樟葉越桔嫩枝1株內生真菌的鑒定及抑菌活性測定
2020-12-04 08:06:58嚴冬曾為林羅旭璐陳肖學劉惠民趙平
生物技術通報 2020年11期
嚴冬 曾為林 羅旭璐 陳肖學 劉惠民 趙平,3
(1.西南林業大學西南地區林業生物質資源高效利用國家林業和草原局重點實驗室,昆明650224;2.西南林業大學西南山地森林資源保育與利用教育部重點實驗室,昆明650224;3.云南森林資源培育與利用協同創新中心,昆明650224)
植物內生真菌是一類在健康植物組織內定殖,但不引起病害癥狀的真菌,并與宿主建立復雜的相互作用[1-2]。大量研究表明,許多經濟林木、藥用植物、農作物等植物中均分布著極為豐富的內生真菌,浸染了內生真菌的植物具有生長快速、抗逆性增加、抗病蟲能力增強等特性,比未感染內生真菌的植物更具生存的競爭力[3-5]。自從1993年首次發現源自短葉紅豆杉(Taxus brevifolia)韌皮部分的1株內生真菌(Taxomyces andreanae)能夠生產抗腫瘤活性成分紫杉醇以來,植物內生真菌的研究受到了極大的關注,并成為獲取活性天然產物的重要來源之一[6-7]。擬盤多毛孢屬(Pestalotiopsis)為半知菌門中一類無性型真菌,是植物內生真菌中最重要的類群之一[8]。Lee等[9]從粗榧(Cephalotaxus sinensis)中分離得到了內生小孢擬盤多毛孢(P.microspora),該內生菌產生的植物毒素對枯草芽孢桿菌(Bacillus subtilis)具有較強的抑制作用。迄今為止,從擬盤多毛孢屬內生真菌中發現結構新奇的化合物超過130多種[10-12],在醫藥、農業和林業領域方面具有潛在的應用價值[13-17]。
樟葉越桔(Vaccinium dunalianum)為杜鵑花科(Ericaceae)越桔屬常綠灌木[18],主要分布于云南、貴州、四川、西藏等地[19],具有清熱解毒、活絡散瘀等功效[20],據記載從明代起就利用其幼嫩葉芽作為茶代用品長期飲用至今。Zhao等[21]發現樟葉越桔富含咖啡酰熊果苷類物質,但其高含量蓄積是否與內生真菌協助增強宿主抗逆性有關尚不明確。越桔屬植物內生真菌多集中于菌根真菌的分離及其多樣性研究[22-24],而對樟葉越桔內生真菌的研究鮮有報道,僅樊苗苗等[25]發現樟葉越桔葉內生真菌對小麥雪腐病菌(Typhla incarnata)和三七絲核病菌(Verticillium cinnabarium)等的拮抗作用較強,陳肖學等[26]發現樟葉越桔葉內生真菌雜色曲霉(Aspergillus versicolor)的發酵液對棉花枯萎病菌(Fusarium oxysporium sp.vasinfectum)等5種農作物病原真菌具有較強的抑制作用。本研究采用形態鑒別和分子生物學手段對樟葉越桔嫩枝分離得到的1株內生真菌Z-78進行了鑒定,采用平板對峙法評價了其對綿腐臥孔菌(Poria placenta)等3種木腐菌和禾谷鐮孢(Fusarium graminearum)等5種植物病原真菌的抑制活性,并進行了Z-78發酵液抑制蕓苔鏈格孢的酸堿穩定性試驗,以期為該內生真菌進一步的開發利用提供參考。
1 材料與方法
1.1 材料
1.1.1 供試菌株 供試內生真菌Z-78從云南省武定縣采集的樟葉越桔健康嫩枝中分離純化得到。供試病原真菌菌株灰葡萄孢(Botrytis cinerea)CGMCC3.3790、腐皮鐮孢(Fusarium solani)CGMCC3.2889、禾谷鐮孢(F.graminearum)CGMCC3.4733、尖孢鐮孢(F.oxysporum)CGMCC3.3633和蕓苔鏈格孢(Alternaria brassicicola)CGMCC3.7805購自中科院微生物研究所菌種保藏中心,密粘褶菌(Gloeophyllum trabeum)CFCC86617、綿腐臥孔菌(Poria placenta)CFCC5608和采絨革蓋菌(Coriolus versicolor)CFCC5336購自中國林業微生物保藏管理中心,均保藏于西南林業大學西南山地森林資源保育與利用教育部重點實驗室。
1.1.2 培養基PDA培養基用于供試Z-78菌株和病原真菌培養,PDB培養基用于Z-78菌株發酵。
1.2 方法
1.2.1 內生真菌Z-78的鑒定 在超凈工作臺上將菌株Z-78接種到PDA培養基上,于28℃的恒溫培養箱中培養,待菌落長滿后觀察菌落的形態、菌絲、顏色、大小等,在光學顯微鏡下觀察菌絲及孢子形態。參照陳肖學等[26]的方法,采用Omegabiotek高純真菌DNA提取試劑盒提取菌株Z-78的DNA,采用真菌通用引物ITS1(5'-AGAAGTCGTAACAAGGTTTCCG TAGG-3')和ITS4(5'-TCCTCCGCTTATTGATATGC-3')進行PCR擴增。擴增體系:DNA模板4.0 μL,ddH2O 6.5 μL,ITS1 1.0 μL,ITS4 1.0 μL,2×Taq PCR maker Mix 12.5 μL。循環體系:第1階段1循環,94℃ 3 min;第2階段35循環,94℃ 30 s,51.5℃ 30 s,70℃ 1 min;第3階段1循環,70℃ 10 min。回收產物經電泳檢測后送上海生物工程有限公司進行序列測定,測序結果與GenBank上登錄的已知序列在Clustal X1.83軟件上進行同源性比對后,采用PAUP 4.0軟件構建系統發育樹。
1.2.2 菌株培養 在超凈工作臺中將已滅菌(121℃、22 min)的PDA培養基倒平板,每平板約20 mL的培養基,待冷卻凝固后用接種針挑取冷凍管里面保存的菌絲體接種于平板正中心位置,于28℃恒溫培養箱中培養7 d。用5 mm打孔器在培養7 d后的菌落邊緣打孔,取帶有菌絲的菌餅接種到新的PDA培養基上培養。發酵培養時,培養7 d的Z-78菌株用5 mm的打孔器在菌落的邊緣打孔切取直徑為5 mm的圓形菌餅,接種于體積為100.0 mL的液體培養基中,在恒溫振蕩器中振蕩培養7-10 d,培養條件28℃、180 r/min。用121℃滅菌后的紗布進行過濾,發酵液用0.22 μm無菌注射器過濾器進行過濾,置于4℃條件下保存。
1.2.3 抑菌活性測試 采用五點對峙培養法[25,27],測定Z-78菌株對供試病原菌的拮抗作用。用二代菌絲進行抑菌實驗,每個帶有PDA培養基的培養皿中分別放入5個菌餅,培養皿中間接入植物病原真菌,以病原菌為中心十字交叉距病原真菌2.5 cm處的4個點分別接入植物內生真菌,以在培養基中心接入植物病原真菌作為空白對照組,每個處理3個平行,恒溫培養,定時觀察,從對峙培養的第3天起用十字交叉法測量抑菌帶距離和菌落的直徑,直到空白長滿時停止測量。按公式計算抑菌率:抑菌率=(對照菌落直徑-處理菌落直徑)/對照菌落直徑×100%。參照劉澤星等[28]的方法,將抑菌率<25%標記為“+”,25%≤抑菌率<50%標記為“++”,抑菌率≥50%標記為“+++”,“-”表示沒有抑制作用。
1.2.4 內生真菌Z-78發酵液的酸堿穩定性測定 參考陳肖學等[26]和張艷軍[29]的方法,對菌株Z-78進行液體發酵,其發酵液分別用NaOH和HCl調至pH為2.0、4.0、6.0、8.0、10.0,靜置24 h之后,采用生長速率法[26,30],取4.0 mL處理后的菌株發酵液加到6.0 mL PDA培養基中,以不加發酵液的平板為空白對照,以未做處理發酵液的平板為條件對照,以對環境適應能力較強的蕓苔鏈格孢[31]作為指示菌,28℃條件下培養,分別從培養第2、4、6、8、10天后測量數據,按公式計算抑菌率。抑菌率=(對照組供試菌凈生長半徑-處理組供試菌凈生長半徑)/對照組供試菌凈生長半徑×100%。
2 結果
2.1 內生真菌Z-78的鑒定
觀察菌株Z-78的形態與檸檬擬盤多毛孢(P.citrina)相似[32],菌落呈輪紋狀均勻的向外擴長,開始生長是白色,一段時間后變為淡黃色,背面呈暗黃色。分生孢子團細小黑點狀,散生在菌絲表面,為梭形5細胞,稍彎曲,(16.5-23.1)×(5.6-7.3)μm;中間3個色胞幾乎同色,第3色胞淺褐色,上2色胞褐色,分隔處稍溢縮,長6.5-14.5 μm;頂胞無色,梯形,頂端有附屬絲2-3根,長10.0-20.0 μm,菌落、菌絲及孢子結構見圖1。參考《中國真菌志》[33],上述形態特征與木防己擬盤多毛孢菌(Pestalotiopsis cocculi)基本一致[34]。
在NCBI中采用BLAST對Z-78的ITS序列進行比對,用于同源性分析的序列分別是:木防已擬盤多毛孢(P.cocculi)KY703411、彌澤那擬盤多毛孢(P.menezesiana)JN198523[35]、AY687302、芒果擬盤多毛孢(P.mangiferae)KU194426、KX778664、聚多擬盤多毛孢(P.sydowiana)FJ478105、JQ322999、P.diploclisia KM199320[36]、KM199315、杭州擬盤多毛孢(P.hangzhouensis)AY526871、AY526870、小孢擬盤多毛孢(P.microspora)KT372347、KX755256和棒狀擬盤多毛孢(P.clavata)KP050596、KC537803,外源序列選擇不同科不同屬的白座蕉孢殼(Diatrype stigma)KT004563[37]。基于rDNA-ITS構建的系統發育樹如圖2所示,Z-78與木防已擬盤多毛孢聚為一支,結合上述形態特征及分子鑒定結果,菌株Z-78鑒定為擬盤多毛孢屬木防已擬盤多毛孢[Pestalotiopsis cocculi Vuill(Guba)G.C.Zhao &N.Li]。

圖1 菌株Z-78的正面(a)、背面(b)生長狀況、菌絲(c)和產孢結構(d)
2.2 內生真菌Z-78對病原菌的拮抗作用
由表1可知,內生真菌Z-78對8種不同供試病原菌呈現出不同程度的抑菌活性,抑菌率在50%-72.22%之間,表明菌株Z-78對供試病原菌具有較強拮抗能力,其拮抗效果見圖3所示。在3種木腐菌中,Z-78對密粘褶菌的抑菌活性最強(圖3-A),對綿腐臥孔菌(圖3-B)次之,對采絨革蓋菌(圖3-C)相對較弱,抑菌率分別為72.22%、70.39%和57.84%。Z-78對5種常見農作物病原真菌的抑菌活性差異不大,按強弱順序依次為尖孢鐮孢(圖3-D)>禾谷鐮孢(圖3-E)>蕓苔鏈格孢(圖3-F)>腐皮鐮孢(圖3-G)>灰葡萄孢(圖3-H),抑菌率分別為61.76%、61.11%、61.00%、52.35%和50.00%。
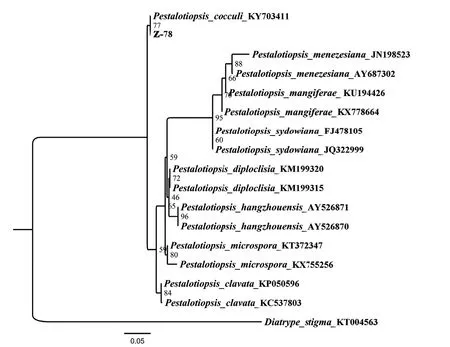
圖2 基于內生真菌Z-78 rDNA-ITS序列的系統發育分析
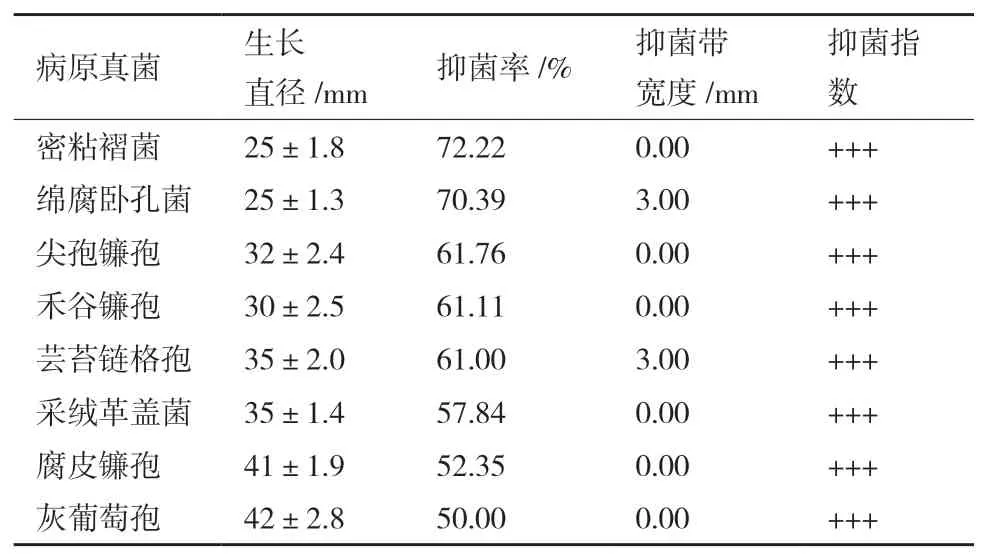
表1 內生真菌Z-78對8種植物病原菌的抑菌率
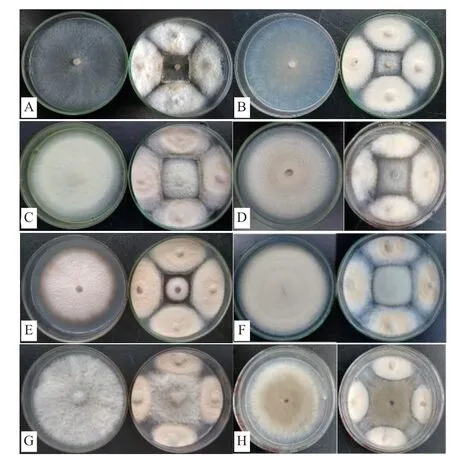
圖3 內生真菌Z-78對8種植物病原菌的拮抗效果
從表1中可以看出,多數對峙結果中的抑菌帶寬度均為0 mm。例如,在菌株Z-78與腐皮鐮孢的對峙培養中(圖3-G),兩者菌落之間沒有產生抑菌帶,但菌株Z-78菌落隨著生長時間增加,逐漸覆蓋腐皮鐮孢菌落,導致腐皮鐮孢的生長受到了抑制。而有些病原菌株的菌落生長速度緩慢(圖3-E),菌株Z-78可能釋放一些代謝產物到培養基中,或產生一些揮發性抑菌氣體,使得禾谷鐮孢生長受制。并形成較寬的抑菌帶。從圖4可以看出菌株Z-78對8種植物病原菌的拮抗作用隨著時間的推移逐漸增強,當抑制率達到頂峰時趨于一個相對平穩的狀態,說明抑菌效果相對穩定。同時,也發現菌株Z-78對8種植物病原菌在不同培養時間也呈現出不同程度的抑菌活性。
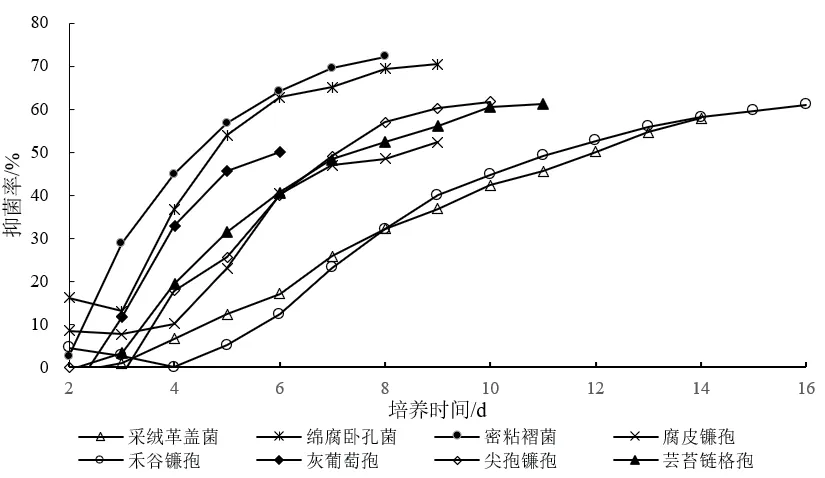
圖4 內生真菌Z-78對8種植物病原真菌的拮抗動態變化
2.3 內生真菌Z-78發酵液的酸堿穩定性試驗結果
內生真菌Z-78不同pH發酵液在不同培養時間下對蕓苔鏈格孢的抑制作用,如表2所示。從表2可知,內生真菌Z-78不同pH發酵液在不同培養時間下對蕓苔鏈格孢的抑制率均有不同程度的變化但差異不大,且變化趨勢一致,即抑菌率從開始逐漸增大,到第8天達到最大值后逐漸減小。Z-78發酵液的初始pH值是5.0-6.0,當pH 6.0時,其對蕓苔鏈格孢的抑菌率為55%,五點對峙培養法所得的抑菌率為61%(表1),說明兩種方法所得的抑菌率較為接近。當發酵液pH達到10.0時,第8天的抑制率達到最大值為60%,pH 6.0以下時的最大抑制率為57%,表明其抑制作用波動不大。從培養第10天的抑菌率來看,除pH 8.0外其他pH發酵液的抑菌率均大于50%,顯示出Z-78發酵液對蕓苔鏈格孢具有較強的抑制作用。在添加了4.0 mL pH值為2.0、4.0、6.0、8.0、10.0的無菌蒸餾水的平板對照組中,病原真菌的菌落生長未受到明顯的影響,表明Z-78發酵液對蕓苔鏈格孢的抗菌活性較為穩定,不會受到酸堿性的影響。
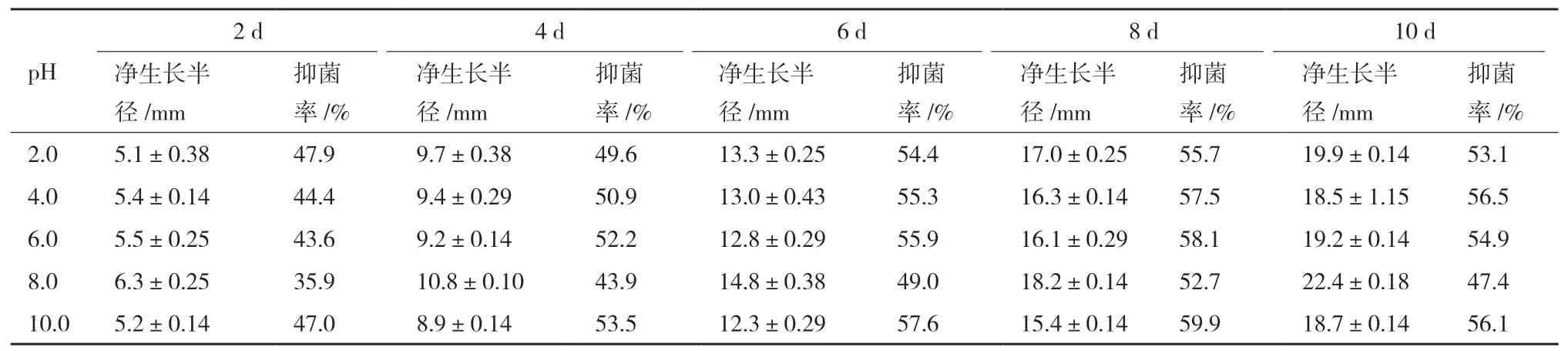
表2 不同pH條件下Z-78發酵液對蕓苔鏈格孢的抑制作用
3 討論
研究結果表明,大約有30%的內生真菌能夠生產抗菌活性化合物,可對多種植物病原真菌具有專一拮抗或廣譜抗性[38],其拮抗機制通常包括分泌抑菌物質、產生抑菌揮發性氣體、競爭作用、重寄生作用等[39]。擬盤多毛孢是植物內生真菌的重要類群,次生代謝產物種類豐富,具有抗癌、抗菌等生理活性,具有良好的應用開發前景[8,13]。易曉華等[40]報道源自除蟲菊(Pyrethryum cinerariifolium)葉擬盤多毛孢屬內生真菌Y1菌株對蘋果炭疽病菌(Glomerella cingulata)、灰葡萄孢、玉米大斑病菌(Exserohilum turcicum)等6種植物病原真菌的抑制作用達到85%以上。李靖等[41]發現石楠銹孢銹菌(Aecidium pourthiaea)重寄生擬盤多毛孢(Pestalotiopsis sp.)對辣椒炭疽病菌(Sclerotinia sclerotiorum)等10種常見植物病原真菌具有一定的抑制作用,李向楠等[42]也發現源自云南熱區的多種該屬內生真菌顯示出廣譜抗菌活性,表明不同來源擬盤多毛孢屬內生真菌具有較為顯著的抑菌活性。Li等[43]報道源自歐洲紅豆杉(Taxus baccata)、落羽杉(Taxodium distichum)、佛羅里達榧樹(Torreya taxifolia)和石斛(Dendrobium sp.)的內生小孢擬盤多毛孢(P.microspora),以及從澳大利亞瓦勒邁杉(Wollemia nobilis)分離的斑污擬盤多毛孢(P.maculans)的液體發酵液含有的環己烷類物質,對腐皮鐮孢、古巴鐮孢(Fusarium cubense)、麥根腐長孺孢(Helminthosporium sativum)、蒂腐色二孢(Diplodia natelensis)等病原真菌具有抗菌作用。源自紅樹林海蓮(Bruguiera sexangula)枝的污斑擬盤多毛孢(P.foedan)中的單萜內酯類對灰葡萄孢和煙草疫霉菌(Phytophthora nicotianae)[44],源自鐵皮石斛幼苗的該屬DO14菌株中的吡喃酮類物質對白色念珠菌(Candida albicans)、新型隱球菌(Cryptococcus neoformans)、紅色毛蘚菌(Trichophyton rubrum)和煙曲霉(Aspergillus fumigatus)[45],源自茶(Camellia sinensis)枝的無花果擬盤多毛孢(P.fici)中的吡喃酮類物質對禾谷鐮孢[46],源自芒果(Mangifera indica)的芒果擬盤多毛孢(P.mangiferae)中的酚類物質對白色念珠菌[47]顯示有較強的抗菌活性,表明不同來源該屬內生真菌的抑菌活性物質基礎及其抑菌機制值得進一步深入研究。
4 結論
本研究對樟葉越桔嫩枝1株內生真菌Z-78進行了形態鑒別和分子鑒定,將其確定為木防已擬盤多毛孢[Pestalotiopsis cocculi(Guba)G.C.Zhao & N.Li]。采用五點對峙法評價其對密粘褶菌等3種木腐菌、尖孢鐮孢等5種常見植物病原真菌的拮抗活性,結果表明菌株Z-78對8種供試病原菌的生長均具有較強的抑制作用,其中對密粘褶菌的抑菌活性最強,并發現Z-78發酵液在酸堿性條件下對蕓苔鏈格孢的抑菌活性較為穩定,研究結果將為樟葉越桔枝內生真菌木防已擬盤多毛孢中抑菌活性物質進一步的分離鑒定和開發利用提供了參考。
