCT 包含細胞膽固醇外流,脂蛋白運輸以及肝臟轉化與排出3 個環節,其過程簡介如下:首先,膽固醇從外周細胞(泡沫細胞)經胞膜上的ABCA1ABCG1SR-B1 轉運蛋白流出至 HDL 或ApoA1,使HDL 不斷成熟。在血漿膽固醇酯轉運蛋白(cholesterol ester transporter,CETP)介導下,HDL顆粒上的膽固醇酯可與LDL、極低密度脂蛋白(verylow-density lipoprotein,VLDL)或乳糜微粒上的甘油三酯進行交換。隨后,HDL-C 和LDL-C 隨血液循環到達肝臟,與肝細胞上相應受體SR-B1 及LDLR 結合,使過多膽固醇運回肝臟。在肝臟中,膽固醇經CYP7A1催化轉變成膽汁酸,經由膽汁進入腸道隨糞便排出[4]。本研究利用同位素示蹤技術,準確評價了體內膽固醇逆轉運和外排效率。動物實驗結果表明:CMPS 純品喂飼提高了高脂飲食CETP-tg 小鼠RCT 效率,3H-膽固醇經血漿脂蛋白轉運至肝臟,分泌入膽汁增多,最終經腸道和糞便排出體外明顯增加。同時,Western blot 結果也顯示:CMPS 可增強肝臟中膽固醇受體SR-B1、LDLR 的表達及膽固醇轉化酶CYP7A1 的表達,提示CMPS 加速了肝臟對血漿膽固醇的攝取、轉化與外排,這可能是膽汁、腸道及糞便中3H-膽固醇明顯增加的重要原因。本實驗中RCT 效率增加是CMPS 降低血漿膽固醇(TC、HDL-C和LDL-C)水平的重要機制。
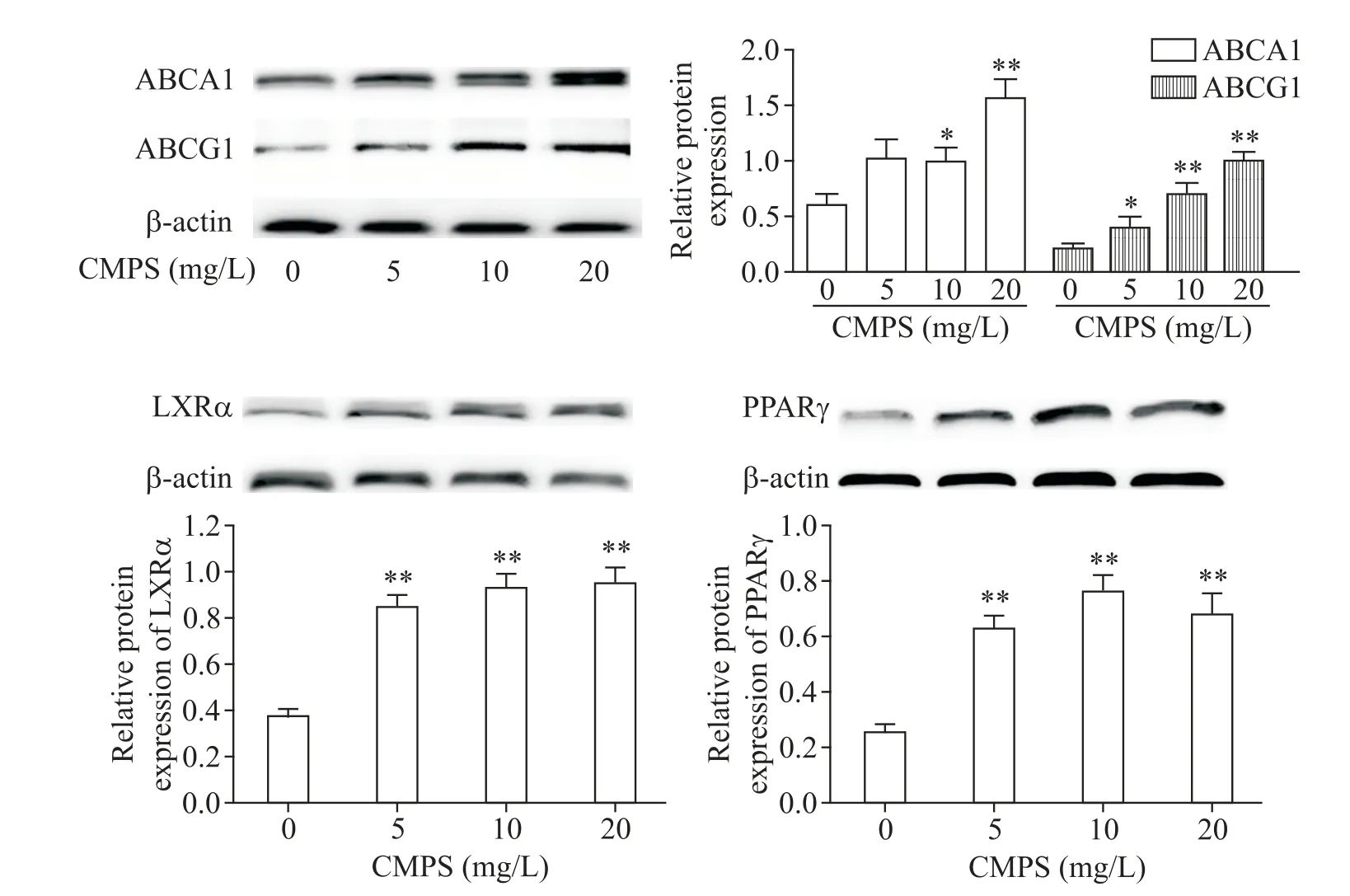
Figure 3.CMPS up-regulated the expression of PPARγ-LXRα-ABCA1/ABCG1 signaling pathway-related proteins in macrophage-derived foam cells.RAW264.7 macrophage-derived foam cells were exposed to CMPS at different concentrations(0,5,10,and 20 mg/L)for 12 h,and the protein levels of ABCA1,ABCG1,PPARγ and LXRα were determined by Western blot.Mean±SD. n=3.*P<0.05,**P<0.01 vs 0 mg/L CMPS group.圖3 蛹蟲草多糖純品上調了巨噬細胞源性泡沫細胞中PPARγ-LXRα-ABCA1/ABCG1信號通路相關蛋白的表達
外周細胞膽固醇外流是RCT第1步,也是最重要的一步,ABCA1/G1膽固醇泵對這一過程至關重要[5]。LXRα是一種轉錄激活因子,可通過激活ABC 超家族中ABCA1/G1和ABCG5/ABCG8等的表達調節糖脂代謝和維持膽固醇穩態[6]。PPARγ是 LXRα 的上游基因,屬于核受體超家族成員。PPARγ 能夠激活LXRα,進而作用于靶基因ABC 超家族介導膽固醇流出,促進RCT[7]。因此,PPARγ-LXRα-ABC 通路在巨噬細胞膽固醇外排機制中具有重要調控作用[8]。本研究中細胞實驗結果顯示CMPS干預可上調PPARγ-LXRα-ABC通路蛋白,減輕巨噬細胞源性泡沫細胞內膽固醇蓄積,促進膽固醇外排,這可能是小鼠RCT示蹤實驗中血漿3H-膽固醇明顯增加的重要原因。
CETP是血液循環中參與脂代謝與RCT過程的重要調節酶,主要介導中性脂質如膽固醇酯(cholesteryl ester,CE)和TG在HDL和含ApoB-100脂蛋白之間進行等摩爾轉移,最終將CE 從HDL 轉運至VLDL 和LDL 上,TG 則反轉至 HDL[9]。不同物種 CETP 表達差別很大,高等生物(人類和靈長類動物)血漿具有高水平CETP,倉鼠、豚鼠和兔CETP水平較低,而嚙齒類動物則沒有 CETP[10]。在人類 RCT 過程中,外周細胞外流的膽固醇與HDL 結合后,被卵磷脂-膽固醇酰基轉移酶(lecithin-cholesterol acyltransferase,LCAT)酯化轉變為CE,并通過CETP轉移給VLDL和LDL,隨后與肝細胞上LDLR 結合,被肝臟攝取與處理,這是RCT過程的重要組成部分[11]。然而,正常小鼠血漿無CETP 活性,其血脂譜與人類不同,血漿膽固醇以HDL-C 為主,non-HDL-C(包含 LDL-C 及 VLDL-C)極少,所以正常小鼠RCT過程與人類存在差異。本研究中,我們選取人CETP-tg小鼠作為實驗動物。與普通C57BL/6 小鼠相比,CETP-tg 小鼠在普通飲食和高脂飲食時其HDL-C 比例降低,non-HDL-C 比例升高,是模擬人類RCT的較好動物模型[12]。
多糖是一類分子結構復雜且龐大的糖類物質。大量研究證實,從植物、動物、真菌、海藻等自然資源中分離純化的多糖具有抗氧化、抗炎、免疫調節、降血糖、抗腫瘤等多種藥理活性[13-15]。近年來,多項動物實驗及臨床觀察表明多糖(香菇多糖[16]、海藻多糖[17]、靈芝多糖[18]等)尚具有降脂功效,然而,深入的分子機制探討的報道較少。本實驗證實CMPS 可促進高脂飲食CETP-tg 小鼠RCT 和泡沫細胞膽固醇外流,并分析了相關調節蛋白的表達,這可能是CMPS降脂的重要機制。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
現代臨床醫學(2022年4期)2022-09-29 07:38:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
昆明醫科大學學報(2021年4期)2021-07-23 01:21:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
海南醫學(2016年8期)2016-06-08 05:43:00
海峽科技與產業(2016年3期)2016-05-17 04:32:12
醫學研究雜志(2015年9期)2015-07-01 17:28:15
