扦插基質對葡萄扦插苗碳水平衡和苗木生長的影響
2020-12-23 04:26:36呂英忠代永欣王林張擁兵
山西果樹 2020年6期
關鍵詞:生長
呂英忠 代永欣 王林 張擁兵
摘?要:
以‘早黑寶葡萄扦插苗為研究對象,測定了基質扦插苗、混合園土苗和純園土苗的苗木生長、水勢、光合作用、非結構性碳(NSC)含量。結果表明:基質扦插苗的正午水勢顯著高于混合園土苗和純園土苗,而凌晨水勢在3個處理間沒有顯著性差異。基質扦插苗的光合作用、氣孔導度和蒸騰速率為3個處理中最高,且基質扦插苗的NSC含量最大,基質扦插苗的苗木生長情況在3個處理組中最好,而純園土扦插苗的以上指標均為3個處理中最差。以上結果表明:不同扦插基質會影響葡萄扦插苗的水分狀況、光合作用,進而影響苗木碳平衡的重建。選擇合適的基質有利于苗木的水-碳平衡和苗木生長,提高苗木質量,研究結果也表明:泥炭珍珠巖蛭石=211的育苗基質有利于葡萄苗的碳水平衡重建和苗木的生長。
關鍵詞:葡萄;扦插基質;生長;光合作用;碳水平衡
文章編號:2096-8108(2020)06-0020-05??中圖分類號:S663.1??文獻標識碼:A
Effects of Cutting Substrate on Carbon and
Water Balance and Growth of Grape Cutting
LV Yingzhong1, DAI Yongxin2, WANG Lin2, ZHANG Yongbing1*
(1.Pomology Institute, Shanxi Agricultural University, Taigu 030815, China;2 College of Forestry, Shanxi Agricultural University, Taigu 030801, China)
Abstract:This study used Zaoheibao Grape cuttings as material. They were planted in three different cutting substrates, growth matrix, mixed garden soil and pure garden soil. The growth, water potential, photosynthetic rate and nonstructural carbohydrates (NSC) content of the cuttings were measured. The midday water potential of the cuttings growing in growth matrix was significantly higher than those growing in mixed garden soil or pure garden soil, but there was no significant difference in predawn water potential among the three treatments. The photosynthetic rate, stomatal conductance and transpiration rate of the cuttings growing in growth matrix was the highest among the three treatments. They also had the greatest NSC content and the best growth state, while all of the indexes of the cuttings growing in pure garden soil were the worst. The results showed that different cutting substrate could influence water state and photosynthesis of the cuttings, then affect the reconstruction of carbon balance. Thus, choosing suitable substrate is beneficial to carbon and water balance and growth of cuttings and to improve the quality of seedlings. The results also showed that when the peatperlitevermiculite =211, seedling substrate was conducive to the reconstruction of the carbon and water balance and seedling growth.
Keywords:grape; substrate; growth; photosynthesis; carbon and water balance
扦插育苗中,基質是影響扦插苗木成活和生長的重要因素,基質中的固、液、氣比例以及微生物活動構成了植物的根際環境,基質的孔隙度大小會影響到內部的氣、水環境,基質成分的變化與苗木的水分、養分供應能力有較大關系[1-2],進而會影響到苗木的水分狀況和光合作用,影響苗木的水分平衡和碳代謝[3-4]。水分代謝和碳代謝是植物的兩種基礎代謝形式,在扦插成苗過程中,水分平衡和碳平衡是植物維持成活的基礎,而且兩者之間存在復雜的相互作用[5-6]。扦插過程中剪切插穗環節破壞了植物原有的水-碳平衡,植物通過自身的修復能力重新生長根和葉片,重新建立這種水分和碳水化合物平衡是植物能夠成活
且正常生長的基礎。但目前對扦插育苗過程中扦插基質對苗木水碳平衡重建的影響了解不足,這也限制了苗木培育生理基礎的完善。
葡萄(Vitis vinifera L.)是廣泛栽培的果樹之一,硬枝扦插是葡萄生產上主要的繁殖方式[7]。單芽扦插可以節省插穗,提高插穗的利用率,但目前我們關于扦插基質對扦插苗木生長過程中生理狀態的影響
尚了解不足,因此研究不同扦插基質條件下扦插苗的生長發育和生理狀態變化,有助于更加深入地了解不同扦插基質對苗木生長發育和生理機制的影響,從而為葡萄扦插育苗提供理論數據。本研究以葡萄扦插
育苗過程中扦插基質對苗木生長、水分狀況和碳素分配的影響為切入點,研究不同基質條件下葡萄單芽扦插苗的生理變化,以期為葡萄育苗生產提供技術依據。
1?材料與方法
1.1?試驗設計
試驗在山西省農業科學院果樹研究所大苗培育基地進行。2017年11月上旬采集1年生早黑寶葡萄枝條,于0 ℃左右土窖洞中貯藏,第2年3月取出枝條,選擇飽滿芽體剪切成5~6 cm長插穗,上方剪口距芽體約0.5~1 cm,剪好的插條在水中浸泡24 h后用IAA250P mg/L + IBA250 mg/L快速蘸插條生物學下端,放在25~27 ℃的電熱溫床上促根15 d催根;催根完成后分別在園土、混合園土和營養缽中扦插,園土為果園表層20 cm以上土壤,混合后過篩,混合園土為園土細沙牛糞=211,營養缽中的育苗基質為泥炭珍珠巖蛭石=211;扦插后保持園土、混合園土和基質的相對含水量在60%~80%,白天溫度20~28 ℃,夜間12~16 ℃;萌發后只保留生長勢較強的一個芽。
試驗設3個處理,每處理扦插苗木300株,100株為1個重復,共3個重復,在7月初(扦插后4個月)的連續晴天測定苗木的凌晨水勢、正午水勢、光合作用和NSC含量。
1.2?測定方法
成活率=成苗苗木數/扦插苗木總數;苗高用卷尺測量;最大葉片的葉面積用葉面積儀測量;苗木總根長用WinRHIZO STD1600根系掃描儀(加拿大REGENT公司)測量;凌晨和正午水勢用PMS 600D水勢儀(PMS Albany,USA)測定,凌晨水勢測定時間為大棚保溫被揭開前,正午水勢測定時間為12:00-14:00;光合作用、氣孔導度和蒸騰速率用 Li-6400 光合儀(Li-Cor,Lincoln,USA)測定;可溶性糖和淀粉含量參照Mitchell等[8]的方法測定;NSC含量=可溶性糖含量+淀粉含量。
1.3?統計分析
文中數據用SPSS22.0軟件進行統計分析,繪圖采用 SigmaPlot 10.0 軟件進行繪圖。
2?結果與分析
2.1?不同處理對葡萄扦插苗生長的影響
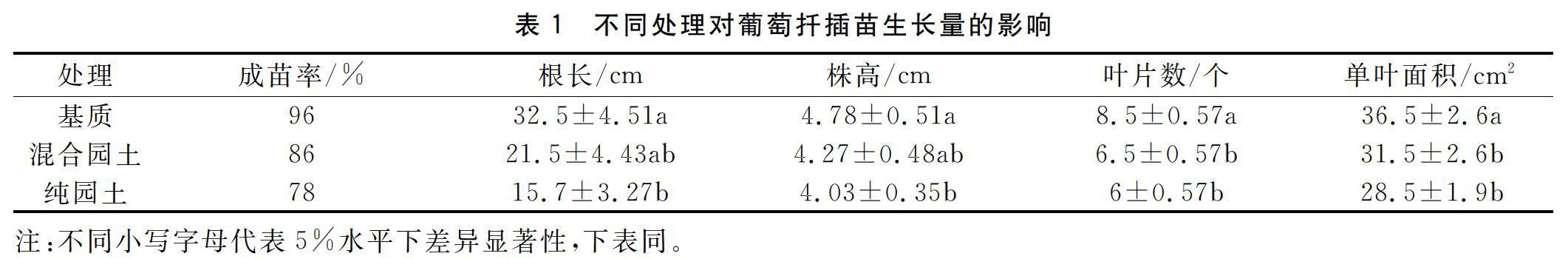
基質扦插苗的根長、株高顯著高于純園土苗(p<0.05),也高于混合園土苗,但與混合園土苗差異不顯著。基質扦插苗葉片數和葉面積均顯著高于混合園土苗和純園土苗(p<0.05),混合園土苗在根長、株高、葉片數和單葉面積上均略高于純園土苗,但二者之間差異不顯著。不同扦插基質的成苗率大小關系為基質>混合園土>純園土。
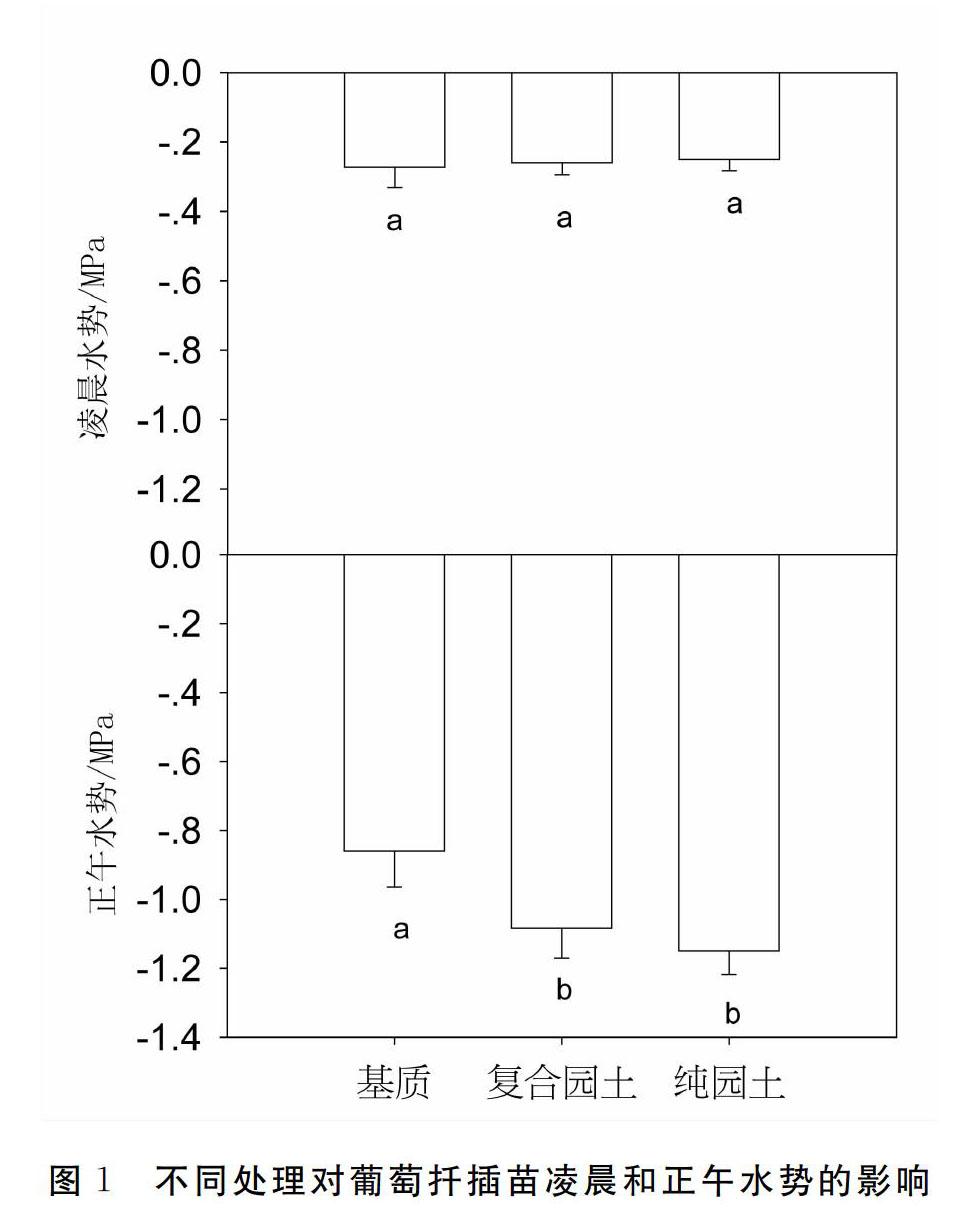
不同處理間在凌晨水勢上均沒有顯著性差異,基質扦插苗的正午水勢顯著高于混合園土和純園土扦插苗,基質扦插苗的正午水勢比混合園土苗和純園土苗高0.22 MPa和0.29 MPa。
2.3?不同處理對葡萄扦插苗光合作用的影響
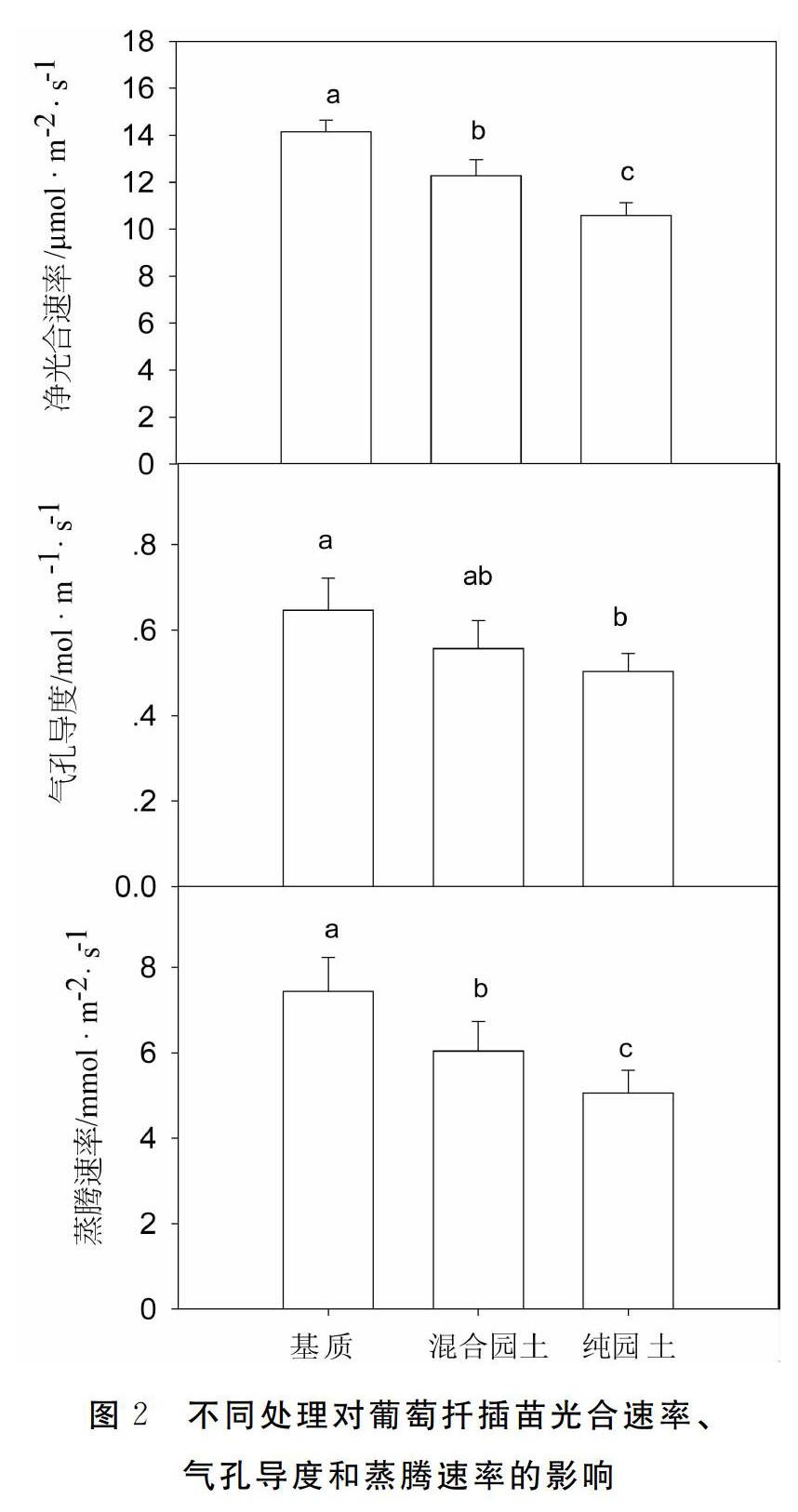
基質扦插苗的光合速率顯著高于混合園土苗和純園土苗(p<0.05),分別比混合園土苗和純園土苗高15.3%和33.7%,混合園土苗的光合速率又顯著高于純園土苗;基質扦插苗的氣孔導度顯著高于純園土苗(p<0.05),但與混合園土苗之間差異不顯著;基質扦插苗的蒸騰速率也顯著高于混合園土苗和純園土苗,分別比混合園土苗和純園土苗高23.1%和47.0%(圖2),混合園土苗的光合速率也顯著高于純園土苗。
2.4?育苗基質對葡萄扦插苗NSC含量的影響
基質扦插苗的可溶性糖含量顯著高于純園土苗(p<0.05),比純園土苗高37%,也略高于混合園土苗,但二者之間差異不顯著;基質扦插苗的淀粉含量顯著高于混合園土苗和純園土苗,分別比混合園土苗和純園土苗高41.5%和97.0%;基質扦插苗的NSC含量也顯著高于混合園土苗和純園土苗(p<0.05),分別比混合園土苗和純園土苗高23.8%和54.1%。混合園土苗的淀粉含量和NSC含量均顯著高于純園土苗,但可溶性糖含量二者之間差異不顯著。
3?討論
扦插苗培育過程中,不定根的發生是成苗的關鍵,而在這一過程中水分代謝和碳代謝的維持是扦插成苗的基礎。水分輸導功能維持的基礎是水力結構的完整性[9-11],因插條剪切破壞了葡萄枝條90%以上的木質部導管完整性,因此水分輸導功能較差,葡萄扦插苗未生根前,只能依靠剪切后的下端吸收水分,即使是新根發生后,較少量的新根對水分和養分的吸收能力也有限,因此當基質的供應能力不足時,苗木水分和養分的吸收就會受到限制,而不同基質對水分保持能力影響較大,進而影響苗木對水分的吸收。試驗結果表明不同處理間在凌晨水勢上沒有差異,而在正午水勢上,基質苗>混合園土苗>純園土苗,這表明基質苗在正午的水分狀況更好(圖1)。在凌晨,由于經歷了一夜的低蒸騰需求階段,苗木體內水分得到了很好的補充,因此不同處理間沒有差
異。而在正午,苗木旺盛的蒸騰需求使得不同基質造成的水分功能能力差別得到體現,即基質的持水力和水分供應能力最好,純園土最差,這主要與基質的持水能力有關,在研究的3種扦插基質中,基質(泥炭珍珠巖蛭石=211)有機質含量較高,且空隙度大,持水能力最好,水分供應能力的差別進而在苗木水分狀況上體現出了差別。這種更好的水分狀況進而影響到了苗木的光合作用,因此,基質扦插苗的光合速率、氣孔導度和蒸騰速率在3個處理中均最高,而園土苗的氣體交換指標在3個處理中均最低(圖2)。植物的光合作用不僅與水分狀況有關,也與養分狀況有關,前人的研究表明養分供應充足也會提高植物的光合能力,不同基質之間的養分狀況也有很大差別,基質的有機質含量高,養分供應能力也會高于園土,因此養分功能也造成了光合作用的差別[12-13]。
碳以不同形式的碳水化合物存在,是植物體的主要組成部分,在植物生長、滲透調節、防御等功能中發揮著重要的作用[14-15]。葡萄扦插苗未形成葉前,因無法進行光合作用,只能依靠插穗中儲藏的碳素營養生長發育[16-17],因此在扦插生根和長葉初期,插穗中的碳向根、芽部位運輸,促進發根和長葉。在扦插苗發芽長葉后,葉片能夠進行光合作用,這是植物的碳平衡在逐漸重建。而在這一扦插苗培育過程中,植物的碳平衡能否維持也是苗木成活的關鍵。我們的檢測結果表明3種基質扦插苗在NSC上存在顯著性差異,即基質扦插苗的NSC含量最高,純園土苗的NSC含量最低,這主要是因為在成苗時基質扦插苗由于光合能力更高(圖2)、葉面積更大(表1),造成基質扦插苗的碳攝取能力最強,因此能更好的維持扦插苗的碳素供應,進而有助于苗木的生長,因此基質扦插苗的生長情況也為3個處理中最好(表1)。而園土苗的光合作用較低,葉面積較小,這導致了碳攝取能力不足,進而影響到了苗木的碳平衡重建速度。我們之前的研究中也表明在扦插成苗過程中NSC含量呈現先下降后上升的趨勢,而本研究測定時期(扦插后4個月)屬于苗木NSC上升階段,因此基質對水分和養分的功能能力不足也限制了苗木碳平衡的重建過程。
不同基質間除在水分和養分的功能能力外,在土壤結構的形成,氧氣含量上也有差別[18],這也可能導致了根的發生、生長和吸收能力的差異。通常情況下,有機質含量更高會更有利于形成土壤良好的結構[18-19],維持水分、養分和氧氣的供應。在本研究選取的3種扦插基質中,基質的有機質含量最高,因此其土壤結構也應最好,這也是影響到苗木生理狀態和生長的重要因素。
4?結論
本研究比較了不同扦插基質下‘早黑寶葡萄單芽扦插苗木的生長發育、水分狀況、光合作用和NSC含量的積累等情況。研究表明,扦插苗生長4個月后,基質扦插苗的正午水勢較高,水分狀況較好,光合速率更高,苗木的NSC含量也更高,扦插苗的生長情況更好。純園土苗以上指標在3個處理中最差。這表明扦插基質會影響到苗木的水分平衡和碳平衡的維持,進而影響到扦插苗的成活和生長,因此在生產上,選擇合適的扦插基質能夠促進扦插快速成苗,提高苗木質量。研究結果也表明泥炭珍珠巖蛭石=211的育苗基質有利于葡萄苗的碳水平衡重建和苗木的生長。
參考文獻
[1]扈紅軍,曹幫華,尹偉倫,等.不同處理對歐榛硬枝扦插生根的影響及生根過程中相關氧化酶活性的變化[J].林業科學,2007(12):70-75.
[2]陳文軍,屈林豐,韓吉思,等.基質、插穗部位和生根劑對亞美馬褂木扦插生根的影響[J].內蒙古農業大學學報(自然科學版),2018,19(6):800-804.
[3]王?林,代永欣,郭晉平,等.刺槐苗木干旱脅迫過程中水力學失敗和碳饑餓的交互作用[J].林業科學,2016,52(6):1-9.
[4]Jim C Y, Peng L L H. Substrate moisture effect on water balance and thermal regime of a tropical extensive green roof[J]. Ecological Engineering, 2012, 47: 9-23.
[5]Reinhardt K, Germino M J, Kueppers L M, et al. Linking carbon and water relations to drought-induced mortality in Pinus flexilis seedlings[J].Tree physiology, 2015, 35(7): 771-782.
[6]代永欣,王?林,萬賢崇.遮蔭和環剝對刺槐,側柏苗木碳素分配和水力學特性的影響[J].林業科學,2017,53(7):37-44.
[7]文穎強,馮嘉玥,萬春雁.不同基質配比對幾種無核葡萄扦插生根的影響[J].中外葡萄與葡萄酒,2007(3):9-11.
[8]Yuan Y, Zhong M, Shu S, et al. Effects of exogenous putrescine on leaf anatomy and carbohydrate metabolism in cucumber (Cucumis sativus L.) under salt stress[J].Journal of Plant Growth Regulation, 2015,34(3):451-464.
[9]Zhao X H, Liu L Y, Nan L J, et al. Development of tyloses in the xylem vessels of Meili grapevine and their effect on water transportation[J]. Russian Journal of Plant Physiology, 2014,61(2):194-203.
[10]Sun Q, Rost T, Matthewe M. Wound-induced vascular occlusions in Vitis vinifera (Vitaceae): Tyloses in summer and gels in winter[J]. American Journal of Botany, 2008,95(12):1498-1505.
[11]Delpierre N, Vitasse Y, Chuine I, et al. Temperate and boreal forest tree phenology: from organ-scale processes to terrestrial ecosystem models[J]. Annals of Forest Science,2016, 73(1): 5-25.
[12]Morales F, Pavlovi A, Abadía A, et al. Photosynthesis in Poor Nutrient Soils, in Compacted Soils, and under Drought[M]//The Leaf: A Platform for Performing Photosynthesis. Springer, Cham, 2018:371-399.
[13]Ding X, Jiang Y, Zhao H, et al. Electrical conductivity of nutrient solution influenced photosynthesis, quality, and antioxidant enzyme activity of pakchoi (Brassica campestris L. ssp. Chinensis) in a hydroponic system[J]. PloS one, 2018, 13(8): e0202090.
[14]王?凱,雷?虹,夏?揚,等.楊樹幼苗非結構性碳水化合物對增加降水和氮添加的響應[J].應用生態學報,2017,28(2):399-407.
[15]Wiley E, Huepenbecker S, Casper B B, et al. The effects of defoliation on carbon allocation: can carbon limitation reduce growth in favour of storage[J]. Tree Physiology,2013,33(11):1216-1228.
[16]Xia Q, Wu W C, Tian K, et al. Effects of different cutting traits on bud emergence and early growth of the Chinese vegetable Toona sinensis[J]. Scientia Horticulturae, 2015,190:137-143.
[17]杜常健,孫佳成,陳?煒,等.側柏古樹實生樹和嫁接樹的扦插生理和解剖特性比較[J].林業科學,2019(9):5.
[18]丁奠元,馮?浩,趙?英,等.氨化秸稈還田對土壤孔隙結構的影響[J].植物營養與肥料學報,2016,22(3):650-658.
[19]宋蒙亞,吳?萌,劉?明,等.不同種植年限設施菜地土壤有機質組成與結構變化[J].土壤通報,2016,47(6):1386-1392.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
