旱柳F1群體速生性和冠型的群體評價
2020-11-16 02:26:44江鈺娜馮汶祥高俊峰吳瑜瑋馮志聰黃婧柯張健劉國元
安徽農業科學 2020年20期
江鈺娜 馮汶祥 高俊峰 吳瑜瑋 馮志聰 黃婧柯 張健 劉國元

摘要 林木是重要的工業原料,也是最穩定且最經濟的纖維原料。開展柳樹速生性和冠型的研究是解決我國木材原料短缺的有效手段。對195個5年樹齡的雜種F1代進行樹高、胸徑、枝角等表型的測定,通過相關分析等發現高生長和徑生長有極顯著相關性,枝角與高生長存在較顯著負相關。進一步通過主成分分析,篩選出18個窄冠速生的新品系,其中包括4個雄性株系。該研究為研究柳樹速生性和冠型的遺傳機制以及進行柳樹新品種的選育奠定了基礎。
關鍵詞 旱柳;速生;冠型;F1群體
中圖分類號 S792.12? 文獻標識碼 A? 文章編號 0517-6611(2020)20-0123-04
doi:10.3969/j.issn.0517-6611.2020.20.033
Evaluation on Fast-growth and Crown Type of F1 Population of Salix matsudana
JIANG Yu-na, FENG Wen-xiang, GAO Jun-feng et al
(College of Life Sciences, Nantong University, Nantong, Jiangsu 226019)
Abstract Trees are important industrial raw materials and the most stable and economical fiber raw materials. The research on the fast-growing and crown type of willow is important to solve the shortage of wood. In this study, tree height, DBH and branch angle of 195 F1 generations were measured. Combining the anova analysis and correlation analysis, we found that hight growth and diameter growth showed a very significant correlation. While the branch angle and hight growth showed significant negative correlation. Under the principal component analysis, we further identified 18 narrow crown and fast-growing generations, including 4 male generations. These results provided an important foundation for further studies of the genetics of fast-growing and crown type, and can be utilized for willow improvement in the future.
Key words Salix matsudana;Fast-growth;Crown type;F1 population
林業在陸地生態的保護和修復中具有十分重要的作用。林木又是重要的工業原料,也是最穩定且最經濟的纖維原料,對應對全球能源危機、氣候變化等問題有著至關重要的經濟和環境效益 [1]。作為全球第二大木材消耗國和第一大木材進口國,我國木材主要靠進口,木材短缺已嚴重制約我國經濟發展。統計數據表明,近10年我國進口木材資源比重由38%增加到58%,僅2018年1年,我國原木材進口量就已高達5 968.6萬m3,鋸材進口量達3 673.6萬m3(中國林業網)。我國現有的林業資源已遠遠不能滿足社會需求,增加林木的速生性就成為了解決我國木材短缺的有效途徑。
柳樹是我國原生樹種,也是優良的人工林樹種,由于喬木柳生長快,抗病蟲害能力強,又具有喜光、喜濕、耐寒、耐旱、耐鹽、抗風、易繁殖、成活率高的特點,在我國東北、華北、西北等溫帶地區都有大面積分布,在沿海等土壤質量較差的地方也有廣泛種植。喬木柳與楊樹、桉樹這些常見人工林樹種相比,具有木材密度、抗壓、抗剪能力強于楊樹,分布性、適應性、對生態環境副作用優于桉樹等特點。喬木柳是優質的工業材料,被廣泛利用在制漿 [2-4]、造紙、建筑器具、人造棉、火藥、刨花板 [5]、重組木 [6]、中密度纖維板 [7]等方面。因此,改良喬木柳的速生性,增加其高生長和徑生長,不僅有利于柳樹的遺傳改良,也對解決我國工業原料短缺的問題有重要作用。
速生性是受許多形態、解剖和生理生化特性控制的復合遺傳性狀,它是既受栽培條件的影響,又受光合作用、生長期、干重、苗高、基徑、激素響應等多個因素影響的綜合指標 [8]。其中高生長和徑生長是描述林木速生性的2個主要方面,高生長的衡量指標是樹高,徑生長的衡量指標有基徑、胸徑等 [9-11]。
在實際生產中,農田林網和農林間作時,柳樹會造成農作物減產 [12],推廣窄冠型旱柳樹種可以保證一定的木材需求和糧食產量的供應。且窄冠型柳樹也常當作行道樹等進行園林綠化。目前對窄冠型樹種的研究主要集中在農田防護、間作效益、林糧間作技術、透光特性等方面。已有研究發現,推廣窄冠型樹種既可增加糧食產量,又可增加林木的生產量。劉振廷等 [13]研究發現窄冠黑楊與小麥間作可有效降低麥田氣溫,提高小麥年產量。選用窄冠白楊進行糧林間作可以使小麥增產約 225 kg/hm2 [14]。窄冠型蘋果樹的產量是寬冠幅產量的3倍 [15]。窄冠型楊樹的推廣應用除可提高糧林產量外,還具有冠幅窄、生長快、木質好、多為雄株無飛絮污染、根系深、耐鹽堿、樹型高大美觀等優點 [16]。
植物分蘗角度或分枝角度是重要的農藝性狀,它影響植物的外觀形態和生長發育,是植物形態結構的重要組成部分 [17]。合理的冠型可以提高光合利用、增加農藝產量、影響景觀植物的應用。植物分枝角度或分蘗角度發育受到環境因素和基因遺傳因素的共同調控 [18-19]。在一定的環境條件下可以通過修剪等方法來改變樹木的冠型。但植物外在形態結構主要是由遺傳因素調控的,不同植物種類擁有獨特的分枝發育程序,并且表現出獨特的表型。目前已有許多研究者對分枝角度進行了相關研究,植物激素是調控植物分枝角度或分蘗角度的主要因子。生長素含量可以影響植物側芽的生長發育 [20],并參與植物頂端優勢的形成 [21]。施用細胞分裂素可以促進小麥側芽的形成 [22]。抑制獨腳金內脂的合成可以使菊花表現出直立緊湊的外觀 [23]。除植物激素外,許多相關基因也被報道與分枝角度相關。水稻中PIN2基因可以調控生長素的極性運輸,影響其分蘗角度 [24];MOCL基因可以調控水稻中分蘗的角度 [25];TAC1基因的上調表達可以導致水稻分蘗角度變大 [26];ZmTAC1基因的突變可以導致玉米葉片角度變小,植株直立緊湊 [27]。蘋果樹中Co基因可以調控其冠型 [28]。
然而,這些研究主要是分枝角度的調控機理,在品種選育角度以及冠型與速生性關系的研究方面還很少,在柳樹中這類研究更是鮮有報道。該研究為篩選適宜在我國東部沿海人造林種植的新品系,利用江蘇地區常見旱柳品種(沿江柳)和速生性好、冠型緊湊的旱柳品種(9901)進行雜交,對F1子代的樹高、胸徑、枝角等性狀進行群體評價,初步明確冠型和速生性之間的遺傳關系,并篩選速生、窄冠的柳樹新品系,為柳樹新品種的選育提供支持。
1 材料與方法
1.1 材料
以在江蘇沿江地區常見的四倍體旱柳品種(沿江柳)為母本,以速生性好、冠型緊湊的旱柳品種(9901)作為父本,進行雜交,得到195個子代并于2014年秋季種植在江蘇省南通市沿江地區農業科學研究所柳樹資源圃基地。生長量是林木育種的重要性狀,它與冠層結構、每年快速生長期的持續時間都有很大關系,其中分枝夾角不僅能影響冠層結構,還會影響樹木的外部形態。研究冠型和生長量的關系和變異可以為柳樹的育種提供依據。當樹齡達到4年和5年時,在落葉期(11月)對群體中的子代進行樹高、胸徑、枝角等性狀的物候調查。
1.2 速生性調查和計算
首先對195株子代進行性別調查,對群體中的子代進行樹高、胸徑測量,胸徑按照主干離地面高度1.3 m的位置進行測量。對樹高年生長量和胸徑年生長量進行計算。
1.3 枝角測量
對75個株系中每個株系前三樹枝角度與主干的夾角進行測量,并以此計算每個子代的平均分枝角。
1.4 數據處理
對所得的表型數據利用Excel、R語言等軟件進行描述性統計分析、方差分析、相關分析和主成分分析。
2 結果與分析
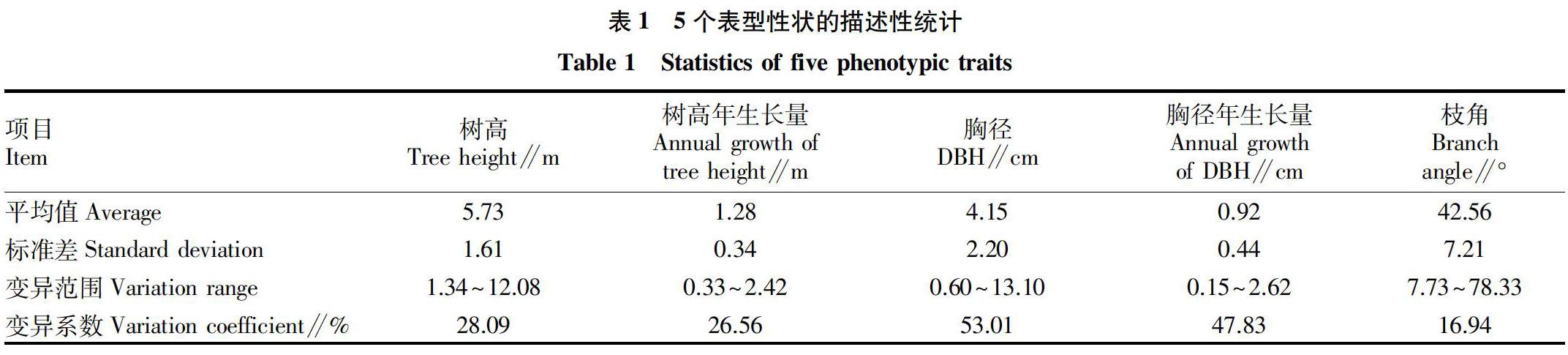
2.1 樹高、胸徑、冠型的描述性統計
首先對195個子代進行性別調查統計,其中有75個子代為雄性,102個雌性,還有18個生長過于緩慢,未能調查清楚其性別。對195個子代生長第4年和第5年時的樹高、胸徑和枝角進行統計,并分別計算樹高年生長量、胸徑年生長量和枝角的平均值。該群體中樹高為1.34~12.08 m,樹高年生長量為0.33~2.42 m,胸徑為0.60~13.10 cm,胸徑年生長量為0.15~2.62 cm。子代的平均枝角為17.73°~78.33°(表1)。各性狀變異幅度也很大,5個性狀的變異系數為16.94%~53.01%,其中胸徑的變異幅度最大。這一結果表明,195個子代在高生長、徑生長以及枝角這幾個性狀上表型變異豐富,利于開展柳樹高生長、徑生長、冠型研究。
2.2 F1群體速生性狀的評價
高生長和徑生長是評價林木速生性的2個重要指標,根據2次調查得到的樹高年生長量和胸徑年生長量可以發現,第2次調查得到的樹高年生長量和胸徑年生長量顯著高于第1次,結果表明該F1群體第5年在高生長和徑生長方面相較于以前都有較快增加。為了進一步探究高生長、徑生長的關系,對樹高、樹高年生長量、胸徑和胸徑年生長量進行相關分析。從表2可以看出,這4個性狀相互之間存在極顯著正相關,由此可以看出,在該群體中高生長和徑生長存在著很高的一致性,即高的子代相應胸徑也更粗。由于高生長和徑生長2個性狀存在很強的正相關性,這也就為選擇更高、更粗的速生新品系提供了理論 依據。
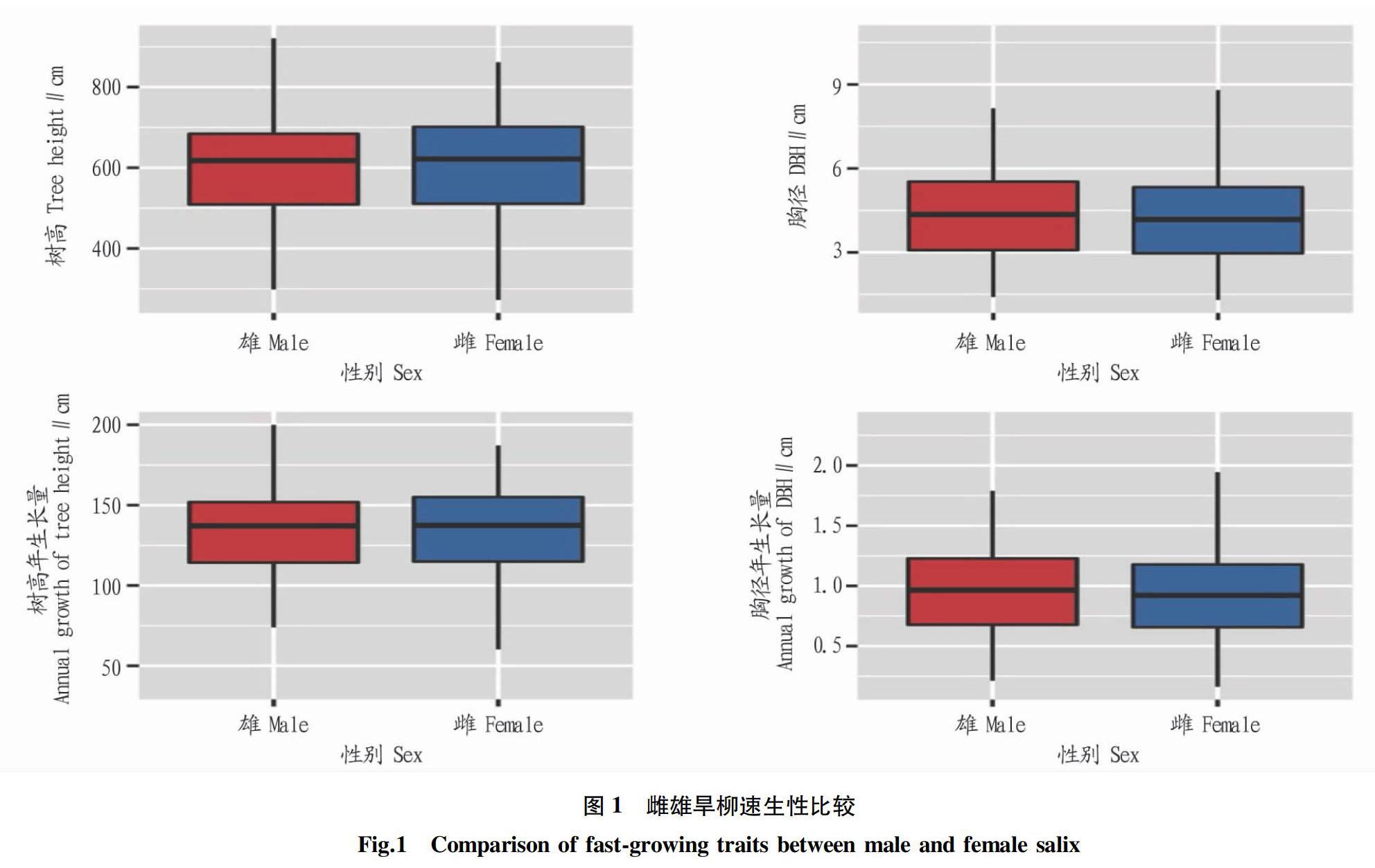
旱柳為雌雄異株的樹種,進一步對不同性別旱柳的速生性狀進行了比較和t檢驗。從圖1可以看出,雄性旱柳樹高和樹高年生長量的平均水平略低于雌性旱柳,胸徑和胸徑年生長量略高于雌性旱柳。通過進一步t檢驗,旱柳性別對這4個速生性狀的影響不大,沒有達到顯著水平。
2.3 冠型與速生性的關系
冠層結構是影響植物光合作用強弱的一個重要因素,冠型的差異也是影響景觀植物的一個重要因素。旱柳常用于人工速生林以及城市綠化的行道樹等,這些用途決定了其冠型不能過于松散。前面已對該群體的枝角進行了測量和統計,發現其變異幅度很大。由于冠層是影響光合能力的一個重要因素,而光合能力又是影響速生性的一個主要因素,為了進一步探究冠型和速生性的關系,對枝角和樹高、樹高年生長量、胸徑以及胸徑年生長量進行了相關分析。相關分析結果表明,枝角的大小與樹高和樹高年生長量存在極顯著負相關,與胸徑年生長量存在顯著負相關,然而與胸徑沒有表現出顯著相關性。這一結果反映出枝角對速生性是有很大影響的,枝角越小,樹高越高、高生長和胸徑年生長量也越大。較小枝角更有利于旱柳的速生。在種植密度達到一定程度時,枝角的增加并不能增加冠層的有效光合面積,相反還會消耗其光合產物,對速生性產生不利影響,而枝角越小,則旱柳的冠型也就越窄,窄冠型能有效增加對光能的利用率,更加利于加快旱柳的生長。
進一步分析了不同性別旱柳的冠型是否存在差異,由圖2a可以看出,雌性旱柳的枝角是略小于雄性的,通過t檢驗分析,發現兩者的枝角差異達到了顯著水平。進一步對枝角性狀進行了分析,為了便于統計,首先把該群體按照枝角 <36°,36°~ 48°,>48°的標準劃分成窄冠型、中間型以及寬冠型三類,并統計各類中雌雄株的個數。通過圖2b發現雌株的窄冠型個體的數量和比例都顯著高于雄株,而雄株的寬冠
型個體數量和比例相對較多。枝角性狀是由控制性別的基因引起的還是與其存在連鎖關系有待于進一步驗證,可以確定的是該群體中寬冠型的雄性旱柳較多,窄冠型的雌性旱柳較多。
為了進一步分析冠型對速生性的影響,對樹高年生長量和胸徑年生長量進行了方差分析。通過方差分析可以看出,冠型對樹高的年生長量存在顯著影響,對胸徑年生長量的影響未達到顯著水平。進一步進行多重比較發現(表3),3種冠型的樹高生長量均存在顯著差異,其中窄冠型的平均樹高年生長量最高,寬冠型的最低。這一結果與前面相關分析的結果也是一致的,枝角越小,冠型越小,其速生性就越高。
2.4 速生性和冠型的主成分分析
為了對該群體的速生性和冠型進行進一步評價,并進一步篩選優異株系,對調查的5個性狀進行了主成分分析,共找到了5個主成分,其中前2個主成分的方差貢獻率較高(圖3a),保留前2個主成分,對主成分進行降維,并計算5個表型性狀與2個主成分的相關系數(表4)。根據所得結果,發現PC1與4個速生表型的相關系數都達0.9以上,表明PC1與速生性有顯著正相關,PC1與枝角的相關系數為負值,表明該主成分同時與枝角大小呈負相關。PC2與速生性的相關系數不大,而與枝角的相關系數很大,達0.96,表明第二主成分主要與枝角相關。2個主成分對5個表型性狀的累計變異解釋率達90%以上,表明這2個主成分能很好地對速生性和冠型進行解釋。
對F1群體的所有子代進行了主成分分析,根據主成分分析的結果,可以對速生性好且冠型優異株系進行進一步的篩選(圖3b)。根據圖3b中紅圈部分,初步篩選出18個速生性好且窄冠型的品系。它們都是利于密植且生長量較高的品系,后續可以通過扦插進行無性繁殖,并進一步進行田間重復試驗,為新品種的選育提供種質資源。這些新品系中有4個雄株,還有14個為雌株。在實際生產應用過程中,當柳樹作為行道樹等城市綠化植物時,為了避免柳絮的污染,要盡量避免雌株種植。該研究選取的這些優異株系中的4個雄性速生窄冠品種將為園林綠化樹種的選育提供優異的種質資源。
3 討論與結論
柳樹由于其速生性好,是我國主要的速生林木之一,其抗逆性強的特點又使得其在全國各地都有很大面積的種植,尤其是東部沿海灘涂等有著大面積的人工林。因此選育速生性好、適應密植的新品種是緩解我國木材短缺的有效途徑。該研究利用在東部沿海地區大面積分布的沿江柳品種和速生性好的9901柳雜交,通過對195個子代的速生性和冠型進行調查、相關分析、方差分析等,對該群體的速生性和冠型進行了系統評價,發現高生長和徑生長有極顯著相關性,枝角與高生長存在較顯著負相關。并利用主成分分析篩選出18個窄冠速生的新品系,其中包括4個雄性株系,為柳樹的品種選育提供了優異的種質資源。
參考文獻
[1] KRAMER P J.The role of physiology in forestry[J].Tree physiology,1986,2:1-16.
[2] 苗婷婷,吳中能,劉俊龍,等.竹柳等4個柳樹無性系造紙材性測定分析[J].甘肅林業科技,2014,39(2):53-56.
[3]
袁志平.迎接竹柳造紙的新時代[J].天津造紙,2015,37(4):8-10.
[4] 郭泰.竹柳化學組分分析及制漿性能的研究[D].濟南:齊魯工業大學,2016.
[5] 董葛平,鄧玉和,王新洲,等.竹柳材性及其刨花板制造工藝研究[J].西南林業大學學報,2013,33(3):92-96.
[6] 吳金絨,鄧玉和,侯天宇,等.竹柳枝丫材性能及重組木制造[J].浙江農林大學學報,2014,31(6):947-953.
[7] 何爽爽,王新洲,鄧玉和,等.竹柳枝丫材與小徑材中密度纖維板的研究[J].林產工業,2015,42(11):31-35.
[8] 孔紅嶺.楊樹無性品種速生機理及早期選擇的研究[D].泰安:山東農業大學,2008.
[9] 吳小龍.南酸棗生長進程研究[J].江蘇林業科技,2003,30(6):6-8,19.
[10] 范輝華.杉木、擬赤楊混交對杉木持續生長的影響[J].林業科學研究,2001,14(4):455-458.
[11] 蘇雪輝,趙自成.楊樹新無性系造林生長節律觀測初報[J].江蘇林業科技,2003,30(4):10-12.
[12] 尹建道,吳春森,李慶臣,等.農用型窄冠黑楊、窄冠黑白楊新品種引種試驗[J].東北林業大學學報,2007,35(11):7-8.
[13] 劉振廷,郭延凱,王書軍,等.窄冠白楊株行距配置對間作小麥產量的影響[J].河北林業科技,2000(1):8-10.
[14] 盧勝西,劉國興,朱西存,等.窄冠白楊間作效益的研究[J].山東林業科技,1995(5):32-36.
[15] JACOB H B.Breeding experiments of apple varieties with columnar growth and low chilling requirements[J].Acta horticulturae,2010,872:159-164.
[16] 龐金宣,張友朋,李際紅,等.窄冠黑楊窄冠黑白楊的選育(I)田間試驗及結果分析[J].山東林業科技,2003(5):1-7.
[17] STEEVES T A,SUSSEX I M.Patterns in plant development[M].Cambridge,UK:Cambridge University Press,1989:124-146.
[18] SHIMIZU-SATO S,MORI H.Control of outgrowth and dormancy in axillary buds[J].Plant Physiol,2001,127(4):1405-1413.
[19] BEVERIDGE C A,WELLER J L,SINGER S R,et al.Axillary meristem development.Budding relationships between networks controlling flowering,branching,and photoperiod responsiveness[J].Plant Physiol,2003,131(3):927-934.
[20] TAKEDA T,SUWA Y,SUZUKI M,et al.The OsTB1 gene negatively regulates lateral branching in rice[J].Plant journal,2003,33(3):513-520.
[21] HUBBARD L,MCSTEEN P,DOEBLEY J,et al.Expression patterns and mutant phenotype of teosinte branched1 correlate with growth suppression in maize and teosinte [J].Genetics,2002,162:1927-1935.
[22] 蔡鐵,徐海成,尹燕枰,等.外源 IAA、GA3和ABA 影響不同穗型小麥分蘗發生的機制[J].作物學報,2013,39(10):1835-1842.
[23] 郗琳.獨角金內脂在缺磷條件下影響菊花側枝伸長的研究[D].北京:中國農業大學,2015.
[24] 趙亞波,卜賢盼,李培綱,等.OsPIN1a 基因在水稻植株不同組織表達與定位研究[J].生物技術,2014,24(1):27-32.
[25] LI X Y,QIAN Q,FU Z M,et al.Control of tillering in riec[J].Nature,2003,422:618-621.
[26] ZHAO H,HUAI Z X,XIAO Y J,et al.Natural variation and genetic analysis of the tiller angle gene MsTAC1 in Miscanthus sicensis[J].Planta,2014,240:161-175.
[27] KU L X,WEI X M,ZHANG S F,et al.Cloning and characterization of a putative TAC1 ortholog associated with leaf angle in maize(Zea mays L.)[J].PLoS One,2011,6(6):1-7.
[28] MORIYA S,OKADA K,HAJI T,et al.Fine mapping of Co,a gene controlling columnar growth habit located on apple(Malus × domestica Borkh.)linkage group 10[J].Plant breeding,2012,131(5):641-647.
