小麥耐冷相關(guān)基因TaCTR的克隆及其表達(dá)特性分析
2020-12-17 12:57:02張麗麗張小紅閔東紅
麥類作物學(xué)報(bào) 2020年9期
關(guān)鍵詞:分析
劉 煥,楊 卓,張麗麗,鄭 琪,張小紅,閔東紅
(1.西北農(nóng)林科技大學(xué)農(nóng)學(xué)院/生命學(xué)院,旱區(qū)作物逆境生物學(xué)國家重點(diǎn)實(shí)驗(yàn)室,陜西楊凌 712100; 2.陜西省農(nóng)牧良種場(chǎng),陜西寶雞 722203)
異三聚體G蛋白(Gα,Gβ/Gγ)偶聯(lián)受體簡(jiǎn)稱G蛋白偶聯(lián)受體(G-protein coupled receptors,GPCRs),是數(shù)量最大的一類膜蛋白受體,廣泛存在于動(dòng)物、真菌與植物中[1-3]。近年來在植物中,研究者主要在擬南芥[4-5]、水稻[6]、豌豆[7]、楊樹[8]、大豆[9]、蘋果[10]以及玉米[11]中發(fā)現(xiàn)GPCRs,并對(duì)其功能進(jìn)行了研究。前人研究結(jié)果表明,GPCRs參與植物的信號(hào)轉(zhuǎn)導(dǎo)過程并影響植物的生長發(fā)育,主要包括調(diào)控ABA信號(hào)通路[12]、參與CK信號(hào)轉(zhuǎn)導(dǎo)[13-14]、調(diào)節(jié)細(xì)胞周期蛋白、DNA合成[15]、磷脂酰肌醇[16]、藍(lán)光信號(hào)轉(zhuǎn)導(dǎo)[17]以及細(xì)菌信號(hào)交流[18]。GPCRs除了介導(dǎo)植物信號(hào)分子影響植物的生長發(fā)育之外,還參與植物逆境脅迫反應(yīng)。蘋果G蛋白偶聯(lián)受體基因MdGCR1在響應(yīng)植物抗旱脅迫中發(fā)揮重要作用[9];擬南芥GCR1參與多種生物和非生物脅迫響應(yīng),在磷酸鹽饑餓響應(yīng)中發(fā)揮作用[19],并會(huì)影響植物體內(nèi)脅迫基因的表達(dá)[17];水稻G蛋白偶聯(lián)受體基因OsGPCR1,可以提高植株的耐冷性、耐鹽性以及抗旱性[20],與其氨基酸序列同源性高達(dá)99.79%的同源基因COLD1,可以明顯的提高水稻的耐冷性[21]。目前,小麥GPCRs的研究主要集中在植株矮化[22]、禾谷鐮刀菌生長發(fā)育、毒性產(chǎn)生以及赤霉病菌致病等方面[23-24],而對(duì)該類基因能否提高小麥耐冷性尚未見報(bào)道。同時(shí),關(guān)于小麥G蛋白偶聯(lián)受體的作用機(jī)制亦不清楚,有待進(jìn)一步研究。
作物在生長過程中會(huì)受到生物脅迫和非生物脅迫,其中溫度、干旱以及鹽害等非生物脅迫是影響作物生產(chǎn)最為重要的因素[25]。近年來,由低溫引起的小麥倒春寒頻發(fā),對(duì)小麥生產(chǎn)造成嚴(yán)重的威脅,已成為小麥產(chǎn)量損失的主要災(zāi)害之一。因此,挖掘小麥耐冷基因,研究小麥耐冷的分子機(jī)制,對(duì)于合理利用小麥耐冷基因,開展小麥耐冷基因工程改良,確保小麥穩(wěn)產(chǎn)、高產(chǎn)具有重要現(xiàn)實(shí) 意義。
本研究采用同源克隆的方法得到水稻耐冷基因COLD1的同源基因TaCTR,利用生物信息學(xué)的方法對(duì)基因結(jié)構(gòu)、蛋白質(zhì)特性以及進(jìn)化關(guān)系進(jìn)行初步分析,通過亞細(xì)胞定位技術(shù)對(duì)該基因表達(dá)的蛋白進(jìn)行定位,利用qRT-PCR技術(shù)分析TaCTR在不同處理下的表達(dá)特性以及組織特異性等,以期為探究TaCTR的功能機(jī)制奠定基礎(chǔ)。
1 材料與方法
1.1 供試材料
供試普通小麥品種小偃22、周麥18、西農(nóng)979、13(36)0-7、西農(nóng)626、西農(nóng)611均由本實(shí)驗(yàn)室保存,其中,小偃22是本研究的主要材料,其具有耐冷性突出、高產(chǎn)穩(wěn)產(chǎn)和適應(yīng)性廣等優(yōu)良性狀。
1.2 方法
1.2.1 材料處理
將飽滿的小偃22種子于2017年10月播種于田間,待發(fā)育至拔節(jié)期,選擇發(fā)育正常的植株移栽至室內(nèi)花盆,緩苗一周,待幼穗發(fā)育進(jìn)入藥隔形成期至雌雄蕊分化時(shí)期,將其放入4 ℃春化培養(yǎng)間,分別在4 ℃冷處理0、0.5、1、2、5、8、12和24 h后,取根、莖、葉、幼穗等組織材料。將所取樣品迅速收集于2 mL離心管內(nèi)并置于液氮中,然后保存于-80 ℃冰箱中,用于后續(xù)熒光定量以及基因克隆。小麥拔節(jié)期取其雌雄蕊用于后續(xù)的組織 定量。
1.2.2 總RNA提取及cDNA第一鏈的合成
小麥組織的總RNA提取采用TRNzol Universal總RNA提取法,具體提取步驟參照TRNzol Universal總RNA提取試劑(DP424)(天根,北京)說明書。用1.2%的瓊脂糖凝膠電泳(200 V,10 min)檢測(cè)提取總RNA的完整性,用NanoDropTMOne/OneC 超微量紫外分光光度計(jì)(賽默飛,美國)測(cè)定RNA的濃度和純度。按照反轉(zhuǎn)錄試劑盒FastKing RT Kit(With gDNase)(KR116)(天根,北京)說明書將RNA 反轉(zhuǎn)錄合成cDNA,于-20 ℃保存?zhèn)溆谩?/p>
1.2.3 小麥TaCTR基因克隆
以水稻COLD1基因的cDNA序列為探針在 NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)與六倍體小麥進(jìn)行比對(duì),得到開放閱讀框長度為 1 407 bp的序列,利用Primer 5.0設(shè)計(jì)引物TaCTR-F(5′-CGAGAAGCGAAGGGGGAGA-3′)/ TaCTR-R(5′-AACCCCCTTTGTGAACT GACTTTA-3′)。以小麥幼穗cDNA為模板,利用PrimeSTAR○RHS DNA Polymerase with GC Buffer(R044A)(TaKaRa,日本)進(jìn)行PCR擴(kuò)增。擴(kuò)增體系及其操作步驟詳見說明書,其中循環(huán)數(shù)為35,退火溫度為59.5 ℃,延伸時(shí)間為1 min 45 s。PCR擴(kuò)增產(chǎn)物經(jīng)1.0%瓊脂糖凝膠電泳檢測(cè)后,利用瓊脂糖凝膠回收試劑盒 (DP219)(天根,北京)純化回收。利用PLB零背景快速連接試劑盒(VT205)(天根,北京)將目的基因連接到載體PLB載體上,轉(zhuǎn)化DH5a感受態(tài)細(xì)胞(天根,北京),挑取單克隆進(jìn)行陽性檢測(cè)并測(cè)序。引物合成及測(cè)序均在奧科鼎盛生物科技有限公司完成。
1.2.4 生物信息學(xué)分析
以小麥TaCTR的氨基酸序列為探針,在NCBI數(shù)據(jù)庫 BLAST,下載與其相似度>85%的其他物種的同源蛋白序列,用MEGA 7.0構(gòu)建系統(tǒng)進(jìn)化樹;用MEME 5.1.0(http://meme-suite.org/tools/meme )分析氨基酸保守序列,并用evolview-v2(https://evolgenius.info//evolview-v2)整合進(jìn)化樹與氨基酸保守序列;用SwissProt(https://swissmodel.expasy.org/)和ExPASy(https://web.expasy.org/cgi-bin/compute_pi/pi_tool )在線分析TaCTR蛋白的理化性質(zhì);用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html )工具預(yù)測(cè)TaCTR蛋白的二級(jí)結(jié)構(gòu);用WoLF PSORT(https://wolfpsort.hgc.jp/)進(jìn)行亞細(xì)胞定位;用TMHMM Server v.2.0 (http://www.cbs.dtu.dk/services/TMHMM/)和NetPhos 3.1 Server(http://www.cbs.dtu.dk/services/NetPhos/)預(yù)測(cè)跨膜結(jié)構(gòu)域和磷酸化位點(diǎn)分析;用SMART(http://smart.embl-hei-delberg.de/)在線分析TaCTR蛋白的功能域。
1.2.5TaCTR基因的亞細(xì)胞定位
對(duì)亞細(xì)胞定位載體pCAMBIA1302-EGFP進(jìn)行BstBI單酶切,酶切產(chǎn)物回收,利用引物TaCTR-GFP-F(5′-CGAGCTCAAGCTTCGAA ATGGGGTGGGGCGTGGTG-3′)/TaCTR-GFP-R(5′-CGTCGACTGCAGAATTCGAAATCAATT GGATGCTT-3′)擴(kuò)增目的基因。按照ClonExpressTMOne Step Cloning Kit試劑盒說明書(諾唯贊,南京)將酶切產(chǎn)物與TaCTR目的片段連接,轉(zhuǎn)化于DH5α感受態(tài)細(xì)胞,挑取單克隆進(jìn)行陽性檢測(cè)并測(cè)序。將重組質(zhì)粒轉(zhuǎn)至農(nóng)桿菌GV3101,挑取單克隆,取1 mL菌液置于50 mL含相應(yīng)抗生素的LB液體培養(yǎng)基中,28 ℃、220 r·min-1振蕩培養(yǎng) 16~18 h,使OD600=1.0; 5 000 r·min-1離心5 min;用懸浮緩沖液分別懸浮pCAMBIA1302-EGFP-TaCTR和pCAMBIA 1302-EGFP菌液沉淀,使OD600=0.6~0.8,室溫靜置3 h,對(duì)生長4周的本氏煙草進(jìn)行全葉片注射; 60 h后制備已注射菌液煙草的原生質(zhì)體,觀察蛋白表達(dá)。
1.2.6TaCTR基因的表達(dá)模式分析
根據(jù)TaCTR基因cDNA序列,避開TaCTR的保守區(qū),利用Primer 5.0設(shè)計(jì)引物RT-TaCTR-F(5′-CCTCCCCTACTACCACTGCT-3′)/RT-TaCTR-R(5′-CGCCAATCCTACTAA CCAAC-3′),用于檢測(cè)TaCTR的表達(dá);以β-Actin(AB181991)為內(nèi)參基因,所用引物為Actin-F(5′-CGATTCAGAGCAGCGTATTGTTG-3′)/Actin-R(5′-AGTTGGTCGGGTCTCTTCTAA ATG-3′)。將反轉(zhuǎn)錄得到的cDNA模板濃度均稀釋到200 ng·μL-1,利用SuperReal熒光定量預(yù)混試劑彩色版(SYBR Green)(FP215)試劑盒進(jìn)行定量分析,在熒光定量PCR儀(ABI7500)上進(jìn)行實(shí)時(shí)熒光定量PCR 反應(yīng)。反應(yīng)體系及程序參見說明書。每個(gè)反應(yīng)3次重復(fù),采用2-ΔΔCT方法分析TaCTR在不同小麥品種中的表達(dá)、組織表達(dá)特異性以及脅迫和激素處理后的相對(duì)表達(dá)量。
2 結(jié)果與分析
2.1 小麥耐冷基因TaCTR的克隆結(jié)果
對(duì)4 ℃冷處理小偃22幼穗組織進(jìn)行熒光定量分析發(fā)現(xiàn),在冷處理5 h內(nèi),TaCTR基因的表達(dá)量處于上升趨勢(shì),5 h時(shí)表達(dá)量達(dá)到最大,大約是0 h的11.16倍,存在顯著性差異,之后逐漸下降(圖1)。以冷處理5 h的小偃22幼穗的cDNA為模板,利用引物TaCTR-F/TaCTR-R進(jìn)行擴(kuò)增,擴(kuò)增產(chǎn)物用1.0%的瓊脂糖凝膠電泳檢測(cè),得到一個(gè)片段大小為1 407 bp的特異性條帶(圖2)。將目的片段回收后,連接至PLB零背景克隆載體,轉(zhuǎn)化于大腸桿菌后挑取單克隆進(jìn)行菌液檢測(cè),并對(duì)陽性單克隆進(jìn)行測(cè)序分析。測(cè)序結(jié)果經(jīng)BLAST分析表明,TaCTR分別位于小麥2A、2B和2D染色體上,并且與2D染色體上的序列完全相同。TaCTR基因序列全長2 192 bp,開放閱讀框長1 407 bp,共編碼468個(gè)氨基酸殘基,分子量為53.43 kDa。DNAMAN序列比對(duì)分析(圖3)發(fā)現(xiàn),TaCTR在2A、2B和2D染色體間具有高度保守性,且功能域相同,三者所編碼的氨基酸序列同源性高達(dá)99.64%,僅在第5、92位存在差異,在第5位(紅色邊框),2A染色體上對(duì)應(yīng)的為蘇氨酸,2B和2D染色體上為纈氨酸;第92位(藍(lán)色邊框),2B染色體上對(duì)應(yīng)的為纈氨酸,2A和2D染色體上為亮氨酸。因此,推斷該基因在2A、2B和2D染色體上具有相似的功能,本研究以2D染色體上的基因進(jìn)行后續(xù)試驗(yàn)。
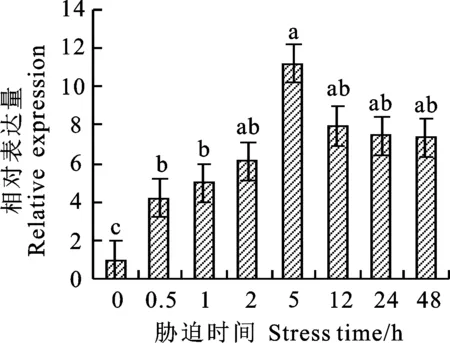
圖柱上不同小寫字母表示不同脅迫時(shí)間之間存在顯著性差異(P<0.05)。
2.2 TaCTR基因的生物信息學(xué)分析
2.2.1TaCTR基因編碼蛋白的系統(tǒng)發(fā)育進(jìn)化分析
蛋白系統(tǒng)進(jìn)化分析結(jié)果(圖4)表明,單子葉植物與雙子葉植物分別聚為一類,小麥TaCTR蛋白與單子葉植物處于同一進(jìn)化分支,且與粗山羊草的親緣關(guān)系最近,這可能是由于小麥的2D染色體來自于粗山羊草的緣故。分析不同物種間CTR蛋白的氨基酸保守序列發(fā)現(xiàn),無論雙子葉還是單子葉植物,CTR蛋白均含有motif 1、2、3、4和5,而且這5個(gè)motif的大小和位置接近,這說明TaCTR基因在不同物種間具有較好的保 守性。
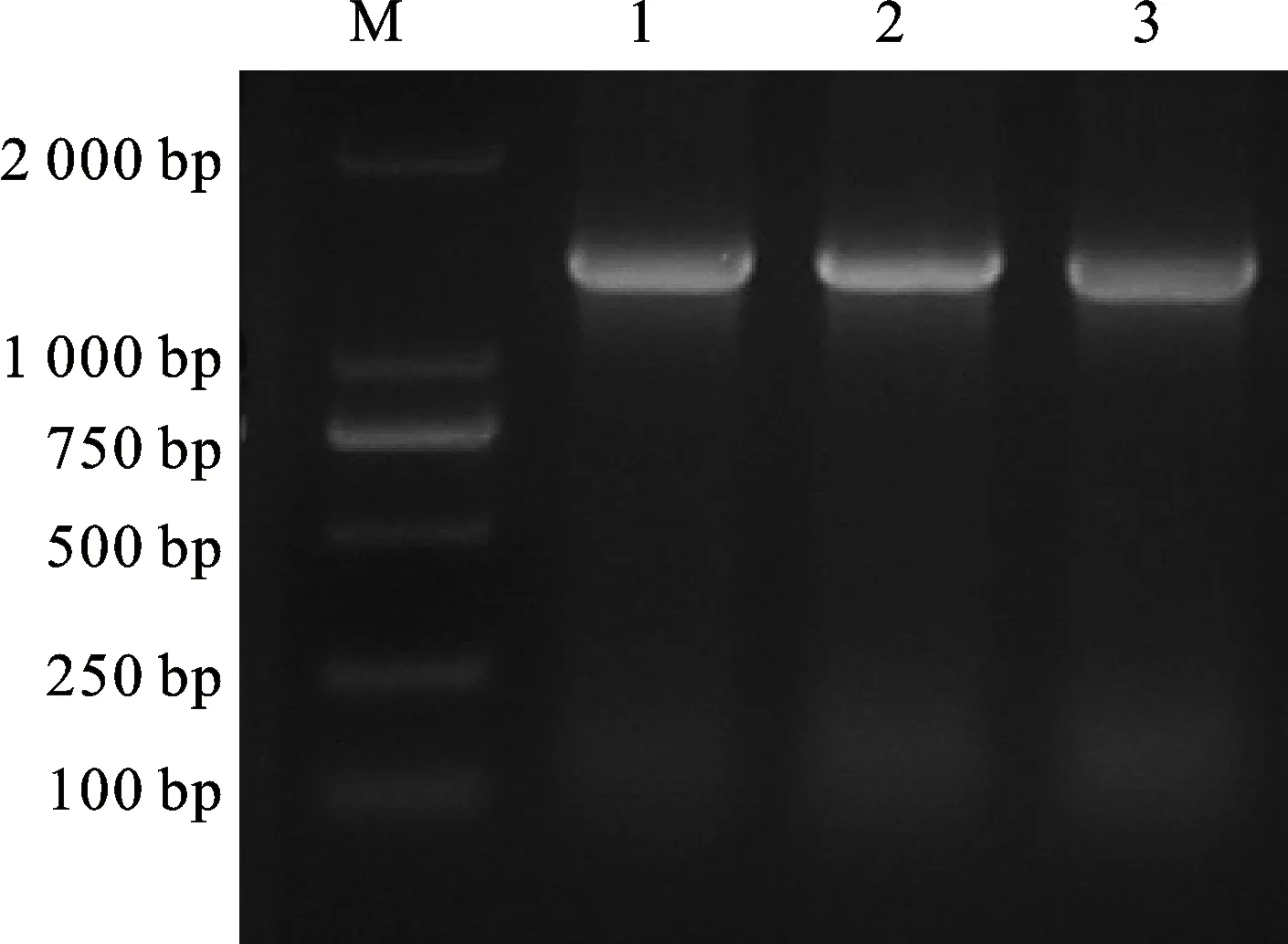
M:DL2000;1~3:TaCTR PCR產(chǎn)物。

紅色邊框:第5位氨基酸;藍(lán)色邊框:第92位氨基酸;紅線:GPHR_N功能域;藍(lán)色:ABA_GPCR功能域。
2.2.2TaCTR基因編碼蛋白的理化性質(zhì)及二級(jí)結(jié)構(gòu)分析
TaCTR蛋白分子式為C2488H3886N622O643S21,分子量為53.43 kDa,理論等電點(diǎn)PI為9.23,屬于堿性蛋白。總平均親水性值為 0.462,表明該蛋白屬于疏水性蛋白;不穩(wěn)定系數(shù)為40.54,說明該蛋白為不穩(wěn)定蛋白。通過SOPMA在線預(yù)測(cè)分析TaCTR的二級(jí)結(jié)構(gòu),發(fā)現(xiàn)該蛋白主要包括α螺旋(alpha helix),無規(guī)則卷曲(random coil)、延伸直鏈(extended strand)和β折疊(beta turn)四種結(jié)構(gòu),其所占比例分別為66.88%、20.09%、 9.62%和3.42%。
2.2.3TaCTR基因編碼蛋白的跨膜區(qū)、結(jié)構(gòu)域及磷酸化位點(diǎn)分析
通過TMHMM Server V.20在線預(yù)測(cè),發(fā)現(xiàn)TaCTR蛋白存在9個(gè)跨膜區(qū),屬于膜蛋白,每個(gè)跨膜區(qū)含20~23個(gè)氨基酸,對(duì)應(yīng)位置分別為6~28 aa、41~63 aa、78~100 aa、113~132 aa、152~174 aa、297~319 aa、347~369 aa、390~409 aa、433~455 aa。通過SMART在線分析,發(fā)現(xiàn)TaCTR蛋白主要包括GPHR_N和ABA_GPCR兩個(gè)功能域,位置分別在143~210 aa和285~458 aa之間,長度為68 bp和174 bp。通過NetPhos 3.1 Server數(shù)據(jù)庫分析,發(fā)現(xiàn)TaCTR蛋白含有16個(gè)絲氨酸、6個(gè)蘇氨酸和1個(gè)酪氨酸磷酸化位點(diǎn)。
2.3 TaCTR蛋白的亞細(xì)胞定位
通過WoLF PSORT對(duì)TaCTR蛋白進(jìn)行定位預(yù)測(cè),發(fā)現(xiàn)其主要存在于細(xì)胞膜上。將含有重組載體pCAMBIA 1302-EGFP-TaCTR和空載體pCAMBIA 1302-EGFP的菌液同時(shí)注射煙草,并制備煙草原生質(zhì)體進(jìn)行觀察。結(jié)果(圖5)表明,陽性對(duì)照pCAMBIA 1302-EGFP在細(xì)胞膜、細(xì)胞質(zhì)和細(xì)胞核均能觀察到熒光信號(hào),含有目的基因TaCTR的重組載體pCAMBIA 1302-EGFP-TaCTR僅在細(xì)胞膜上具有較強(qiáng)的熒光信號(hào),進(jìn)一步證明TaCTR編碼的蛋白為膜蛋白。
2.4 TaCTR基因在不同小麥品種中的表達(dá)及組織特異性分析
分別取生長至三葉期的13(36)0-7、西農(nóng)626、西農(nóng)611、小偃22、周麥18、西農(nóng)979的幼苗葉片,對(duì)TaCTR的相對(duì)表達(dá)量進(jìn)行分析,結(jié)果發(fā)現(xiàn),其相對(duì)表達(dá)量依次分別為1.00、1.56、1.64、3.71、5.19、 1.61, 說明在小偃22和周麥18中TaCTR表達(dá)水平較高,這也進(jìn)一步說明了小偃22具有較好的耐冷性,與多年的田間觀察結(jié)果相吻合。組織特異性表達(dá)水平(表1)顯示,TaCTR基因在小偃22的根、莖、葉、小穗以及雌雄蕊中均有表達(dá),但在小穗和雌蕊中表達(dá)量較高,這與小麥表達(dá)網(wǎng)站W(wǎng)heat Expression Browser(http://www.wheat-expression.com/)預(yù)測(cè)結(jié)果一致。

圖4 TaCTR蛋白的系統(tǒng)進(jìn)化樹(A)及氨基酸保守基序分析(B)Fig.4 Phylogenetic tree(A) and amino acid conserved motifs(B) analysis of TaCTR protein
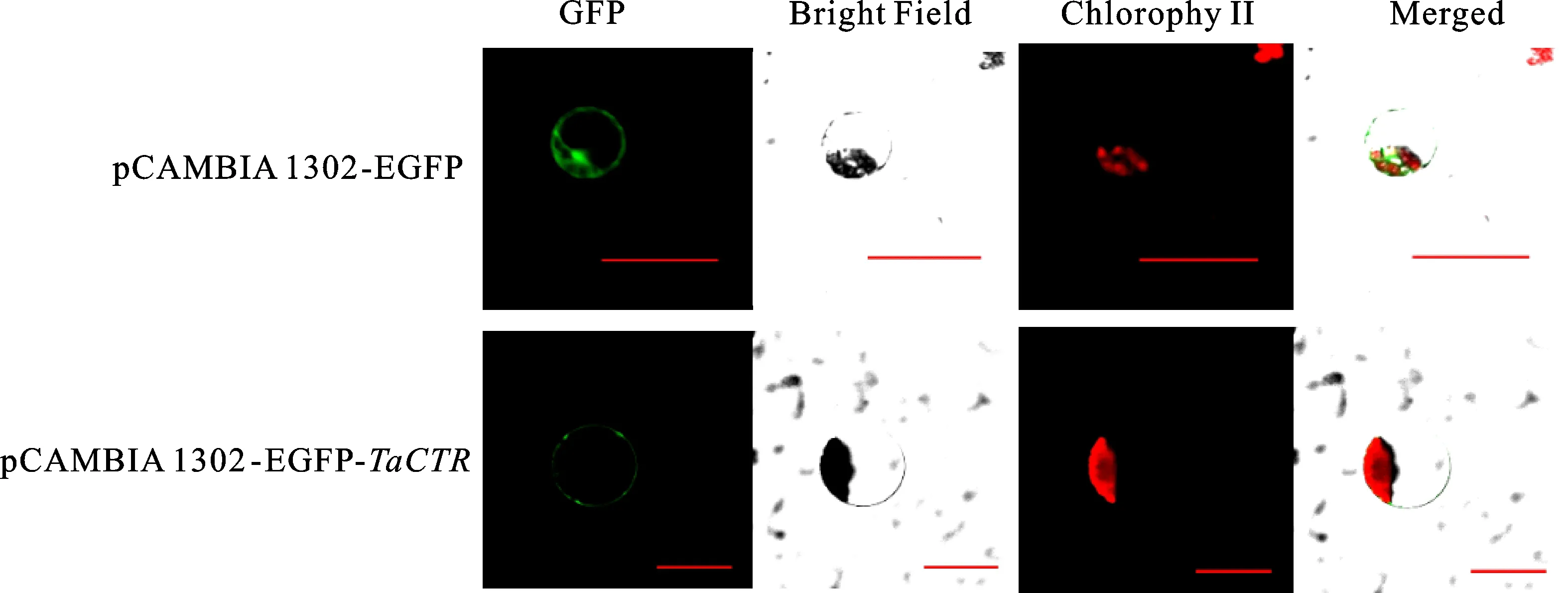
Bar=50 μm圖5 TaCTR蛋白亞細(xì)胞定位Fig.5 Subcellular localization of TaCTR
2.5 非生物脅迫及激素處理對(duì)TaCTR基因表達(dá)量的影響
由表2可以看出,在NaCl脅迫處理下,TaCTR的表達(dá)量在0~4 h間隨時(shí)間推移呈上升趨勢(shì),在處理4 h時(shí)表達(dá)量達(dá)到最大水平,為0 h的3.42倍,二者差異達(dá)到顯著水平(P<0.05);4 h后,其表達(dá)量逐漸下降。在干旱處理下,TaCTR的相對(duì)表達(dá)量在處理48 h時(shí)最高,約是0 h的 2.05倍,二者差異達(dá)到顯著水平。在GA處理下,TaCTR的表達(dá)量在1 h時(shí)達(dá)到最高水平,為0 h的3.13倍,二者差異達(dá)到顯著性差異。以上結(jié)果說明,TaCTR的表達(dá)受干旱以及鹽脅迫的誘導(dǎo),并且可能參與GA信號(hào)通路,因此,推測(cè)TaCTR參與植物的抗逆信號(hào)轉(zhuǎn)導(dǎo)。
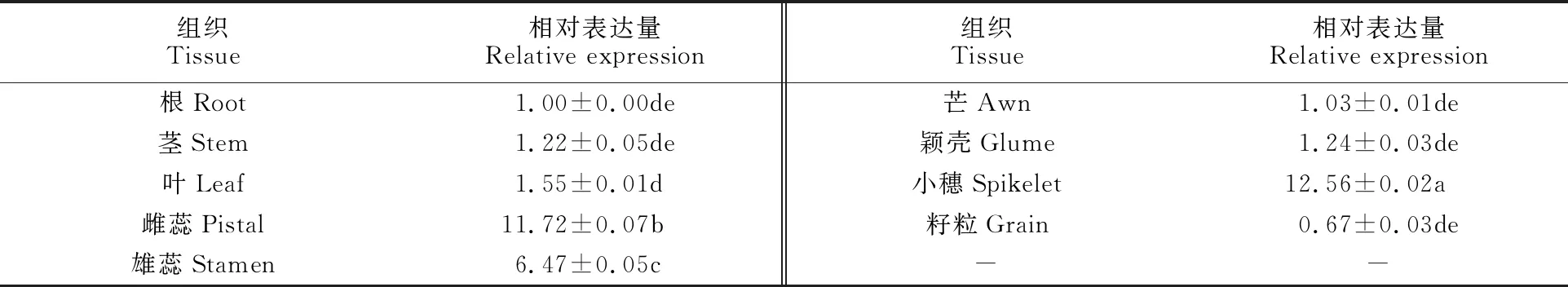
表1 小偃22中TaCTR基因的組織特異性分析Table 1 Tissue-specific expression analysis of TaCTR in Xiaoyan 22
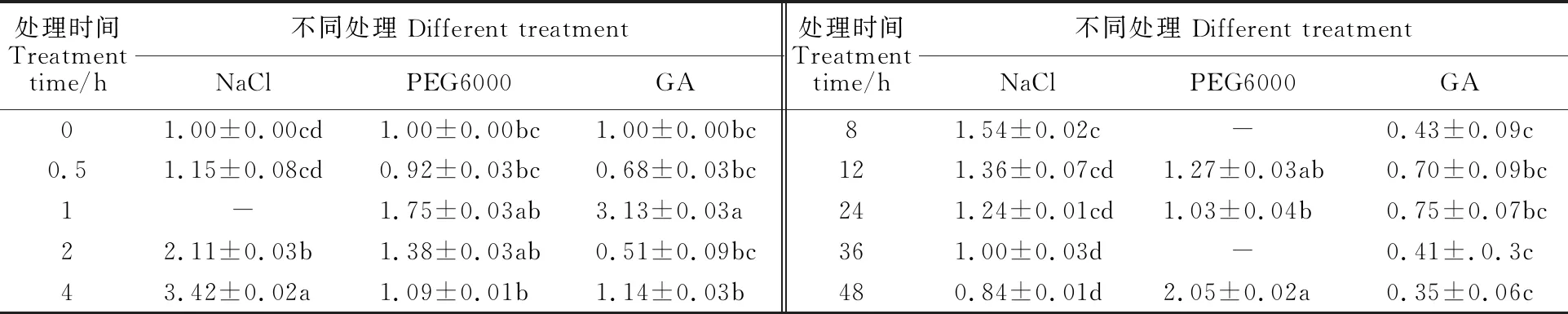
表2 不同處理對(duì)小偃22葉片中TaCTR基因表達(dá)量的影響Table 2 The effect of TaCTR expression in leaf of Xiaoyan 22 under different treatments
3 討 論
低溫、高溫、干旱以及鹽害是限制小麥生產(chǎn)的幾個(gè)主要非生物脅迫因子。其中,低溫冷害(春季倒春寒)已成為中國黃淮麥區(qū)影響小麥生產(chǎn)的重要災(zāi)害之一。春季倒春寒常發(fā)生在2月下旬至4月上旬,大田小麥正處于拔節(jié)期,其穗分化進(jìn)程正處于雌雄蕊分化期和藥隔發(fā)育時(shí)期,此時(shí)正是小麥幼穗對(duì)低溫較為敏感的時(shí)期[26],也是小麥產(chǎn)量形成的關(guān)鍵時(shí)期。倒春寒的發(fā)生會(huì)導(dǎo)致小花停止發(fā)育、幼穗凍死,或麥穗上部和下部受凍嚴(yán)重,因而結(jié)實(shí)率明顯降低,造成小麥減產(chǎn)10%~50%左右[27]。
研究發(fā)現(xiàn),受到低溫脅迫時(shí),水稻COLD1基因與G蛋白α亞基相互作用,激活Ca2+通道,并增強(qiáng)G蛋白GTPase的活性,從而保護(hù)水稻避免低溫脅迫[21]。本研究利用同源克隆方法從小偃22中分離到水稻耐冷基因COLD1的同源基因TaCTR,兩者高度同源,均具有9個(gè)典型的跨膜結(jié)構(gòu)域。TaCTR基因序列與中國春序列比對(duì)結(jié)果發(fā)現(xiàn),兩者相似度達(dá)到100%,說明該基因在不同的小麥品種之間具有很高的保守性,而且該基因在2A、2B和2D三個(gè)染色體組之間編碼蛋白的同源性達(dá)到99.64%,說明該基因在不同染色體組間也具有高度保守性。由于異源六倍體小麥的三個(gè)染色體組的部分同源基因可能是通過表達(dá)的差異來調(diào)控小麥的生長發(fā)育,其調(diào)控網(wǎng)絡(luò)復(fù)雜[28-29],加之小麥基因可變剪切較為普遍,且涉及小麥發(fā)育、生理生化代謝和逆境脅迫響應(yīng)等多個(gè)過程,導(dǎo)致異源六倍體小麥存在多個(gè)等位基因位點(diǎn),致使小麥基因表達(dá)調(diào)控十分復(fù)雜[30-31],關(guān)于TaCTR基因在普通小麥的具體表達(dá)模式與調(diào)控機(jī)制還有待進(jìn)一步深入探討。
進(jìn)化樹分析結(jié)果顯示,小麥TaCTR基因在單雙子葉中都存在同源基因,暗示了該基因在單雙子葉分化之前就已存在。該基因與粗山羊草之間的遺傳關(guān)系最近,表明該基因在小麥進(jìn)化過程中由粗山羊草提供。至于該基因在單、雙子葉植物基因組中保守域無明顯差異,說明該基因在單、雙子葉分化演變中仍具有較高的保守性,但是具體的機(jī)制還需進(jìn)一步探究。
組織特異性表達(dá)結(jié)果發(fā)現(xiàn),TaCTR基因在小麥的各個(gè)部位均有不同程度的表達(dá),這與水稻G蛋白偶聯(lián)受體基因OsGPCR的表達(dá)模式[20]以及擬南芥中基因GTG1/2的GUS染色結(jié)果一致[32]。TaCTR基因在小麥的小穗以及雌雄蕊中表達(dá)量較高,說明小麥在遭受春季低溫脅迫時(shí),其小穗對(duì)低溫的反應(yīng)最為敏感,最先做出相應(yīng)的應(yīng)激反應(yīng)。亞細(xì)胞定位結(jié)果表明,該蛋白主要在細(xì)胞膜上,這與生物信息學(xué)分析結(jié)果以及水稻G蛋白偶聯(lián)受體基因OsGPCR的定位結(jié)果一致[20],推測(cè)該基因編碼的蛋白與水稻OsGPCR編碼的蛋白在功能上具有相似性。而植物受到低溫脅迫時(shí),細(xì)胞膜最先受到傷害,該基因定位于細(xì)胞質(zhì)膜上,這對(duì)其感受冷害信號(hào)具有重要意義。
Assman等[5]研究結(jié)果顯示,G蛋白偶聯(lián)受體是植物中存在的可能感受環(huán)境脅迫的受體分子。而TaCTR基因與水稻中編碼G蛋白偶聯(lián)受體的基因OsGPCR1具有高度同源性,并且與人體內(nèi)的G蛋白偶聯(lián)受體HsGCR68以及擬南芥中的GPCR型G蛋白有著共同的GPHR_N和ABA_GPCR結(jié)構(gòu)域,并且均含有9個(gè)典型跨膜結(jié)構(gòu)域。因此,推測(cè)TaCTR基因?yàn)樾←溨幸粋€(gè)潛在的G蛋白偶聯(lián)受體基因,在小麥的低溫信號(hào)轉(zhuǎn)導(dǎo)以及逆境脅迫中發(fā)揮著極其重要的作用。
在本研究中,對(duì)處于三葉期的小麥幼苗進(jìn)行非生物脅迫以及激素處理后進(jìn)行熒光定量試驗(yàn),結(jié)果顯示,TaCTR的表達(dá)在低溫、干旱以及高鹽脅迫時(shí)均上調(diào)表達(dá),這與水稻基因OsGPCR1研究結(jié)果一致[20]。在GA處理下,TaCTR的表達(dá)量明顯上調(diào),在水稻中發(fā)現(xiàn)COLD1與GA信號(hào)的關(guān)鍵元件D1/RGA1在功能上是相互關(guān)聯(lián)的[21],而TaCTR與COLD1有著高度的同源性,這也暗示著小麥TaCTR與水稻COLD1有著相似的調(diào)控機(jī)制。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
