橡膠樹HbCPA基因的克隆、表達及生物信息學分析
2020-12-04 08:06:58楊洪胡燕玲岳鎰繁鄧治代龍軍李德軍
生物技術通報 2020年11期
楊洪 胡燕玲,2 岳鎰繁,2 鄧治 代龍軍 李德軍
(1.中國熱帶農業科學院橡膠研究所 農業農村部橡膠樹生物學與遺傳資源利用重點實驗室 省部共建國家重點實驗室培育基地-海南省熱帶作物栽培生理學重點實驗室,海口 571101;2.南京農業大學農學院,南京 210095)
橡膠是四大工業原料之一,根據來源不同分為天然橡膠和合成橡膠。天然橡膠在彈性、耐磨性和延展性等特性方面具有合成橡膠不可替代的優勢,在一些重要工業領域(如航空、航天和重型汽車等制造業)廣泛應用。我國是天然橡膠第一大消費國和進口國,自2017年我國天然橡膠年消費量和進口量均在5.0×106t以上,而我國天然橡膠年產量持續在8.0×105t上下波動,供需缺口懸殊,需要通過進口來填補國內天然橡膠資源短缺。巴西橡膠樹(Hevea brasiliensis,簡稱橡膠樹)是典型的熱帶雨林喬木,是天然橡膠的主要來源[1]。天然橡膠產業是典型的環境約束型產業,受自然因素約束我國適宜種植橡膠樹的地區主要集中在海南、廣東、云南三地。截至2018年末,我國天然橡膠種植面積約為1.15×106hm2。目前,我國天然橡膠產業面臨植膠區域有限且種植趨于飽和及天然橡膠價格持續低迷等問題,已很難通過擴大種植面積來增加產量。我國地處熱帶北緣,橡膠樹在生長周期中不可避免地會遭受低溫寒害、臺風和季節性干旱等氣候影響。不良的氣候環境嚴重影響橡膠樹單位面積產量,縮短橡膠樹經濟壽命。因此,提高橡膠樹單位面積產量及其對逆境脅迫的耐受能力是緩解天然橡膠供需平衡壓力的最佳途徑。
多胺(Polyamine,PAs)是一類小分子脂肪族含氮多聚物,幾乎存在于所有的生物體。在植物中多胺主要以腐胺(Putrescine,Put)、亞精胺(Spermidine,Spd)和精胺(Spermine,Spm)的形式存在,其中Put是多胺生物合成的中心產物[2-4]。Put的合成涉及兩條途徑,一條為通過鳥氨酸脫羧酶(Ornithine decarboxylases,ODC)催化鳥氨酸脫羧形成腐胺的簡單反應,稱為鳥氨酸途徑;另一條以精氨酸為起始物質,因此稱為精氨酸途徑。精氨酸經精氨酸脫羧酶(Arginine decarboxylase,ADC)脫羧形成鯡精胺(Agmatine,Agm),鯡精胺在鯡精胺亞氨水解酶(Agmatine iminohydrolase/Agmatine deiminase,AIH)的催化下水解生成N-氨甲基腐胺(N-carbamoylputrescine,NCP),最后在N-氨甲酰基腐胺酰胺水解酶(N-carbamoylputreseine amidohydrolase,CPA)的催化水解下生成腐胺[5-7]。植物是唯一能夠通過精氨酸途徑合成Put的真核生物,該途徑是某些植物(如擬南芥、小立碗蘚)腐胺合成的唯一途徑[8]。
在農業生產中,常通過外源施加PAs來提高作物產量和增強作物耐逆性。例如,外源施加Put能夠提高小麥的耐熱性[9],減輕蘋果愈傷組織鹽脅迫損傷[10]。利用轉基因技術提高PAs合成相關基因的表達能使內源PAs在生物體內積累,從而達到作物抗逆、增產的目的[11-15]。Put是橡膠樹膠乳中的主要PAs,割膠能夠促進其生物合成[16]。目前,關于橡膠樹PAs合成相關基因的研究僅限于SAMDC、ADC、AIH等少數幾個基因的克隆及功能分析[17-19]。為進一步豐富和了解橡膠樹PAs合成途徑相關基因的生物學功能及表達特性,本文從橡膠樹品系熱研7-33-97中克隆了首個CPA基因并對其進行生物信息學及表達特性分析。
1 材料與方法
1.1 材料
本文所用材料均為橡膠樹品系熱研7-33-97。健康樹樹皮、葉片、莖尖、雌花、雄花、膠乳等組織樣品及死皮橡膠樹膠乳、樹皮樣品均采自中國熱帶農業科學院試驗農場;不同發育階段葉片樣品采自國家橡膠樹種質資源圃(儋州);過氧化氫(H2O2)、乙烯利(Ethephon,ET)和茉莉酸甲酯(Methyl jasmonate,MeJA)處理的膠乳樣品均來自于已達到開割標準的未開割橡膠樹;低溫、干旱、高鹽脅迫處理的葉片樣品取自移栽6個月且長勢基本一致的橡膠樹組培苗,處理方法參照楊洪等[19]方法。
1.2 方法
1.2.1 總RNA提取及cDNA第一鏈合成 用北京百泰克生物技術有限公司生產的通用植物總RNA提取試劑盒,按其使用說明書提取樣品總RNA,分光光度法檢測RNA濃度,瓊脂糖凝膠電泳進行完整性檢測。參照PrimeScriptTMRT reagent Kit with gDNA Eraser試劑盒說明書進行樣品cDNA合成。
1.2.2 HbCPA全長序列克隆 進行橡膠樹轉錄組測序結果分析時我們獲得了一條與植物CPA基因序列高度同源的unigene,通過NCBI的blast比對確定其為CPA基因。由于該unigene缺少閱讀框3'末端序列,因此采用SMARTerTMRACE cDNA Amplification Kit(Clontech,USA)試劑盒進行3'-RACE擴增。3'-RACE所用引物為5'-CACATTTTATGGGAACTCATTC-3'和5'-CAAATCCAAGAGACATAGTTGG-3'。按照試劑盒說明書用3'-RACE CDS Primer A對1 μg各組織混合RNA進行反轉錄,獲得用于3'-RACE的模板cDNA。以獲得的3'-RACE cDNA為模板用PrimeSTAR Max Premix(TaKaRa,China)進行3'端序列擴增。將獲得的3'端序列與轉錄組中獲得的unigene序列進行拼接,根據拼接序列并設計全長cDNA擴增引物,進行PCR和測序驗證。
1.2.3 HbCPA生物信息學及進化分析 用NCBI的ORF finder預測開放閱讀框;用NCBI保守結構域數據庫網站(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)預測橡膠樹CPA蛋白保守結構域;用SMART(http://smart.embl-heidelberg.de/)分析蛋白保守結構域;用Expasy網站的Prot-Param程序(http://web.expasy.org/protparam/)分析蛋白質理化性質;用NCBI中的BLAST和DNASTAR軟件進行同源性分析;用SignalP 4.1Server 軟件(http://www.cbs.dtu.dk/services/SignalP/)進行信號肽預測分析;用TMHMM 2.0程序(http://www.cbs.dtu.dk/services/TMHMM/)預測跨膜結構域;利用在線工具(https://wolfpsort.hgc.jp)分析目的序列的亞細胞定位;用ClustalX和MEGA6.0進行多序列比對和Neighbor-Joining進化樹的構建,用多序列比對顯色工具ESPript對多序列比對顯示美化[20]。
1.2.4 實時熒光定量PCR 根據CPA序列設計特異性熒光定量引物(F:5'-GACGTCCAGATTTGTACAAGG-3'和R:5'-CCACAAAAAATTCATGGTTGGG-3')。以橡膠樹18S rRNA基因為內參基因(F:5'-GCTCGAAGACGATCAGATACC-3'和R:5'-TTCAGCCTTGCGACCATAC-3')。以不同樣品反轉錄cDNA稀釋5倍后為模板,反應體系為20 μL,包含2 μL模板,10 μL 2×SYBR Premix、10 μmol/L上游引物和下游引物各1 μL、滅菌水補足至20 μL。qPCR在Bio-Rad CFX96 qPCR儀進行,程序設置為:95℃預變性5 min;95℃ 10 s,58℃ 30 s,72℃ 20 s,40個循環后進行熔解曲線分析,以確定引物的特異性。基因相對表達量采用公式2-ΔΔCt計算。
2 結果
2.1 HbCPA的克隆及序列分析
通過對橡膠樹轉錄組測序結果分析,獲得了一條長度為1 068 bp的unigene。NCBI在線blast比對顯示該unigene與高度植物CPA基因序列相似性均在85%以上,推測其為橡膠樹CPA基因。由于該unigene缺少閱讀框3'末端序列,因此進行了3'-RACE擴增。經過測序、拼接及RT-PCR驗證,獲得具有完整閱讀框的橡膠樹CPA基因全長序列,并將其命名為HbCPA。HbCPA序列全長1 342 bp,開放閱讀框906 bp,編碼301個氨基酸(圖1)。
HbCPA預測理論分子量約為33.48 kD,理論等電點為6.01。不穩定系數為33.67,為穩定蛋白。信號肽預測結果表明,HbCPA不含信號肽,為胞內蛋白。疏水性分析表明,該蛋白親水性氨基酸殘基為62個,占20.60%;疏水性氨基酸殘基為113個,占37.54%,平均疏水系數為-0.269,為親水性蛋白。氨基酸組成中帶負電荷的氨基酸(Asp+Glu)共有40個,占13.29%;帶正電荷的氨基酸(Arg+Lys)共有35個,占11.63%。跨膜結構域預測結果表明,HbCPA不含跨膜結構域,為非膜蛋白。亞細胞定位預測結果表明,HbCPA定位于細胞質。
2.2 HbCPA同源比對及二級結構特征
氨基酸序列比對結果表明CPAs在不同植物中高度相似,本研究所選物種的CPAs與HbCPA的一致性均大于87%。HbCPA與木薯MeCPA氨基酸序列一致性最高,達98.01%;其次是克萊門氏小柑橘CcCPA和大戟科的麻風樹JcCPA,一致性均為92.36%。二級結構預測結果表明無規則卷曲為HbCPA二級結構的主要構成元件,占39.87%;其次是α-螺旋和β-片層結構,占比分別為29.90%和23.36%;β-轉角結構最少,僅占6.98%(圖2)。
為了分析HbCPA與其他植物CPA蛋白的親緣關系,選取與HbCPA同源的15個不同植物CPAs進行系統進化分析。結果(圖3)表明,橡膠樹HbCPA先與同屬大戟科的木薯MeCPA、蓖麻RcCPA和麻風樹JcCPA聚為一枝,然后再與胡楊PeCPA和毛白楊PtCPA聚為一枝,表明它們有較近
2.3 HbCPA系統進化分析
的親緣關系。而蒺藜苜蓿MtCPA、大豆GmCPA和水稻OsCPA聚為較遠的一枝,暗示它們與HbCPA的親緣關系較遠。

圖1 HbCPA cDNA序列及其編碼的氨基酸序列
2.4 HbCPA組織表達特性及對橡膠樹死皮的響應
實時熒光定量PCR結果(圖4-A)顯示,HbCPA在橡膠樹的不同組織(樹皮、葉片、莖尖、雌花和雄花)均有較高的表達量,但其表達存在組織差異。表達量從高到低依次最高為樹皮、葉片、雌花、雄花、莖尖。葉片不同發育時期HbCPA表達結果(圖4-B)表明,淡綠期HbCPA的表達量遠高于其他4個時期,變色期和穩定期表達量相當,古銅期和衰老期表達量相當,但變色期和穩定期的表達量高于古銅期和衰老期。比較健康和死皮橡膠樹中表達差異發現(圖4-C),HbCPA在死皮橡膠樹樹皮中表達明顯上升,而在膠乳中HbCPA的表達基本一致。
2.5 HbCPA在逆境脅迫及激素處理的表達模式
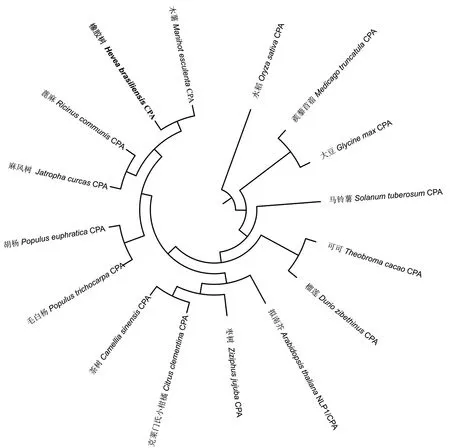
圖3 HbCPA與其它物種CPAs蛋白系統進化分析
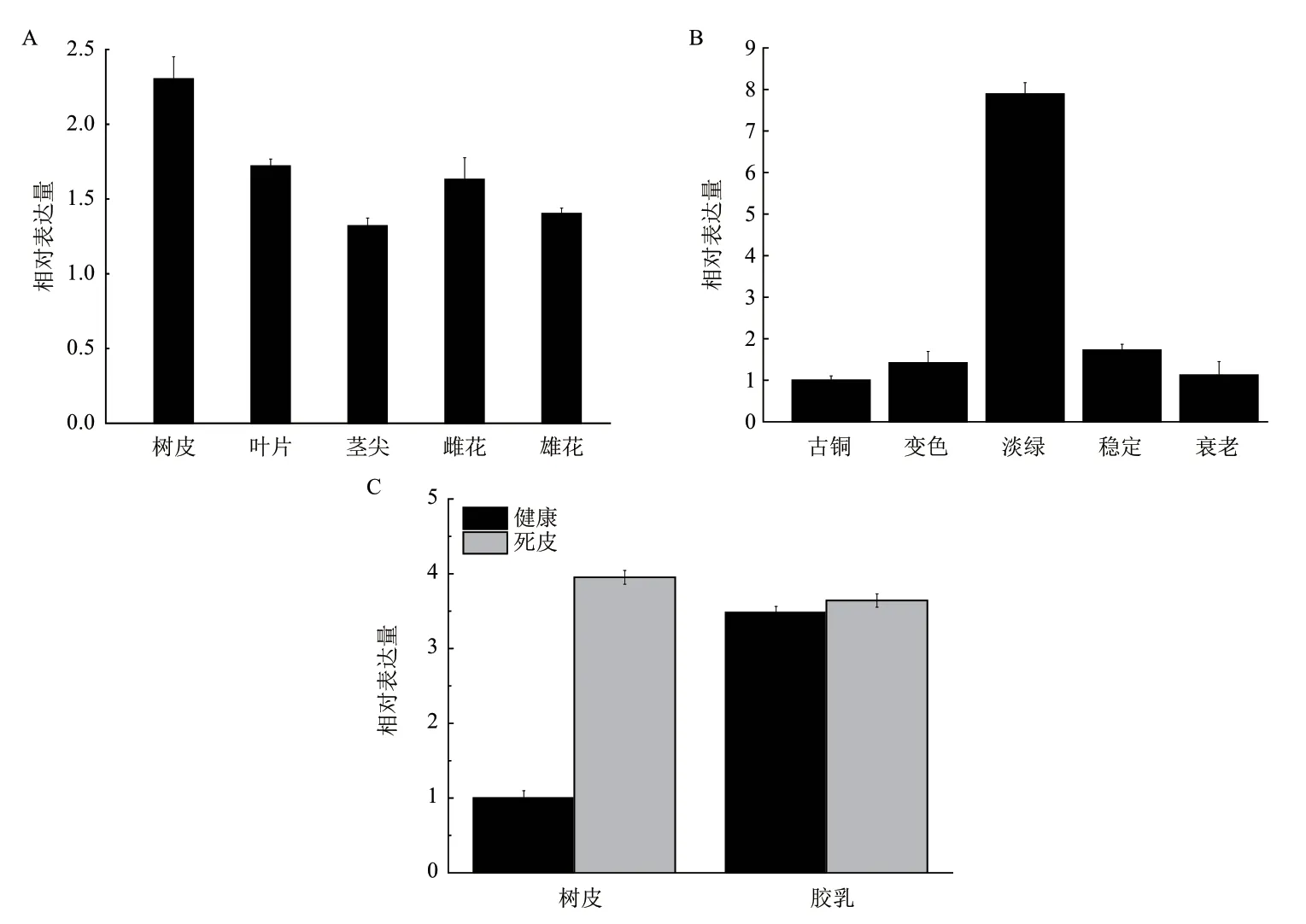
圖4 HbCPA在橡膠樹不同組織(A)、葉片不同發育時期(B)及健康/死皮橡膠樹膠乳和樹皮(C)表達模式
橡膠樹膠乳中HbCPA的表達還受乙烯利(ET)、茉莉酸甲酯(MeJA)等處理和低溫、干旱、高鹽和H2O2等逆境條件的影響。H2O2處理后HbCPA表達先下降再上升,處理24 h表達量最低,至48 h表達量略有上升但也低于處理前水平(圖5-A)。ET處理后膠乳中HbCPA表達呈現波動變化,處理后各時間點表達量均低于處理前水平。ET處理4 h表達量急劇下降至8 h表達量達到最低水平,24 h開始表達量開始上調,72 h開始下降至24 h的表達水平(圖5-B)。MeJA處理4 h HbCPA表達急劇下降,處理8 h后逐漸升高直至處理48 h,然后又開始下降至處理72 h其表達水平與24 h基本一致(圖5-C)。低溫處理抑制HbCPA的表達,并且隨著處理時間延長抑制作用越明顯(圖5-D)。膠乳中HbCPA的表達還受干旱和高鹽脅迫調控,且表達趨勢基本一致(圖5-E,F)。干旱或高鹽處理24 h,HbCPA表達達到最高水平,分別為處理前的3.3倍和4.4倍;處理48 h表達水平明顯下降,但仍略高于處理前水平(干旱)或與處理前一致(高鹽)。
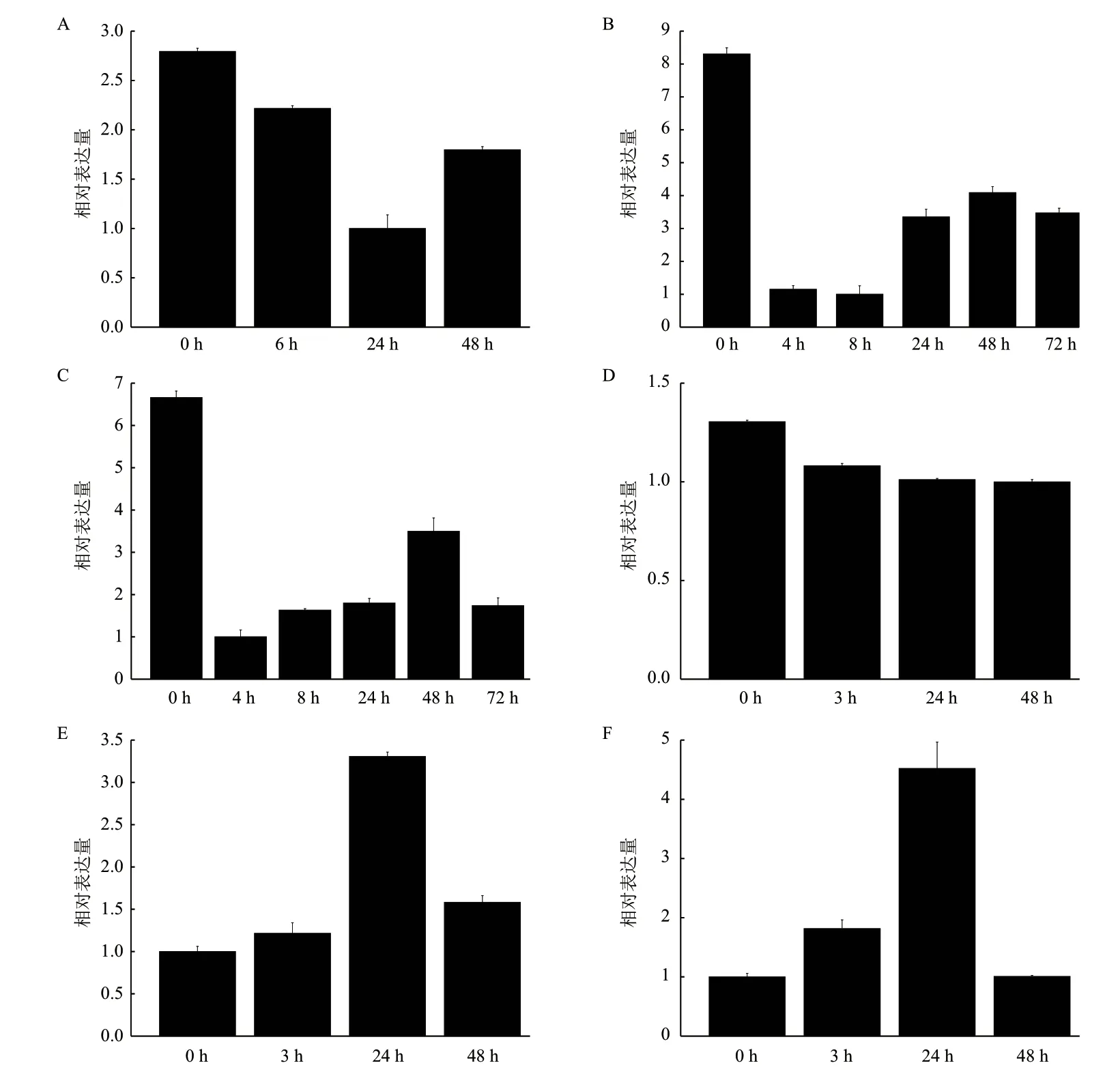
圖5 不同處理條件下HbCPA的表達模式
3 討論
CPA是一種腈水解酶,其功能活性依賴于Glu-Lys-Cys催化三聯體結構[21]。作為精氨酸途徑合成腐胺過程的終反應酶,深入了解CPA基因的結構、功能對理解生物體內PAs代謝平衡具有重要意義。本文首次從橡膠樹中克隆了一個CPA基因HbCPA,該基因ORF長906 bp,編碼301個氨基酸,含有其功能活性所需的Glu-Lys-Cys催化三聯體結構。生物信息學分析表明,HbCPA為親水性蛋白,不含跨膜結構域和信號肽,是一個胞質酶。序列分析表明HbCPA與其他植物的CPAs一致性很高,說明CPAs在不同植物中具有相似的結構和功能。進化分析表明HbCPA與同屬大戟科的木薯MeCPA、蓖麻RcCPA和麻風樹JcCPA親緣關系最近,與植物學傳統分類一致。
天然橡膠是在樹皮特化細胞——乳管中合成。組織表達譜分析表明,HbCPA在樹皮中的表達量最高,同時該基因在死皮樹樹皮中明顯上調表達。死皮是橡膠樹割面部分或全部不排膠的現象,是橡膠樹產膠、排膠障礙的極端形式。HbCPA的表達分析表明,該基因可能與橡膠樹橡膠生物合成和代謝相關,可能是橡膠樹產膠排膠的負向調控因子。橡膠樹葉片發育分為古銅期、變色期、淡綠期、穩定期和衰老期五個時期。質體的發生、發育是橡膠樹葉片由淡綠期轉化到穩定期的重要原因[22]。橡膠樹葉片不同發育時期的表達分析表明,HbCPA在葉片淡綠期大量表達,推測其可能與葉片由淡綠期轉化到穩定期密切相關,可能與質體的發生、發育有關。
乙烯作為延長排膠時間提高產膠量而被廣泛應用的植物激素,而多胺和乙烯的合成競爭同一底物S-腺苷蛋氨酸(SAM)[23]。MeJA是橡膠樹乳管分化和發育的重要調節因子,可以誘導橡膠樹乳管分化,調節橡膠生物合成[24-26]。本研究發現ET和MeJA處理均抑制了HbCPA的表達,但隨著處理時間的延長HbCPA表達整體呈上升趨勢,暗示HbCPA可能在調控橡膠樹產排膠信號轉導中發揮重要作用。CPAs還與植物逆境響應密切相關。水稻含有4個CPAs基因,分別為OsCPA1-4。干旱脅迫下OsCPA3表達下調而OsCPA1、OsCPA2、OsCPA4轉錄水平未出現變化[27]。擬南芥CPA在水脅迫處理下出現滯后上調表達[28];同樣的,在本研究中發現HbCPA表達也受干旱、低溫、高鹽和H2O2等逆境脅迫影響,表明HbCPA可能參與橡膠樹逆境脅迫應答。
4 結論
從橡膠樹中克隆獲得HbCPA,其開放閱讀框906 bp,編碼301個氨基酸,預測其為親水性蛋白。HbCPA表達無組織特異性,在檢測的組織中均有表達。HbCPA可能參與橡膠樹葉片由淡綠期向穩定期轉化過程,參與低溫、干旱、高鹽、氧化脅迫等非生物逆境應答等過程,并可能負向調控橡膠樹死皮發生。
