利用重組自交系定位水稻種子低氧發芽力QTL
2021-01-08 07:13:44張所兵張云輝陳海元汪迎節朱曉妹宋春鳳方先文
華北農學報 2020年6期
關鍵詞:水稻
張所兵,張云輝,陳海元,林 靜,汪迎節,朱曉妹,宋春鳳,方先文
(1.江蘇省農業科學院 種質資源與生物技術研究所,江蘇省農業種質資源保護與利用平臺,江蘇 南京 210014;2.江蘇省中國科學院 植物研究所(南京中山植物園),江蘇 南京 210014)
水稻栽培方式主要有移栽和直播2種栽培模式。移栽稻是將水稻種子先育苗,再栽插到大田中的一種栽培方式。直播稻是指將水稻種子直接播入大田中的一種栽培方式。較之移栽稻,直播稻省去了育秧和移栽2個環節,極大節約了種植成本,提高了經濟效益。目前,許多國家水稻栽培模式主要以直播為主,如美國、澳大利亞、意大利、日本、斯里蘭卡和馬來西亞等[1-3]。隨著農村青壯年勞動力向非農業領域轉移,我國直播稻面積也呈現逐年增加的趨勢。直播稻的栽培面積在2007年以前的僅為2%,現在已增加至30%以上[4]。
制約直播稻發展的主要因素之一是直播時低溫、低氧的環境因素導致的出苗不齊、缺苗等現象[1, 5-10]。解決該問題的有效措施是提高水稻種子耐低溫和低氧發芽能力。已有研究證實,水稻低氧發芽力在不同的水稻品種間存在明顯差異,且低氧發芽力是多基因控制的復雜的數量性狀[11-13]。侯名語等[11]利用Kinmaze/DV85 重組自交系群體分析了控制水稻低氧發芽力的QTL, 共檢測到5 個低氧發芽力QTL,分別位于第1,2,5,7 號染色體上,貢獻率10.5%~19.6%。Jiang等[14]利用USSR5/N22 F2群體定位了2個低氧發芽力QTLs,分別定位于第5,11號染色體,分別可以解釋15.51%和10.99%表型變異。陳孫祿等[12]利用R0380/RP2334回交自交系群體定位了4個低氧發芽力QTLs,分別定位于第2,3,8號染色體,貢獻率最小的為9.37%,貢獻率最大為17.34%。孫凱等[15]利用200 份水稻種質為材料對低氧條件下,芽鞘長度進行GWAS分析,共檢測到15 個與芽鞘長顯著關聯的位點,分布在第3,4,5,6,8,11 染色體上。Yang等[16]利用YZX/02428重組自交系群體定位了25個低氧發芽力的QTL,分別位于除第11號染色體外的其他11條染色體上。
雖然人們在水稻低氧發芽資源篩選及相關的QTL定位和基因克隆等方面取得了一定進展,但總的來說,控制低氧發芽遺傳位點的挖掘仍然比較有限。篩選新的耐淹水種質資源,定位更多控制低氧發芽的QTL,并對相關基因進行克隆,對水稻低氧萌發育種和闡明耐低氧發芽分子機理有重要意義。本研究以云南省秈稻地方品種扎西瑪和江蘇省優良食味粳稻品種南粳46 構建的RIL群體,對控制低氧發芽性狀進行QTL定位,為下一步基因克隆和育種利用奠定基礎。
1 材料和方法
1.1 試驗材料
以云南省秈稻地方品種扎西瑪(母本)與江蘇省優良食味粳稻品種南粳46(父本)為親本,用單粒傳法構建了含有135個家系的RIL群體[15-17]。
2019年正季,親本材料及其包含135個家系的RIL群體種植于江蘇省農業科學院南京試驗田。株距和行距分別為13.3,30.0 cm。田間管理與一般大田相同。
1.2 低氧發芽檢測
為了打破休眠,將親本及各家系的種子置于50 ℃烘箱中放置7 d。低氧發芽試驗參照陳孫祿等[12]和Jiang等[14]進行,具體如下:從親本及135個家系中,隨機選取健康飽滿一致的種子,用84消毒液(原液)消毒30 min,然后用無菌去離子水清洗5次后,置于裝滿無菌去離子水的10 cm高的離心管中,并密封。每份材料4管,每管5粒, 3次重復。于人工氣候室中28 ℃,黑暗培養。7 d后調查胚芽鞘長度,作為低氧發芽力的指標[12,16]。
1.3 QTL分析
用QTL IciMapping 4.0[18]檢測低氧發芽力QTL,LOD閾值設為2.5。QTL 命名按McCouch等[19]的方法進行。
SSR標記RM1300和RM1227選自Gramene(https://archive.gramene.org/)。RM1300引物序列為:RM1300-F:CAGCCATGAATGTTGGCTAC和RM1300-R:GCCATGTCCATTTATGGTGC,RM1227引物序列為:RM1227-F:CATGGTAGCACACACCCTTG和RM1227-R:CATCGACATGTGGACCACTC。
2 結果與分析
2.1 親本及群體低氧發芽分析
在低氧條件下發芽7 d后,以胚芽鞘長度為標準,對扎西瑪和南粳46低氧發芽力進行了調查,結果發現:扎西瑪胚芽鞘長度為2.92 cm,而南粳46胚芽鞘長度為6.68 cm,二者差異在P=0.01水平上達到極顯著水平(圖1)。扎西瑪/南粳46 RIL群體胚芽鞘長度呈現為連續的分布,胚芽鞘長度為1.32~5.99 cm,群體胚芽鞘長度次數呈正態的分布(圖2)。
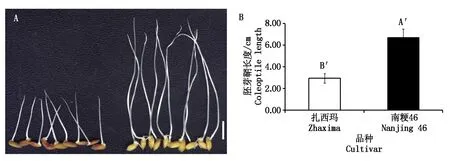
A.低氧條件下發芽7 d,扎西瑪(左)和南粳46表型(右);B.低氧條件下發芽7 d,扎西瑪和南粳46胚芽鞘長度;A′和B′表示P=0.01水平差異顯著。
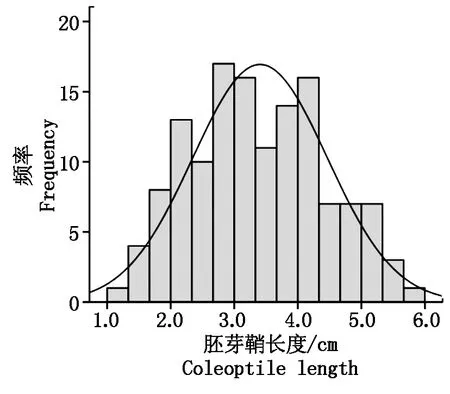
圖2 扎西瑪/南粳46 RIL群體胚芽鞘長度次數分布
2.2 低氧發芽QTL分析
本研究前期構建了扎西瑪/南粳46 RIL群體SSR標記分子連鎖圖譜,該圖譜共包含202對SSR標記,總長度為1 437.3 cM,平均圖距為8.1 cM,能滿足QTL 作圖的要求。根據群體中135個家系低氧發芽力數據,利用QTL Ici-Mapping 4.0對控制低氧發芽QTL進行檢測,在第12號染色體上的RM1300和RM1227之間檢測到一個控制低氧發芽力的QTL(qAG-12),qAG-12位于第12染色體上26.00~27.34 Mb的1.34 Mb范圍內。該QTL來自南粳46,LOD值為3.04,可以解釋11.24%的表型變異(圖3)。
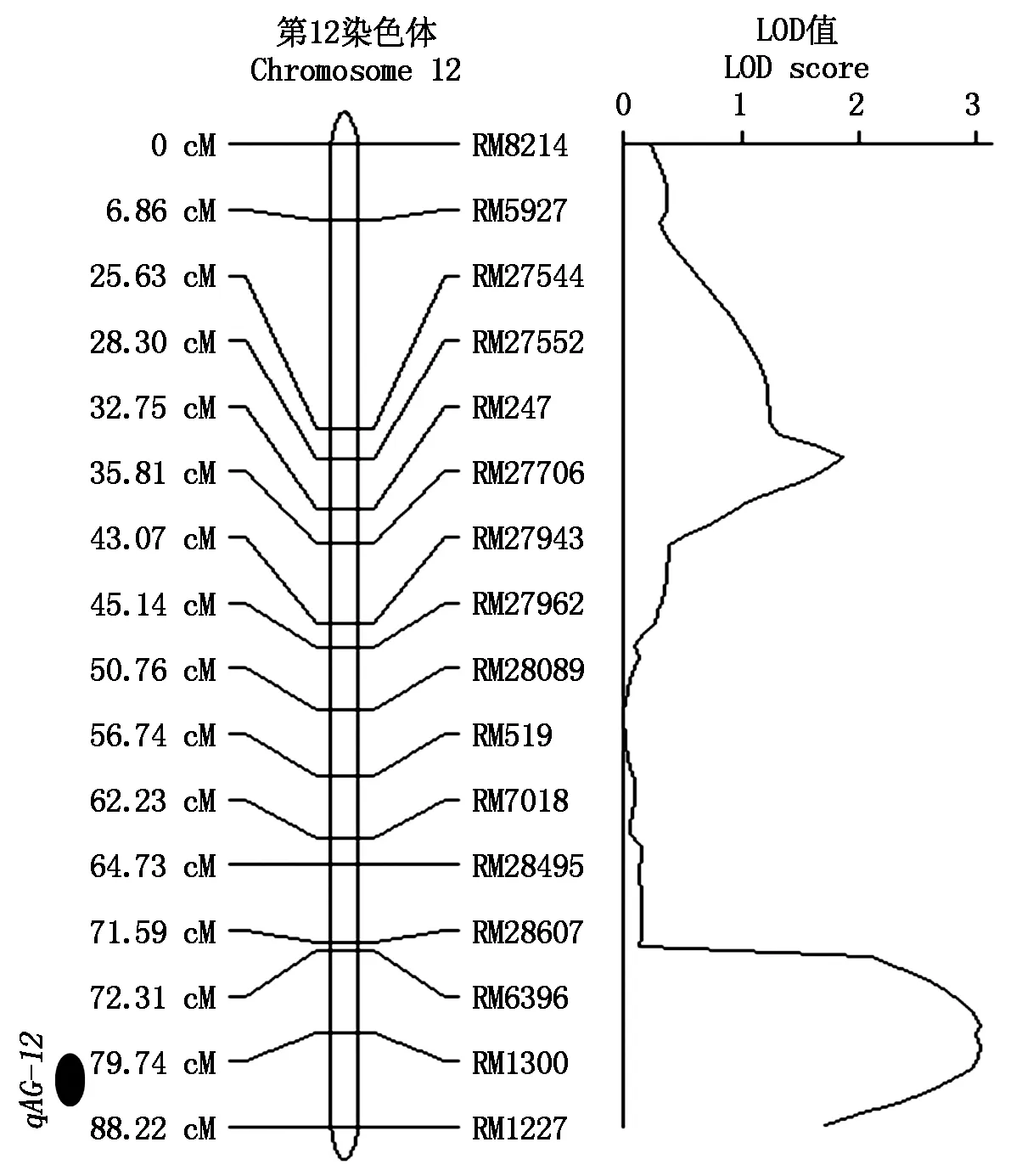
圖3 水稻低氧發芽力QTL檢測
3 討論與結論
耐淹成苗率低、雜草問題等是影響直播稻產量的重要因素。與傳統的移栽稻相比,直播稻種子在萌發時處于淹水環境之中。淹水環境有利于控制田間雜草卻不利于直播稻成苗,水直播稻成苗率通常在30%左右[20]。這主要歸因于以往的育種過程中,很少對低氧萌發等與直播水稻相關的性狀進行人為選擇,這直接導致目前市場上正在推廣的絕大多數水稻品種不適合進行田間直播[20-22]。開展耐淹水的種質資源篩選,挖掘在低氧條件下萌發能力強的水稻資源,并在水稻育種過程中加以利用和選擇是提高耐淹成苗率的關鍵。此外,控制耐淹成苗率的遺傳位點的挖掘仍然比較有限,且控制耐淹成苗率的遺傳位點都是由多基因控制的復雜的數量性狀,遺傳力較低,常規的育種選擇費時費力。因此,開展控制耐淹成苗率的遺傳位點的定位研究,繼而進行多基因分子標記輔助聚合育種,有望提高水稻種子低氧發芽能力,進而改善解決直播稻全苗問題。
本研究對云南地方品種扎西瑪和江蘇省優質稻品種南粳46低氧發芽進行了鑒定,發現二者差異極顯著,進而利用扎西瑪/南粳46 RIL群體在12染色體上定位到一個主效QTL,位于SSR標記RM1300和RM1227之間,為該QTL育種利用提供了材料和分子標記。將RM1300和RM1227在日本晴基因組中進行定位,發現其位于第12染色體上26.00~27.34 Mb。該候選區域未見報道的低氧發芽的QTL,故qAG-12是一個新的QTL。在Rice Genome Annotation Project(http://rice.plantbiology.msu.edu/index.shtml)對該候選區間注釋發現,在這1.34 Mb范圍內共有200多個注釋基因。接下來,擬從扎西瑪/南粳46 RIL群體中選擇含有qAG-12單株與扎西瑪構建F2次級分離群體,對qAG-12進行精細定位,為下一步基因克隆奠定基礎。
在淹水條件下,扎西瑪和南粳46種子低氧發芽力呈現極顯著差異,并利用扎西瑪/南粳46 RIL群體在第12染色體上定位到一個新的控制水稻種子低氧發芽力的QTL(qAG-12)。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
