新型豬α干擾素YNS突變體的構(gòu)建及其抗病毒活性檢測(cè)
2020-12-09 08:36:36王朋濤梁秀麗徐盟龍趙福杰閆曉光魏戰(zhàn)勇
畜牧與獸醫(yī) 2020年12期
關(guān)鍵詞:檢測(cè)
王朋濤,梁秀麗,徐盟龍,趙福杰,閆曉光,魏戰(zhàn)勇, 3*
(1. 河南農(nóng)業(yè)大學(xué)牧醫(yī)工程學(xué)院,河南 鄭州 450002;2. 安陽(yáng)工學(xué)院,河南 安陽(yáng) 455000;3. 河南省動(dòng)物性食品安全重點(diǎn)實(shí)驗(yàn)室,河南 鄭州 450002)
干擾素(interferon, IFN)是一類具有抗病毒、抑制細(xì)胞增殖等多功能糖蛋白,在適應(yīng)性免疫調(diào)節(jié)中起著重要作用[1]。干擾素在病原入侵機(jī)體時(shí)誘導(dǎo)相關(guān)抗病毒蛋白的表達(dá),例如:蛋白激酶R、干擾素誘導(dǎo)黏病毒抵抗蛋白等[2-4],使機(jī)體處于抗病毒狀態(tài)。干擾素廣泛用于養(yǎng)殖業(yè),在藍(lán)耳病、口蹄病等病毒性疾病治療中效果顯著[5-6]。
豬α干擾素基因編碼166個(gè)氨基酸,其空間結(jié)構(gòu)主要由5個(gè)α螺旋組成,無(wú)內(nèi)含子,有至少17個(gè)亞型[7-8]。豬α干擾素在機(jī)體內(nèi)分布廣泛,但表達(dá)量較低,無(wú)法滿足生產(chǎn)需要[9]。因此許多學(xué)者利用基因工程對(duì)豬α干擾素進(jìn)行研究。Lefevre等[10]對(duì)豬α干擾素進(jìn)行體外克隆、基因序列分析和少量表達(dá)。陳濤等[11]改變豬α干擾素第86 位氨基酸, 并對(duì)其密碼子進(jìn)行優(yōu)化,獲得蛋白活性為52 000 IU/mg的突變體。李爽等[12]改變豬α干擾素不同的氨基酸活性位點(diǎn),獲得了蛋白活性為2.97× 108IU/mg的突變體。本研究將天然豬α干擾素的第58位組氨酸H、59位谷氨酸E及61位亮氨酸L分別突變?yōu)槔野彼醂、天冬氨酸N及絲氨酸S,成功構(gòu)建了豬α干擾素YNS突變體,運(yùn)用大腸桿菌系統(tǒng)表達(dá)純化后,其抗病毒活性與天然豬α干擾素相比明顯提高,為干擾素的臨床應(yīng)用提供理論基礎(chǔ)。
1 材料與方法
1.1 質(zhì)粒與菌株
含有豬α干擾素基因的重組質(zhì)粒、原核表達(dá)載體pET-32a、大腸桿菌感受態(tài)細(xì)胞DH-5α和Rosetta(DE3)由河南省動(dòng)物性食品安全重點(diǎn)實(shí)驗(yàn)室保存。
1.2 細(xì)胞與病毒
Vero細(xì)胞和水泡性口炎病毒(VSV-Indiana)由中科院武漢病毒研究所惠贈(zèng)。
1.3 主要試劑
限制性內(nèi)切酶、T4 DNA連接酶等購(gòu)自北京寶日醫(yī)生物技術(shù)有限公司,豬α干擾素單克隆抗體(G16,Santa Cruz)購(gòu)自北京百奧曼科技有限公司,羊抗鼠IgG(HRP標(biāo)記)購(gòu)自武漢三鷹生物技術(shù)有限公司,其他試劑均為國(guó)產(chǎn)分析純。
1.4 引物設(shè)計(jì)及合成
根據(jù)豬α干擾素基因(登陸號(hào):AB369102.1)設(shè)計(jì)兩對(duì)引物, 其中F1和R1用于擴(kuò)增完整的豬α干擾素基因并包含BamH Ⅰ和XhoⅠ酶切位點(diǎn),W1和W2反向互補(bǔ)且包含突變位點(diǎn)(表1)。引物由鄭州尚亞生物工程有限公司合成。
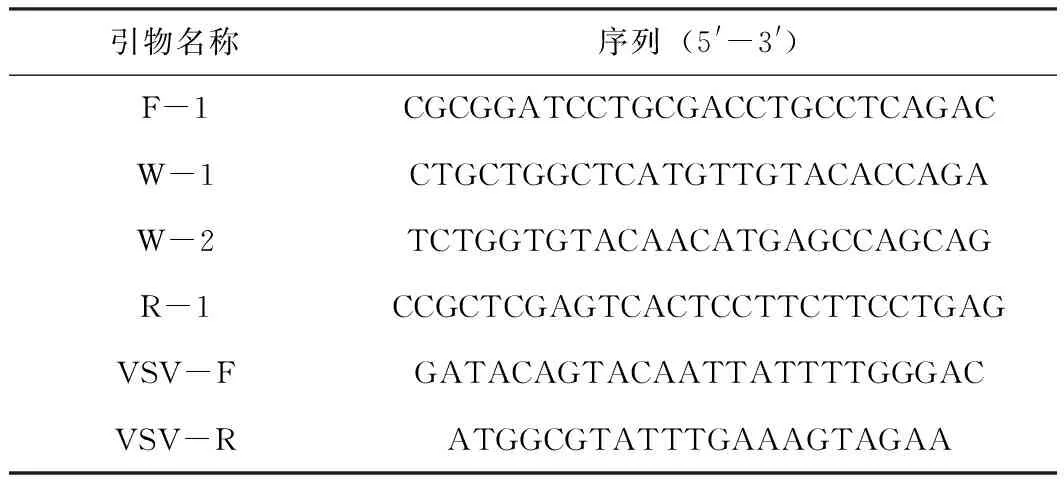
表1 引物序列
1.5 豬α干擾素YNS突變體基因的擴(kuò)增
以豬α干擾素基因重組質(zhì)粒為模板,利用F-1、W-1和W-2、R-1兩對(duì)引物進(jìn)行PCR擴(kuò)增,擴(kuò)增片段命名為P1和P2。PCR反應(yīng)程序?yàn)椋?5 ℃預(yù)變性5 min,95 ℃變性30 s,58 ℃退火30 s,72 ℃延伸20 s,35個(gè)循環(huán)。反應(yīng)結(jié)束后,以F-1、R-1為引物,以P1、P2為模板進(jìn)行擴(kuò)增,擴(kuò)增程序如上。反應(yīng)結(jié)束后用1%的瓊脂糖凝膠電泳檢測(cè)PCR產(chǎn)物。
1.6 重組表達(dá)載體pET-32a-IFN-α的構(gòu)建
用BamHⅠ、XhoⅠ分別對(duì)PCR產(chǎn)物和表達(dá)質(zhì)粒進(jìn)行雙酶切,16 ℃連接。轉(zhuǎn)入大腸桿菌DH5α,篩選陽(yáng)性克隆并送至武漢奧科生物有限公司進(jìn)行序列測(cè)定。
1.7 重組質(zhì)粒pET-32a-IFN-α的表達(dá)及鑒定
將鑒定正確的陽(yáng)性質(zhì)粒轉(zhuǎn)至表達(dá)菌Rosetta中,挑選陽(yáng)性克隆,37 ℃、200 r/min 振蕩擴(kuò)大培養(yǎng)至OD值為0.6~0.8,加入IPTG(1 mmol/L)誘導(dǎo)8 h。收集菌體進(jìn)行超聲波裂解,4℃、12 000 r/min離心,取沉淀和上清進(jìn)行SDS-PAGE檢測(cè)和Western blot鑒定。
1.8 表達(dá)產(chǎn)物的純化
經(jīng)超聲波破碎后,4 ℃、12 000 r/min離心,4 mol/L尿素(無(wú)菌PBS配制)洗滌沉淀3次以除去雜蛋白,4 ℃、12 000 r/min離心。取適量的6 mol/L尿素溶解沉淀,分別用5、4、3、2、1 mol/L尿素溶液依次透析,每個(gè)階段6 h,最后用無(wú)菌的PBS(pH 7.2)透析24 h。測(cè)定蛋白含量,-80 ℃保存。
1.9 蛋白內(nèi)毒素測(cè)定
利用ToxinSensorTM內(nèi)毒素檢測(cè)試劑盒(L00350)檢測(cè)純化后干擾素內(nèi)毒素的含量,具體步驟參照試劑盒說明書。
1.10 干擾素的生物活性測(cè)定
VSV的半數(shù)細(xì)胞感染量(TCID50)測(cè)定:將Vero細(xì)胞接種到96孔板中, 24 h后每孔加入10倍倍比稀釋的VSV病毒100 μL,每個(gè)稀釋度設(shè)6個(gè)重復(fù)孔。繼續(xù)培養(yǎng)72 h,按Reed-Muench法計(jì)算TCID50。
MTT法檢測(cè)干擾素在Vero細(xì)胞上的安全濃度:利用MTT試劑盒(索萊寶M1020-500T)檢測(cè),酶聯(lián)免疫檢測(cè)儀在OD490nm處測(cè)量生物活性。
抗病毒活性的測(cè)定:采用細(xì)胞病變(CPE)抑制法在Vero-VSV系統(tǒng)上測(cè)定豬α干擾素突變體蛋白的抗病毒活性。同時(shí)對(duì)VSV的TCID50和基因拷貝數(shù)進(jìn)行檢測(cè)。將Vero細(xì)胞接種到6孔板中,24 h后每孔加入濃度分別為100 ng/mL和1 ng/mL的干擾素100 μL,同時(shí)設(shè)置對(duì)照組。繼續(xù)培養(yǎng)24 h,試驗(yàn)組每孔接入100 TCID50/0.1 mL VSV。培養(yǎng)至病毒對(duì)照組有75%細(xì)胞出現(xiàn)病變時(shí),分別收取上清和細(xì)胞檢測(cè)。VSV(Real-time pCR)檢測(cè)方法及其標(biāo)準(zhǔn)曲線參照文獻(xiàn)[13]方法建立。引物序列如表1。
1.11 統(tǒng)計(jì)分析
采用Prism 6.0進(jìn)行的統(tǒng)計(jì)分析,每個(gè)數(shù)據(jù)重復(fù)3次,試驗(yàn)數(shù)據(jù)是用平均值顯示,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結(jié)果
2.1 豬α干擾素突變體的克隆
豬α干擾素突變體基因擴(kuò)增產(chǎn)物,核酸電泳和測(cè)序顯示,擴(kuò)增片段大小符合預(yù)期,長(zhǎng) 501 bp并含有3個(gè)預(yù)期突變位點(diǎn),與參照序列同源性97.6%。雙酶切鑒定結(jié)果顯示成功構(gòu)建豬α干擾素突變體表達(dá)質(zhì)粒(如圖2)。
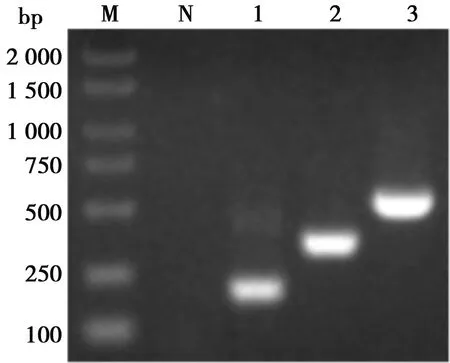
M. DL 2000 DNA Marker;1. 片段P1; 2. 片段P2; 3. 豬α干擾素;N. 陰性對(duì)照
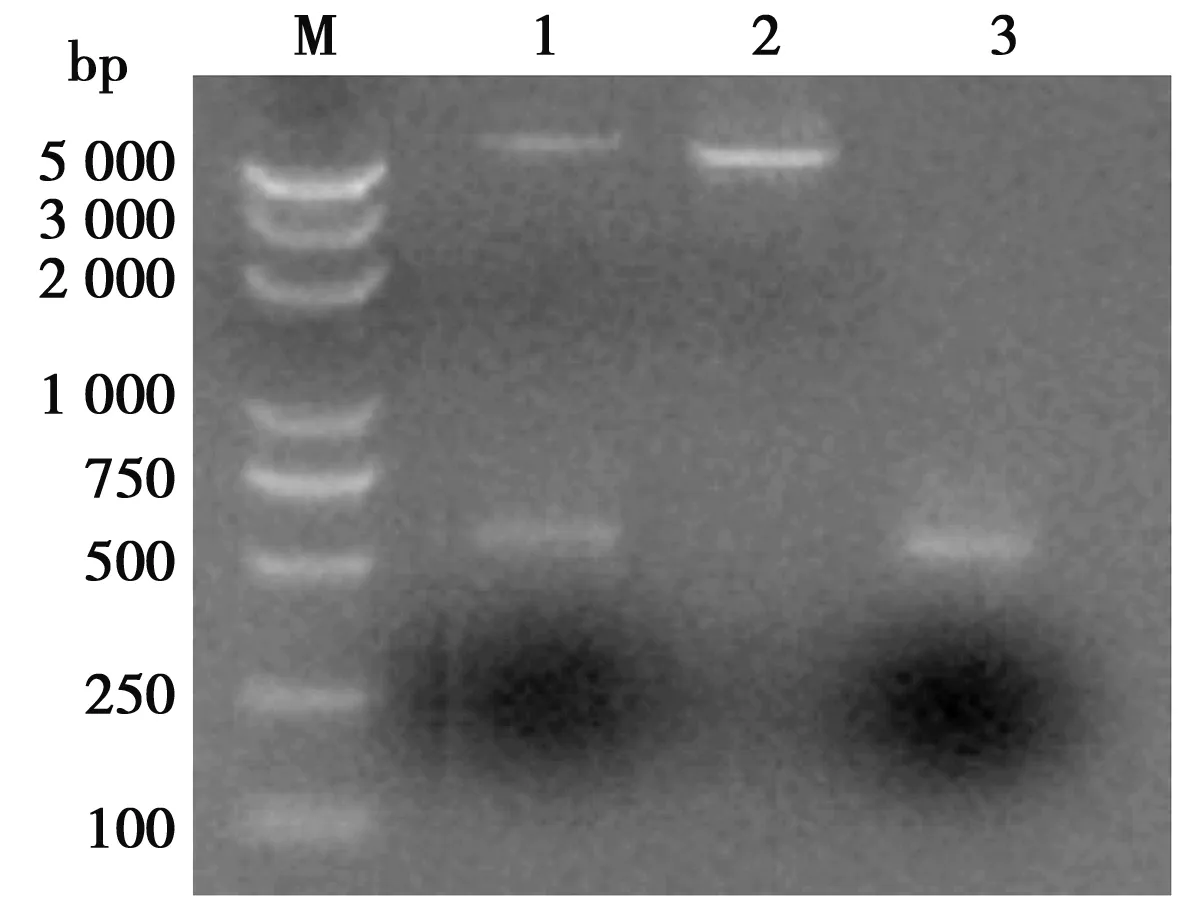
M. DL 5000 DNA Marker;1. pET-32a--IFN-α雙酶切產(chǎn)物; 2. pET-32a 雙酶切產(chǎn)物; 3. 豬α干擾素
2.2 重組蛋白的表達(dá)及鑒定
將重組質(zhì)粒轉(zhuǎn)入Rosetta(DE3)誘導(dǎo)表達(dá),超聲破碎后,SDS-PAGE檢測(cè)(如圖3)結(jié)果顯示,蛋白分子量約33 kDa,符合預(yù)測(cè)結(jié)果,蛋白表達(dá)形式為包涵體。電泳結(jié)束后,將蛋白轉(zhuǎn)移到硝酸纖維素膜上,與豬α干擾素單克隆抗體和羊抗鼠HRP抗體反應(yīng),結(jié)果顯示在33 kDa處顯示一條清晰的特異性條帶,與目的蛋白相對(duì)分子質(zhì)量一致(如圖4)。
2.3 內(nèi)毒素檢測(cè)
利用內(nèi)毒素檢測(cè)試劑盒,檢測(cè)純化后干擾素蛋白內(nèi)毒素的含量。檢測(cè)結(jié)果顯示,豬α干擾素突變體和參考蛋白的內(nèi)毒素(0.8 mg/mL)的檢測(cè)平均值分別為0.59 EU/mL和0.62 EU/mL,符合2015版《中國(guó)藥典》中通則1143要求(如圖5)。
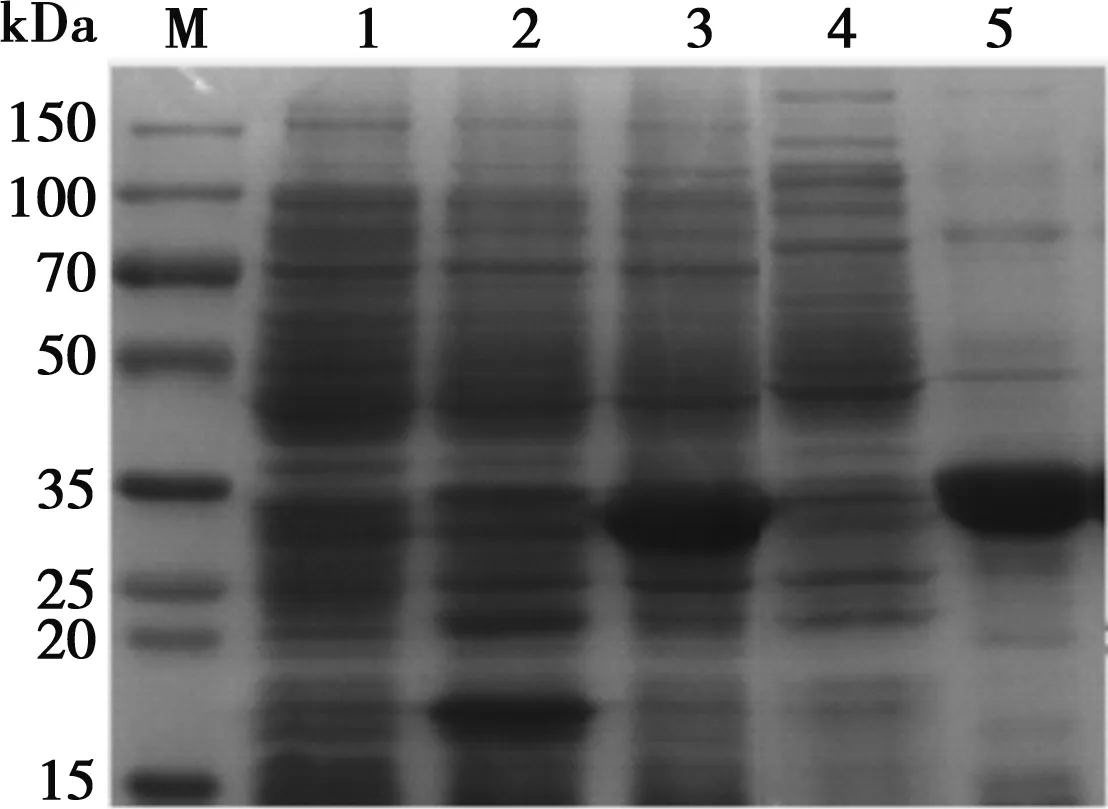
M. 蛋白Marker;1. Rosetta;2. pET-32a空載體誘導(dǎo);3.重組質(zhì)粒pET-32a--IFN-α誘導(dǎo);4. 沉淀中的重組蛋白;5. 上清中的重組蛋白
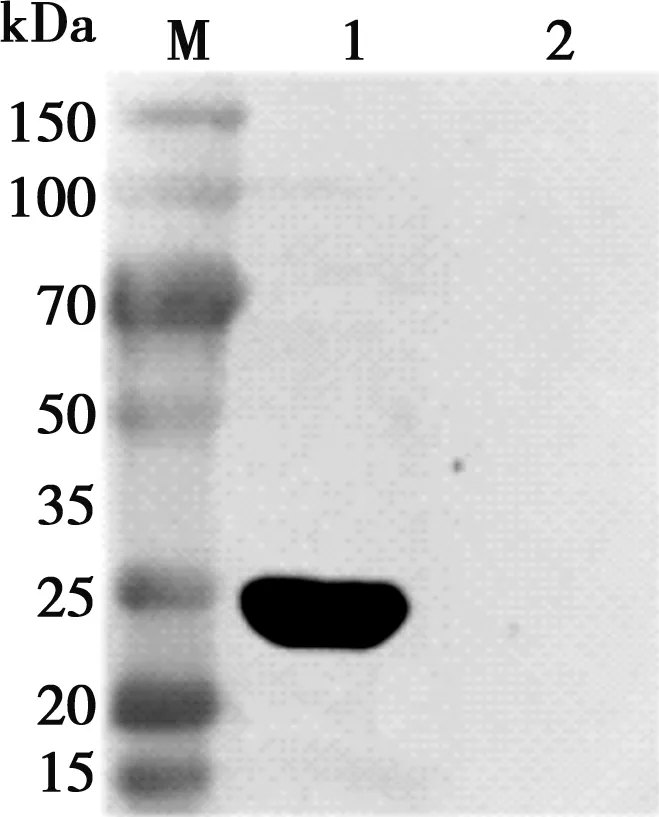
M. 蛋白Marker;1. 重組質(zhì)粒pET-32a-IFN-α誘導(dǎo);2. pET-32a空載體誘導(dǎo)
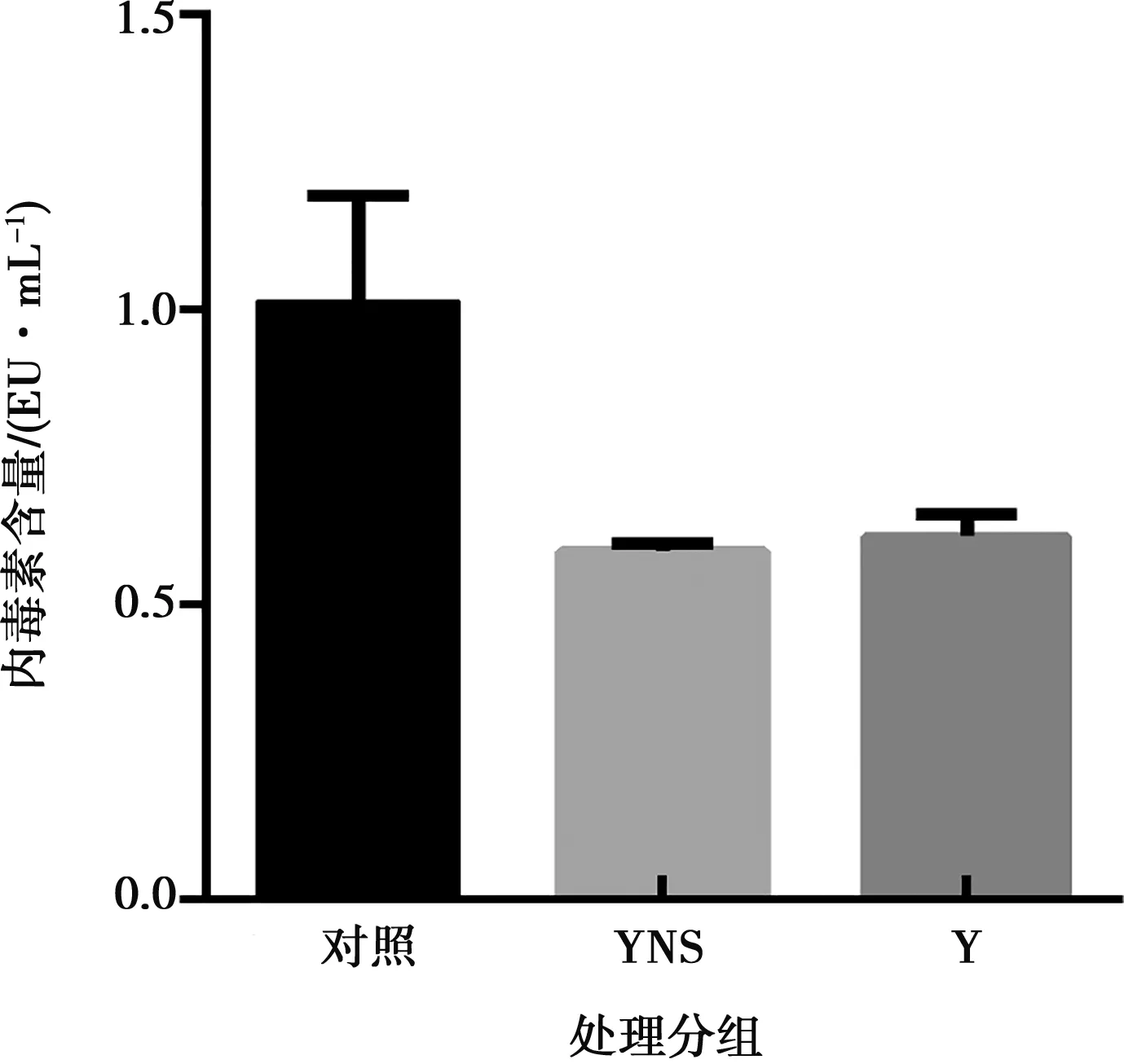
注:Y表示參照序列蛋白,YNS 表示豬α干擾素YNS突變體。下同
2.4 蛋白生物活性測(cè)定結(jié)
2.4.1 干擾素細(xì)胞安全濃度檢測(cè)
利用MTT法,在Vero細(xì)胞上檢測(cè)干擾素蛋白的安全濃度。結(jié)果表明,豬α干擾素突變體的CC50為462.7 μg/mL,參考序列蛋白的CC50為402.9 μg/mL,二者對(duì)細(xì)胞活性的影響呈現(xiàn)濃度依賴性,倍比稀釋到約25 μg/mL后對(duì)細(xì)胞增殖沒有影響(如圖6)。
2.4.2 抗病毒活性檢測(cè)結(jié)果
采用細(xì)胞病變抑制法,在Vero/VSV系統(tǒng)測(cè)定豬干擾素的抗病毒活性。檢測(cè)結(jié)果表明,豬α干擾素突變體表現(xiàn)出較高的抗病毒活性,突變體蛋白活性為1.63×106IU/mg。利用TCID50和Real-time PCR測(cè)定病毒滴度和基因拷貝數(shù),結(jié)果顯示豬α干擾素能夠有效地抑制VSV增殖,與參考序列蛋白相比差異顯著,具有更高的生物活性。
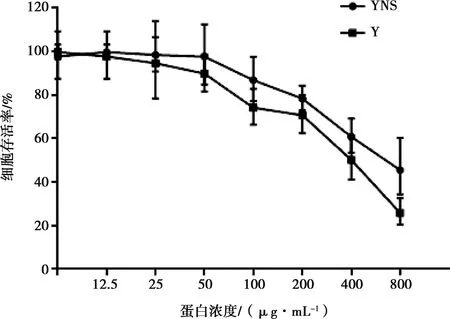
圖6 細(xì)胞活性檢測(cè)
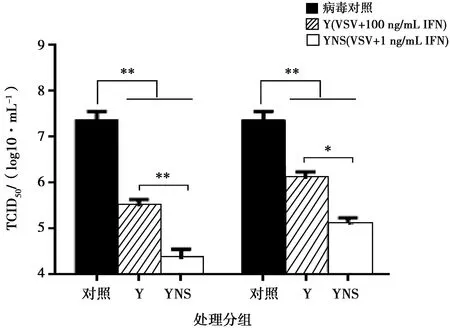
注:*表示差異顯著(P<0.05),**表示差異極顯著(P<0.01)。下同
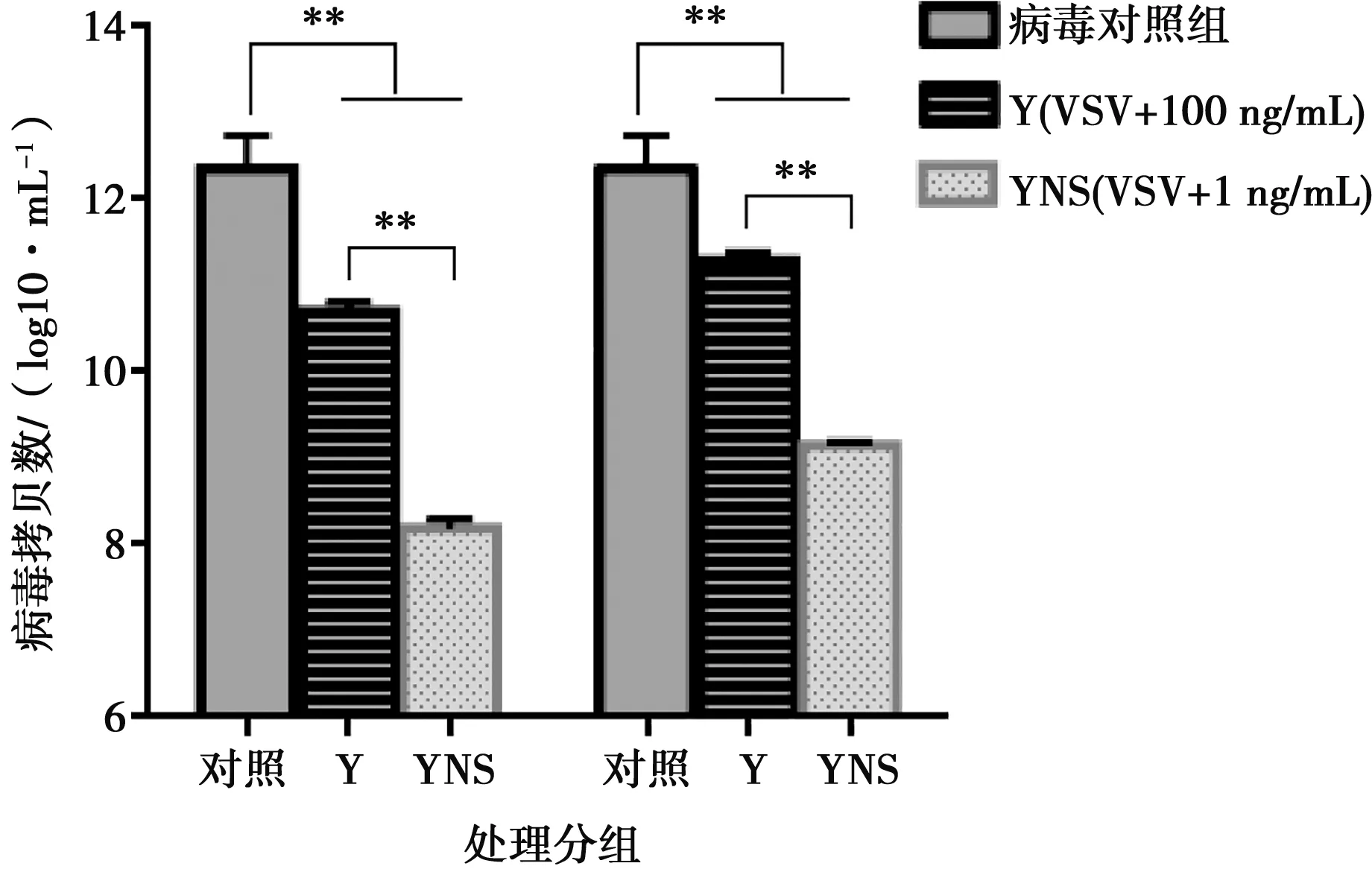
圖8 VSV基因拷貝數(shù)測(cè)定
3 討論
豬α干擾素具有速效廣譜的抗病毒效應(yīng),在預(yù)防和治療病毒性疾病方面發(fā)揮著不可替代的作用[14]。近年來(lái),國(guó)內(nèi)外許多學(xué)者利用原核、真核和桿狀病毒等表達(dá)系統(tǒng)對(duì)干擾素進(jìn)行表達(dá)[15-17]。Lefevre等[8]在大腸桿菌中表達(dá)豬α干擾素,并對(duì)其生物活性檢測(cè)。曹瑞兵等[18]對(duì)豬α干擾素密碼子進(jìn)行優(yōu)化,獲得干擾素蛋白的生物活性為6. 4×106IU/mg。王彥彬等[15]利用桿狀病毒表達(dá)系統(tǒng)表達(dá)干擾素,蛋白在細(xì)胞上有效抑制豬繁殖與呼吸綜合征病毒的增殖。Huang等[17]利用基因工程的方法改變豬α干擾素基因并在畢赤酵母中大量表達(dá),獲得了具有高活性的干擾素。
研究表明,改變?nèi)薎FNα2的3個(gè)氨基酸位點(diǎn)H57Y、E58N和Q61S,可使其活性相比野生型人IFN-α2得到提高[19]。人干擾素α和豬α干擾素同源性高達(dá)70%,因此本試驗(yàn)采用融合PCR的方法克隆包含58、59和61位氨基酸突變位點(diǎn)的豬α干擾素突變體基因,利用大腸桿菌系統(tǒng)表達(dá)突變體,優(yōu)化蛋白純化方式,縮短了純化時(shí)間,更加適合大規(guī)模制備和生產(chǎn)應(yīng)用。
本試驗(yàn)利用試劑盒檢測(cè)純化后的豬α干擾素內(nèi)毒素殘留,結(jié)果表明,內(nèi)毒素含量低于1 EU/mg,符合細(xì)胞因子制品標(biāo)準(zhǔn)。MTT試驗(yàn)結(jié)果表明突變體蛋白的抗增殖能力呈現(xiàn)濃度依賴性,在蛋白濃度為25 μg/mL時(shí),不影響Vero細(xì)胞增殖,對(duì)細(xì)胞無(wú)毒性。試驗(yàn)還運(yùn)用TCID50和Real-time PCR方法分別從病毒滴度和病毒基因拷貝數(shù)上探究豬α干擾素抑制VSV增殖的能力。結(jié)果表明,YNS突變體和參照序列蛋白均能顯著降低VSV的病毒滴度和基因拷貝數(shù),YNS突變體抑制病毒增殖作用更明顯,與參照序列蛋白相比降低約2個(gè)病毒滴度和2log10病毒基因拷貝數(shù)。
王彥彬等[16],通過對(duì)干擾素與其受體IFNAR2三級(jí)結(jié)構(gòu)分析,突變豬α干擾素與受體結(jié)合的氨基酸位點(diǎn),可以使豬α干擾素與受體更快、更緊密的結(jié)合,從而使干擾素活性增強(qiáng),但是具體某個(gè)氨基酸的改變對(duì)干擾素活性影響較大,仍需進(jìn)一步探究。
總之,本研究成功構(gòu)建了新型豬α干擾素YNS突變體,表達(dá)純化后具有較高的生物活性,為干擾素的臨床應(yīng)用提供理論基礎(chǔ)。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
