IRS-1/TAZ信號通路對骨髓脂肪化的影響及機制
2020-12-02 04:20:48WANGXiao李玉坤劉斯靜
河北醫科大學學報 2020年11期
王 娜,劉 暢,薛 鵬,WANG Xiao,李玉坤,劉斯靜
(1.河北醫科大學第三醫院內分泌科,河北省骨科生物力學重點實驗室,河北 石家莊 050051;2.約翰·霍普金斯醫學院肌肉與骨骼研究中心,美國 馬里蘭 21212;3.河北醫科大學期刊社,河北 石家莊 050017)
成骨細胞和脂肪細胞共同來源于骨髓間充質干細胞(bone marrow derived mesenchymal stem cells,BMSCs),因此其分化過程存在競爭關系。脂肪細胞占骨髓細胞的15%~70%,疾病狀態下(如骨質疏松癥)或者隨著年齡增長,脂肪細胞數目增多,替代正常的其他類型細胞分化,如成骨細胞分化[1]。胰島素受體底物1(insulin receptor substrate 1,IRS-1)是胰島素信號通路的樞紐因子,可以通過激活下游通路的一系列信號轉導,完成對細胞增殖、凋亡、分化等功能的調節[2]。目前,關于IRS-1對成脂分化的作用尚存爭議。一項動物實驗表明,敲除IRS-1基因可使成脂分化呈進行性降低[3]。Kim等[4]研究發現,過表達IRS-1改善脂肪細胞脂質儲存,即IRS-1促進脂質合成。具有盤狀同源區域結合序列的轉錄共活化因子(transcriptional coactivator with PDZ-binding motif,TAZ)是一種調節干細胞增殖、分化等功能的重要轉錄調控因子[5]。Byun等[6]研究表明,TAZ一方面通過共活化Runt相關轉錄因子2(Runt related transcription factor 2,RUNX2)促進BMSCs向成骨細胞方向分化,另一方面通過結合過氧化物酶體增生物激活受體γ(Peroxisome proliferator activated receptorγ,PPARγ)抑制成脂分化相關基因轉錄,抑制BMSCs向脂肪細胞分化。TAZ是否還有其他機制并不明確,且其所受調控的上游信號通路并未完全闡明[7]。本研究旨在探索IRS-1/TAZ信號通路調控BMSCs向成脂細胞分化的潛在機制,為糖尿病發病候選基因IRS-1參與調控骨代謝和脂肪代謝提供新依據。
1 材 料 與 方 法
1.1實驗動物 3月齡SPF級雌性未受孕SD(Sprague Dawley)大鼠15只用于動物實驗;另4周齡SD大鼠9只用于BMSCs的原代培養。實驗動物購于河北省實驗動物中心,動物合格證書號 1708080。
1.2方法
1.2.1動物分組及骨質疏松模型的構建 將15只3月齡SD大鼠隨機分為對照組、假手術組和去卵巢手術(ovariectomy,OVX)組,每組5只。去卵巢手術操作步驟:大鼠禁食禁水12 h后進行麻醉,俯臥位定位大鼠卵巢,消毒手術部位,依次剪開皮膚、肌肉、筋膜,分離脂肪團,手術結扎輸卵管,完整切除雙側卵巢,止血并逐層縫合。假手術組僅切除卵巢等體積脂肪組織,保留卵巢。術后3 d連續應用抗生素預防感染。術后12周用于后續實驗。
1.2.2大鼠BMSCs的原代培養及成脂分化 將4周齡SD大鼠CO2麻醉后處死,用75%乙醇浸泡5 min。在無菌條件下取雙側股骨,剪除股骨近端骨骺,用5 mL注射器吸取DMEM培養基反復沖洗骨髓腔。細胞取出后,以5×106/cm2的密度接種于含12%胎牛血清及1%青鏈霉素的DMEM培養基中,放入細胞培養箱中培養。原代細胞達到80%融合后傳代,P3代細胞用于后續實驗。應用成脂誘導培養基(DMEM含12%胎牛血清,1%青鏈霉素,10 g/L胰島素,500 mmol/L甲基異丁基黃酸鈉和1 μmol/L地塞米松)誘導BMSCs向脂肪細胞分化。
1.2.3組織化學染色 取SD大鼠股骨,用4%多聚甲醛固定,4 ℃,24 h;0.5 mol/L乙二胺四乙酸脫鈣3周,4 ℃,搖床;酒精梯度脫水后石蠟包埋,制作4 μm石蠟,切片。將石蠟切片65 ℃烘干,二甲苯脫蠟,酒精梯度復水。采用蘇木精和伊紅(hematoxylin and eosin,HE)染色的標準步驟進行染色。顯微鏡下對脂肪細胞進行計數并拍照。
1.2.4免疫熒光分析 取SD大鼠股骨,用4%多聚甲醛固定,4 ℃,24 h;0.5 mol/L乙二胺四乙酸脫鈣3周,4 ℃,搖床;應用8%明膠包埋,制作20 μm冰凍切片。將冰凍切片37 ℃烘干,浸泡于PBS液15 min,5% BSA封閉1 h,一抗孵育(抗圍脂滴蛋白,Sigma),4 ℃,過夜;TBST清洗,熒光二抗孵育1 h,TBST清洗,封片。顯微鏡下對熒光標記細胞進行計數并拍照。
1.2.5酶聯免疫吸附測定(enzyme-linked immunosorbent assay,ELISA) 取SD大鼠股骨,將周圍組織剔除干凈,于液氮條件下研磨,加入RIPA裂解液,超聲破碎后離心,取上清液。根據IRS-1 ELISA試劑盒(武漢菲恩生物科技有限公司)操作步驟測定IRS-1濃度。
1.2.6質粒轉染 課題組在前期實驗中已經自行構建IRS-1高表達質粒(pCMV-IRS-1)[8]。本研究構建了分別用于敲除IRS-1和TAZ表達的質粒,即嵌入可敲除IRS-1表達的小干擾RNA(SiIRS-1)的質粒,以及嵌入可敲除TAZ表達的小干擾RNA(SiTAZ)的質粒(上海吉凱基因化學有限公司)。應用脂質體3 000(Thermo)對BMSCs進行轉染。細胞轉染24 h后,更換為成脂誘導培養基進行培養。同時設立對照組,未進行質粒轉染組設置為“空白對照組”,轉染空白質粒組作為“陰性對照組”。
1.2.7油紅O染色 將細胞接種至35 mm塑料培養皿中成脂誘導14 d后,PBS清洗2次,4%多聚甲醛室溫固定20 min,蒸餾水清洗3次。將油紅O儲存液(0.5 g油紅O溶于100 mL異丙醇)與蒸餾水以3∶2的比例配置工作液,室溫放置10 min后過濾。應用油紅O工作液室溫染色15 min。蒸餾水清洗3次后,顯微鏡下拍照。然后,應用異丙醇對油紅O進行脫色,加入等體積PBS液后放入通風櫥,室溫,30 min。應用酶標儀(520 nm)讀取其OD值。
1.2.8蛋白免疫印跡(Western blot) 將細胞接種至60 mm塑料培養皿中成脂誘導3 d后,利用RIPA裂解液提取蛋白。應用12% SDS-PAGE膠對蛋白進行電泳分離后,采用半干法將其轉移至PVDF膜。5%牛奶封閉,37 ℃,室溫。一抗孵育(抗TAZ,Cell Signaling;抗CCAAT/增強子結合蛋白β(CCAAT/enhancer binding protein β,C/EBPβ),Boster;抗PPARγ,Boster;抗GAPDH,Bioworld),4 ℃,過夜。二抗37 ℃孵育1 h。分析蛋白條帶灰度值,并計算目的蛋白與內參蛋白條帶灰度值的比值。
1.3統計學方法 應用SPSS 20.0統計軟件分析數據。所有實驗至少重復3次。計量資料比較采用單因素方差分析和SNK-q檢驗。P<0.05為差異有統計學意義。
2 結 果
2.1不同組別大鼠股骨內脂肪細胞計數、熒光標記細胞計數、IRS-1濃度比較 對大鼠股骨進行HE染色,計數脂肪細胞,結果發現,3組間脂肪細胞計數差異有統計學意義(P<0.05),OVX組脂肪細胞計數多于對照組和假手術組,差異有統計學意義(P<0.05),對照組和假手術組脂肪細胞計數差異無統計學意義(P>0.05)。利用免疫熒光染色對脂肪細胞標記物圍脂滴蛋白進行標記并計數,分析結果與HE染色結果趨勢一致。ELISA法測定大鼠股骨內IRS-1濃度,結果發現,3組間IRS-1濃度差異有統計學意義(P<0.05),OVX組IRS-1濃度低于對照組和假手術組,差異有統計學意義(P<0.05),對照組和假手術組IRS-1濃度差異無統計學意義(P>0.05)。見圖1,表1。
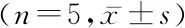
表1 不同組別大鼠股骨內脂肪細胞、熒光標記細胞計數和IRS-1濃度比較
2.2IRS-1基因高表達對油紅O染色結果和成脂分化相關蛋白表達的影響 BMSCs成脂誘導14 d后,對其進行油紅O染色,結果發現,應用pCMV-IRS-1轉染后,IRS-1組BMSCs的脂滴明顯少于空白對照組和陰性對照組(圖2A)。進一步對油紅O吸光度進行定量分析后發現,3組油紅O吸光度差異有統計學意義(P<0.05),IRS-1高表達組吸光度值低于空白對照組和陰性對照組,差異有統計學意義(P<0.05),空白對照組和陰性對照組吸光度值差異無統計學意義(P>0.05)。BMSCs成骨誘導3 d后,采用Western blot方法對成脂分化相關蛋白TAZ、C/EBPβ、PPARγ的表達水平進行分析,結果發現,IRS-1組TAZ的表達量明顯高于空白對照組和陰性對照組,C/EBPβ、PPARγ的表達量明顯低于空白對照組和陰性對照組(圖2B)。進一步對蛋白條帶灰度值進行定量分析后發現,3組TAZ、C/EBPβ、PPARγ的表達量差異有統計學意義(P<0.05), IRS-1組TAZ的表達量高于空白對照組和陰性對照組,C/EBPβ、PPARγ的表達量低于空白對照組和陰性對照組,差異有統計學意義(P<0.05),空白對照組和陰性對照組TAZ、C/EBPβ、PPARγ的表達量差異無統計學意義(P>0.05)。見表2。

表2 IRS-1基因高表達對油紅O染色和TAZ、CEBPβ、PPARγ表達影響的定量分析
2.3IRS-1或TAZ基因敲除對油紅O染色結果和成脂分化相關蛋白表達的影響 油紅O染色結果提示,SiIRS-1組和SiTAZ組脂滴明顯多于空白對照組和陰性對照組(圖3A)。進一步對油紅O吸光度進行定量分析后發現,4組間油紅O吸光度差異有統計學意義(P<0.05),SiIRS-1組和SiTAZ組吸光度值高于空白對照組和陰性對照組,差異有統計學意義(P<0.05), SiIRS-1組和SiTAZ組、空白對照組和陰性對照組吸光度值差異無統計學意義(P>0.05)。Western blot結果顯示,SiIRS-1組和SiTAZ組TAZ的表達量明顯低于空白對照組和陰性對照組,C/EBPβ、PPARγ的表達量明顯高于空白對照組和陰性對照組(圖3B)。進一步對蛋白條帶灰度值進行定量分析后發現,4間組TAZ、C/EBPβ、PPARγ的表達量差異有統計學意義(P<0.05),SiIRS-1組和SiTAZ組TAZ的表達量低于空白對照組和陰性對照組;C/EBPβ、PPARγ的表達量高于空白對照組和陰性對照組,差異有統計學意義(P<0.05)。SiIRS-1組和SiTAZ組、空白對照組和陰性對照組TAZ、C/EBPβ、PPARγ的表達量差異無統計學意義(P>0.05)。見表3。
表3 IRS-1或TAZ基因敲除對油紅O染色和TAZ、CEBPβ、PPARγ表達影響的定量分析
3 討 論
在骨髓微環境中,干細胞除了向成骨細胞分化,還可以向其他類型細胞分化。在干細胞定向分化系統中,成骨細胞和成脂細胞來源于共同的祖細胞,成骨分化和成脂分化之間的信號轉導聯系最為密切[9]。若BMSCs向成脂細胞分化,則成骨分化信號轉導受到抑制,它們之間相互競爭以保持骨髓中不同細胞類型的穩定增長[10]。因此,骨質疏松癥患者,尤其是在老年人群,常常伴隨骨髓脂肪細胞蓄積。
IRS-1是激素受體后錨定蛋白,能結合下游特定靶分子,進而參與調控多種細胞代謝過程,脂代謝便是其中之一[11]。本研究采用大鼠BMSCs作為研究對象,油紅O染色及半定量分析結果表明,過表達IRS-1則BMSCs向成脂細胞分化被抑制,敲低IRS-1則BMSCs向成脂細胞分化增多。既往研究發現,轉錄共活化因子TAZ可與成脂分化關鍵轉錄因子PPARγ結合,減少其下游靶基因轉錄,從而抑制成脂分化進程[12-13]。作為YAP類似物,TAZ第89位絲氨酸位點被磷酸化后,與14-3-3蛋白結合,將復合體阻滯于細胞質,使復合體進一步蓄積,從而發揮相應信號轉導或信號組織作用。目前研究表明,TAZ可能受上游多條信號通路的調控,包括TGFβ、BMP、Wnt等信號通路[14]。可見,胰島素信號通路的樞紐分子IRS-1也是TAZ的上游信號,可通過靶向調控TAZ表達影響BMSCs向成脂細胞分化的進程。
本研究結果顯示,過表達IRS-1則成脂分化標志物C/EBPβ、PPARγ表達下調,敲低IRS-1和TAZ則成脂分化標志物C/EBPβ、PPARγ表達上調。C/EBPs蛋白家族表達于多種細胞類型,廣泛參與調控細胞增殖、凋亡、分化等生命過程[15]。目前已發現6個C/EBPs蛋白家族成員:C/EBPα、C/EBPβ、C/EBPγ、C/EBPδ、C/EBPε和C/EBPζ(又稱為CHOP-10)。其中,C/EBPβ和C/EBPδ是脂肪細胞分化早期標志物,在成脂分化誘導劑處理后,脂肪細胞前體中C/EBPβ、C/EBPδ表達即可達峰。更重要的是,即使不添加成脂誘導劑,C/EBPβ也可促使脂肪細胞前體進入脂肪細胞分化程序。另一方面,PPARs是一類配體激活核轉錄因子超家族,包括3種亞型:PPARα、PPARβ和PPARγ[16]。在成脂分化早期,活化的C/EBPβ和C/EBPδ將進一步激活C/EBPα和PPARγ。C/EBPα和PPARγ不僅促進自身表達,同時還能促進相互表達,并且對大部分脂肪細胞分化的特異性基因(如aP2、GLUT4等)的表達是必不可少的[17]。本研究結果顯示,過表達IRS-1引起C/EBPβ、PPARγ表達下調,說明成脂分化信號轉導部分阻斷。反之,敲低IRS-1和TAZ引起C/EBPβ、PPARγ表達上調,說明成脂分化信號轉導增強。可見,IRS-1/TAZ信號通路可通過C/EBPβ、PPARγ調控BMSCs成脂分化進程。
綜上所述,IRS-1通過上調TAZ表達,抑制BMSCs向脂肪細胞分化。因此,IRS-1/TAZ信號可作為控制骨質疏松癥骨髓脂肪化的重要靶點。(本文圖見封三)
